UNIVERSITA' DEGLI STUDI DI PADOVA
Dipartimento di Scienze Biomediche Sperimentali
SCUOLA DI DOTTORATO DI RICERCA IN BIOSCIENZE
INDIRIZZO BIOLOGIA CELLULARE
CICLO XXI
Il recettore olfattivo e la formazione dei circuiti nel
sistema olfattivo: meccanismi molecolari associati al
recettore sul cono di crescita.
Direttore della Scuola : Ch.mo Prof. Tullio Pozzan
Supervisore : Dr.ssa Claudia Lodovichi
Dottoranda : Micol Maritan
1
2
Indice
SUMMARY
pag. 7
SOMMARIO
pag. 9
1. INTRODUZIONE
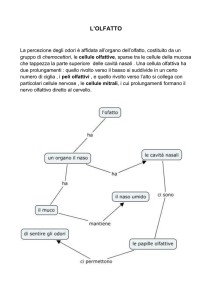
1.1 L’olfatto.
pag. 11
1.2 Il sistema olfattivo principale.
pag. 11
1.3 L’epitelio olfattivo.
pag. 12
1.4 I recettori olfattivi.
pag. 14
1.5 Via di trasduzione del segnale odoroso.
pag. 16
1.6 Componenti molecolari della via di trasduzione del segnale
pag. 17
odoroso.
1.6.1 Le adenilato ciclasi.
pag. 17
1.6.2 Le fosfodiesterasi.
pag. 19
1.6.3 I canali associati a nucleotidi ciclici.
pag. 20
1.7 Il codice combinatoriale.
pag. 21
1.8 Il bulbo olfattivo.
pag. 22
1.9 Organizzazione dell’epitelio olfattivo e del bulbo olfattivo.
pag. 26
1.9.1 Organizzazione dell’epitelio olfattivo.
pag. 27
1.9.2 Organizzazione del bulbo olfattivo.
pag. 27
1.9.3 Formazione dei glomeruli.
pag. 30
1.9.4 Colonna olfattiva.
pag. 31
1.9.5 Ruolo del RO nella convergenza assonale e nella
pag. 32
formazione dei circuiti intra- ed inter-bulbari.
3
1.10 Axon guidance.
pag. 34
1.11 Formazione della mappa sensoriale nel sistema olfattivo.
pag. 38
1.12 Il progetto di ricerca.
pag. 47
1.13 Sensori per il cAMP.
pag. 48
1.14 Espressione di Gs e Golf.
pag. 51
2. MATERIALI E METODI
2.1 Colture primarie di neuroni sensoriali olfattivi.
pag. 53
2.2 Trasfezione di neuroni sensoriali olfattivi.
pag. 54
2.3 Imaging di cAMP in neuroni sensoriali olfattivi isolati.
pag. 54
2.4 Traslocazione nucleare della subunità catalitica di PKA.
pag. 56
2.5 Ca2+ imaging in neuroni sensoriali olfattivi isolati.
pag. 56
2.6 Ca2+ imaging ex vivo.
pag. 57
2.7 Immunocitochimica per OMP.
pag. 58
2.8 Immunoistochimica per Gs.
pag. 58
2.9 Stimoli su neuroni sensoriali olfattivi isolati.
pag. 59
2.10 Stimoli su preparati di emiteste.
pag. 60
2.11 Terreni e soluzioni utilizzate
pag.60
4
3. RISULTATI
3.1 Progetto di ricerca
pag. 63
3.2 Colture primarie di neuroni sensoriali olfattivi
pag. 63
3.3 Trasfezione di neuroni sensoriali olfattivi in coltura
pag. 64
3.4 Dinamiche spazio-temporali di cAMP in neuroni
pag. 65
sensoriali olfattivi isolati.
3.4.1 Effetto dello ione Cl- sull’emissione di fluorescenza
pag. 65
di CFP e YFP.
3.4.2 Dinamiche spazio-temporali di cAMP in neuroni
pag. 67
sensoriali olfattivi trattati con stimoli farmacologici.
3.4.3 Dinamiche spazio-temporali di cAMP in neuroni
pag. 69
sensoriali olfattivi in seguito a stimolazione odorosa.
3.5 Dinamiche spazio-temporali di Ca2+ in neuroni sensoriali
pag. 73
olfattivi isolati.
3.6 Traslocazione nucleare della subunità catalitica di PKA
pag. 76
3.7 Dinamiche spazio-temporali di Ca2+ ex vivo.
pag. 79
3.8 Espressione di Gs
pag. 83
pag. 87
4. DISCUSSIONE
BIBLIOGRAFIA
pag. 93
RINGRAZIAMENTI
pag.103
5
6
Summary
A unique feature in the topographic organization of the olfactory system is the
“dual role” of the odorant receptor (OR). The OR is involved in the transduction
of chemical signals (odors), but it plays also an instructive role in the glomerular
convergence of the olfactory sensory neuron (OSN) axons. The OR is indeed
expressed on the cilia and on the axon termini-growth cone of OSN. The OR is a
G-protein coupled receptor that upon binding of the odorant ligand at the cilia
activates an adenylyl cyclase to synthesize cAMP. The molecular mechanism
underpinning the role of the OR at the axon termini-growth cone remains to be
established. If the OR acts through the generation of cAMP at the growth cone is
not known.
To elucidate the functional properties and intracellular signalling coupled to the
OR on the OSN axon termini, we have investigated the dynamics and the
intracellular distribution of the second messenger cAMP in living OSN of rat in
culture transfected with a genetically encoded, FRET based sensor for cAMP.
Upon treatment with pharmacological agents such as forskolin (a generic activator
of adenylyl cyclases) or IBMX (a non specific inhibitor of phosphodiesterases),
cAMP rises throughout OSN including the axon termini-growth cone. By
analyzing the spatial distribution and the temporal dynamics of cAMP, we found
that the cAMP increase is faster at the neurites than at the soma level.
To assess whether the OR at the axon termini-growth cone is a functional receptor
we challenged the OSN with odor mixture. Odors puff applied to the axon
termini-growth cone produces a rise in cAMP limited exclusively to the growth
cone.
Exposure of OSN to odors stimulates the influx of Ca2+ through cAMP gated
channels (CNG) in the cilia. To assess whether this signalling cascade takes place
at the axon termini-growth cone, we measured Ca2+ signals at the axon terminigrowth cone in OSN loaded with Ca2+ indicator (FURA-2-AM). Odors puff
applied at the growth cone gives rise to a rapid Ca2+ increase limited exclusively
to the axon termini-growth cone.
7
Taken together the present data demonstrate that the OR on the axon terminigrowth cone is a functional receptor, whose activation generates a local rise in
cAMP and Ca2+.
To assess whether the phenomena observed in primary culture of OSN in vitro
take place with analogous dynamics in vivo, we performed Ca2+ imaging
experiments on hemi head explants preparation. In this preparation the mouse
head is cut sagitally and the connections between the olfactory epithelium, the
bulb and the brain are maintained along the medial axis.
OSN were loaded with the Ca2+ indicator Calcium Green dextran and the cells
resulted entirely labeled, from the cilia to the axon termini.
We stimulated labeled glomeruli with pharmacological (forskolin) and
with
physiological stimuli (odor mix). In both cases we found a significant increase in
fluorescence, indicating a rise in Ca2+ concentration in the presynaptic terminals at
the glomeruli level.
The influx of Ca2+ is due to cAMP-induced CNG chanell opening, since it is
abolished in the presence of the adenylyl cyclase blocker SQ22536.
We demonstrated that in isolated neurons as in the intact olfactory bulb the
olfactory receptor at the axon termini-growth cone is functional and coupled to
localized cAMP and Ca2+ increases.
8
Sommario
L’elemento caratterizzante l’organizzazione topografica del sistema olfattivo è il
“duplice ruolo” del recettore olfattivo (RO). Esso è coinvolto nella trasduzione del
segnale odoroso, ma svolge anche un ruolo istruttivo nella convergenza degli
assoni dei neuroni sensoriali olfattivi (NSO) al corretto glomerulo target a livello
del bulbo olfattivo. Questa duplice funzione è corroborata dalla duplice
localizzazione di tale recettore: esso è infatti presente sia sulle cilia dei NSO sia
sull’assone terminale-cono di crescita.
Il RO è un recettore accoppiato a proteine G, il quale, in seguito al legame
dell’odore alle cilia, induce l’attivazione di un’adenilato ciclasi, che porta alla
sintesi di cAMP.
I meccanismi molecolari che sottendono al ruolo del recettore olfattivo sul cono di
crescita rimangono tuttora sconosciuti. Non è noto se il RO in questa posizione sia
attivo e se agisca tramite la sintesi di cAMP.
Per studiare le proprietà funzionali ed il signalling intracellulare associato al
recettore olfattivo sul cono di crescita abbiamo analizzato le dinamiche temporali
e la distribuzione intracellulare del secondo messaggero cAMP in neuroni
sensoriali olfattivi di ratto in coltura, trasfettati con una sonda geneticamente
codificata per il cAMP, basata su FRET.
In seguito al trattamento con stimoli farmacologici (forscolina, un generico
attivatore delle adenilato ciclasi, ed IBMX, un inibitore aspecifico delle
fosfodiesterasi), si registra un incremento di cAMP nell’intero neurone sensoriale
olfattivo, compreso l’assone terminale-cono di crescita. Analizzando la
distribuzione spaziale e le dinamiche temporali del segnale cAMP, abbiamo
osservato che l’incremento di cAMP si verifica più velocemente nei neuriti
rispetto al soma.
Per stabilire se il RO a livello del terminale assonico-cono di crescita è un
recettore funzionale, abbiamo stimolato i NSO con una miscela di odori.
L’applicazione diretta di un puff odoroso all’assone terminale-cono di crescita
induce un aumento di cAMP localizzato esclusivamente al cono di crescita.
9
L’esposizione dei NSO agli odori porta ad un influsso di Ca2+ attraverso i canali
CNG nelle cilia.
Per stabilire se la stessa cascata di signalling si verifichi all’assone terminale-cono
di crescita, abbiamo misurato il segnale Ca2+ a livello del terminale assonico in
NSO caricati con un indicatore del Ca2+ (FURA-2-AM). Un puff di odori
applicato all’assone terminale-cono di crescita induce un rapido aumento di Ca2+
circoscritto esclusivamente al terminale assonico.
I dati ottenuti dimostrano che il recettore olfattivo all’assone terminale-cono di
crescita è un recettore funzionale, la cui attivazione genera incrementi locali di
cAMP e di Ca2+.
Per valutare se i risultati ottenuti in vitro non fossero dovuti alle condizioni di
coltura ed all’isolamento dei neuroni, ma fossero presenti nel bulbo olfattivo
intatto, abbiamo effettuato esperimenti di imaging del Ca2+ sul sistema olfattivo
integro, utilizzando particolari preparati, definiti espianti di emiteste. In questi
preparati la testa di un topo è tagliata sagittalmente e le connessioni tra l’epitelio
olfattivo, il bulbo ed il cervello sono mantenute lungo l’asse mediale. I neuroni
sensoriali olfattivi sono caricati con un indicatore del Ca2+ (Calcium Green
dextran) e le cellule risultano marcate completamente, dalle cilia fino al terminale
assonico.
I glomeruli marcati sono stati stimolati con stimoli farmacologici (forscolina) e
fisiologici (mix di odori). In entrambi i casi si è osservato un significativo
aumento nella fluorescenza, indicante un incremento della concentrazione di Ca2+
nei terminali pre-sinaptici a livello dei glomeruli. L’influsso di Ca2+ è dovuto
all’apertura dei canali CNG indotta dal cAMP, poichè il segnale Ca2+ è abolito in
presenza dell’inibitore delle adenilato ciclasi SQ22536.
Abbiamo quindi dimostrato che sia in neuroni sensoriali olfattivi isolati sia nel
bulbo olfattivo integro il recettore olfattivo al terminale assonico-cono di crescita
è un recettore funzionale ed accoppiato ad aumenti locali di cAMP e Ca2+.
10
1. Introduzione
1.1 L’olfatto.
I mammiferi sono in grado di riconoscere e distinguere migliaia di odori diversi
presenti nell’ambiente anche a bassissime concentrazioni, che influenzano il loro
comportamento e forniscono informazioni essenziali per la loro sopravvivenza. Il
sistema olfattivo è coinvolto infatti in molteplici meccanismi fisiologici, quali
risposte emozionali (ansia, paura, piacere), funzioni riproduttive (comportamenti
sessuali e materni) e relazioni sociali (riconoscimento di individui della stessa
specie o famiglia, delle prede e dei predatori). Per svolgere queste funzioni così
diverse i mammiferi possiedono due organi olfattivi anatomicamente e
funzionalmente distinti. Il sistema olfattivo principale, costituito dall’epitelio
olfattivo, dal bulbo olfattivo principale e dalle aree corticali e non, correlate,
permette il riconoscimento di molecole odorose volatili. Il sistema olfattivo
accessorio, costituito dall’organo vomeronasale, dal bulbo olfattivo accessorio e
dalle aree corticali e non, correlate, è invece specializzato nel rilevamento dei
feromoni, sostanze secrete da un organismo, che regolano risposte fisiologiche e
comportamentali degli individui della stessa specie.
In questa sede ci limiteremo a trattare il sistema olfattivo principale.
1.2 Il sistema olfattivo principale.
La percezione olfattiva ha inizio nei neuroni sensoriali olfattivi (NSO) presenti
nell’epitelio nasale. Questi neuroni trasmettono poi il segnale al bulbo olfattivo
principale e da qui passa alla corteccia cerebrale. Iniziamo quindi a trattare più
dettagliatamente i diversi componenti del sistema olfattivo principale.
11
1.3 L’epitelio olfattivo.
L’ epitelio olfattivo è un epitelio colonnare pseudostratificato, che riveste strutture
cartilaginee convolute dette turbinati, poste nella parte posteriore delle cavità
nasali. L’epitelio olfattivo è costituito da tre tipi cellulari principali: i neuroni
sensoriali olfattivi, le cellule sustentacolari di sostegno e le cellule staminali basali
(1,2). I neuroni sensoriali olfattivi rappresentano il 70-80% della popolazione
cellulare totale dell’epitelio olfattivo e rigenerano costantemente durante la vita
dell’organismo con un’emivita di circa 60-90 giorni. Essi hanno una tipica
morfologia bipolare, con un unico dendrite non arborizzato, che si porta verso la
superficie dell’epitelio, a contatto con le cavità nasali, e l’assone amielinico che
proietta al bulbo olfattivo nel cervello (fig.1). Il dendrite termina con
un’espansione globosa, detta knob, da cui si dipartono numerose cilia, estensioni
filiformi che protrudono nella cavità nasale e su cui si trovano i recettori olfattivi
(RO).
Fig.1: rappresentazione di un neurone sensoriale olfattivo. E’ possibile osservare la tipica
morfologia bipolare: un unico dendrite non arborizzato che termina con un’espansione globosa
(knob), da cui dipartono numerose cilia, ed un unico assone. (3)
Le cilia sono immerse nel muco secreto dalle cellule sustentacolari dell’epitelio
olfattivo e dalle ghiandole di Bowman (fig.2). Si ritiene che le specifiche
caratteristiche biochimiche del muco siano rivolte a creare l’ambiente ideale per la
percezione degli odori. Il muco contiene proteine leganti gli odori (olfactory
binding protein, OBP), che sono secrete dalla ghiandola nasale laterale; esse
hanno la funzione di legare gli odoranti idrofobici, permettendo il loro passaggio
12
alla soluzione acquosa che costituisce il muco. Sono presenti più forme di OBP
nell’epitelio nasale, nel ratto ne sono state identificate quattro (1).
Gli odori si dissolvono nel muco dell’epitelio olfattivo per raggiungere e legarsi ai
recettori olfattivi presenti sulle cilia ed innescare una catena di segnali
intracellulari, che culmina con la generazione del potenziale d’azione e la
trasmissione del segnale al bulbo olfattivo nel cervello.
Le cellule sustentacolari di sostegno svolgono diverse funzioni: isolano
elettricamente i neuroni sensoriali olfattivi, secernono le componenti del muco
che riveste la superficie dell’epitelio olfattivo e fattori di crescita importanti per lo
sviluppo dei neuroni sensoriali olfattivi e contengono enzimi detossificanti che
inattivano gli odori.
Le cellule basali di tipo globoso, che costituiscono la lamina basale al di sotto
dell’epitelio olfattivo, rappresentano i precursori dei neuroni sensoriali olfattivi.
Fig.2: rappresentazione dell’epitelio olfattivo. Si distinguono i tre tipi cellulari: i neuroni
sensoriali olfattivi, le cui cilia sono esposte verso il lume della cavità nasale e sono immerse nel
muco, le cellule di supporto sustentacolari, e le cellule staminali basali. Sulle cilia dei NSO sono
espressi i recettori olfattivi che legano le molecole odorose e originano la cascata di trasduzione
del segnale odoroso. (4)
13
1.4 I recettori olfattivi
Gli odori sono piccole molecole organiche, che si differenziano tra loro per
numerosi parametri, tra cui la struttura stereochimica della catena idrocarbonica, il
tipo e la posizione dei gruppi funzionali laterali, le dimensioni e la carica. La
percezione di un dato odore dipende dalla struttura della molecola odorosa, ma è
influenzata anche da altri parametri, quali la concentrazione della sostanza
odorosa e la variabilità soggettiva. Per esempio la molecola di indolo viene
percepita come un odore putrido ad alte concentrazioni, mentre a basse
concentrazioni come un odore floreale. L’androstenedione, invece, a basse
concentrazioni è percepito come odore di urina da alcuni individui, come odore
mediamente piacevole da altri e non è percepito per niente da altri ancora (5).
Le molecole odorose vengono riconosciute e legate a livello delle cilia dei NSO
dai recettori olfattivi (RO). L’esistenza di tali recettori è stata dimostrata nel 1991
da R.Axel e L.Buck (6) , valendo loro il premio Nobel per la Medicina nel 2004.
Questi recettori appartengono ad una grande famiglia multigenica che nei
mammiferi comprende più di 1000 membri, arrivando ad occupare fino al 2% del
genoma nel topo (6,7). I geni codificanti per i RO sono raggruppati in cluster
presenti nella maggior parte dei cromosomi (7,8). Da un punto di vista evolutivo,
il numero degli pseudogeni nell’ambito della famiglia dei RO è andato
aumentando. Essi sono infatti solo il 20% nei roditori, che contano quindi un
repertoire molto più vasto rispetto ai mammiferi superiori, specie l’uomo, in cui
circa il 60% dei geni codificanti per i RO sono pseudogeni (7-10). L’espressione
di questi geni è ristretta ai neuroni sensoriali olfattivi maturi ed inizia durante la
vita embrionale, prima che l’assone raggiunga il bulbo olfattivo (4) .Per
distinguere i NSO maturi da quelli immaturi viene utilizzata l’olfactory marker
protein (OMP). Questa è una proteina citoplasmatica, di funzione non nota,
espressa tipicamente ed esclusivamente dai neuroni sensoriali olfattivi maturi.
Essa inizia ad essere espressa intorno al giorno embrionale 13-14 (E13-E14) nel
topo e al giorno 18 (E18) nel ratto e viene considerata un marker caratteristico dei
NSO maturi (11,12). Ogni neurone olfattivo esprime un solo tipo di RO, cosicchè
ciascun neurone sensoriale olfattivo è distinto funzionalmente ed identificato
14
molecolarmente dal tipo di recettore espresso, dando luogo alla regola: un
recettore-un neurone (13-15).
I RO sono espressi in modo monoallelico: in ciascun NSO l’espressione del
recettore olfattivo deriva esclusivamente da un allele (15). Questo meccanismo
sembra assicurare che un solo tipo di recettore sia presente in ogni neurone
sensoriale.
Da un punto di vista strutturale, la regione codificante dei RO è lunga circa 1kb ed
è priva di introni. I RO appartengono alla famiglia dei recettori associati a
proteine G e ne condividono le caratteristiche principali, quali la presenza di sette
domini transmembrana legati da loop intracellulari ed extracellulari di varia
lunghezza e numerose sequenze brevi altamente conservate. Tuttavia presentano
anche tratti peculiari dei recettori olfattivi, quali un secondo loop extracellulare
insolitamente lungo con residui di cisteina altamente conservati. Il grado di
omologia nella sequenza dei RO varia tra il 40 ed il 96%. Da notare la presenza di
regioni ipervariabili, in cui la sequenza è fortemente diversa tra un recettore e
l’altro, nella terza, quarta e quinta regione transmembrana. Nei modelli
tridimensionali di recettori associati a proteine G, questi tre tratti formano una
tasca che molto probabilmente costituisce il sito di legame per le molecole
odorose (fig.3) (4). Nei neuroni sensoriali olfattivi sono espressi molteplici tipi di
proteine G. In particolare sono presenti due tipi di proteine G stimolatorie, Gs e
Golf (G olfactory protein), ciascuna capace di attivare l’adenilato ciclasi (16). Nei
NSO maturi, Golf è maggiormente espressa ed associata al RO posto sulle cilia nel
processo di trasduzione del segnale odoroso. Topi knock out per Golf presentano
infatti una drammatica riduzione delle risposte agli odori. Gs, invece, è espressa
soprattutto nel corso dello sviluppo embrionale e nelle fasi precoci di sviluppo
post-natale (17).
15
Fig.3: struttura di un recettore olfattivo. I residui aminoacidici più conservati sono rappresentati
nelle tonalità del blu, mentre quelli caratterizzati da una maggiore variabilità sono rappresentati
nelle tonalità del rosso (4).
1.5 Via di trasduzione del segnale odoroso.
La cascata di trasduzione del segnale attivata dal legame delle molecole odorose
ai recettori olfattivi presenti sulle cilia dei NSO è ben nota e coinvolge come
principali secondi messaggeri il cAMP ed il Ca2+ (18-20).
Una volta entrati nella cavità nasale, gli odori si disciolgono nel muco che ricopre
la superficie dell’epitelio olfattivo e si legano ai recettori olfattivi sulle cilia dei
neuroni sensoriali olfattivi. In seguito a tale legame si ha l’attivazione della
proteina G stimolatoria Golf (21), che a sua volta attiva l’adenilato ciclasi di tipo 3
(AC3) (22), portando ad un incremento della concentrazione di cAMP all’interno
della cellula. Il cAMP si lega alla porzione intracellulare dei canali CNG,
causandone l’apertura (20). Questi canali permettono l’ingresso sia di Ca2+ che di
Na+. L’aumento della concentrazione di Ca2+ nelle cilia in seguito all’apertura dei
canali CNG regola sia l’attivazione sia la desensitizzazione dei neuroni sensoriali
olfattivi. Infatti l’aumentata concentrazione di Ca2+ induce una corrente uscente di
Cl- e quindi un’ulteriore depolarizzazione, che porta all’insorgenza di un
potenziale d’azione (23,24). Il Ca2+ ha anche un effetto di feedback negativo a
vari livelli della catena di trasduzione del segnale. Il Ca2+, legando la calmodulina,
forma un complesso Ca2+-calmodulina (Ca2+-CaM) che si associa direttamente ai
canali CNG, riducendone l’affinità per il cAMP e quindi la probabilità di apertura
16
dei canali stessi (fig.4) (25). Il Ca2+ ha inoltre un’azione inibitoria sull’adenilato
ciclasi, bloccando la sintesi di cAMP, ed un’azione attivatoria sulle fosfodiesterasi
di tipo 1C2, aumentando la degradazione di cAMP.
Fig.4: trasduzione del segnale odoroso nelle cilia dei neuroni sensoriali olfattivi (26). In seguito al
legame con la molecola odorosa, il recettore olfattivo attiva la proteina Golf che a sua volta attiva
l’adenilato ciclasi di tipo 3. L’incremento di cAMP che ne segue porta all’apertura dei canali
CNG, con conseguente influsso di Ca2+ e Na+. L’aumento di Ca2+ permette l’apertura dei canali
per il Cl- ,che porta alla fuoriuscita di Cl- dalla cellula ed ulteriore depolarizzazione.
1.6 Componenti molecolari della via di trasduzione del segnale
odoroso.
Diversi studi hanno permesso di identificare i componenti molecolari della via di
signalling attivata dal recettore olfattivo sulle cilia dei NSO, quali: le adenilato
ciclasi (AC), le fosfodiesterasi (PDE) e i canali associati ai nucleotidi ciclici
(CNG). Analizziamo ora più dettagliatamente i vari elementi della via di
trasduzione del segnale odoroso.
1.6.1 Le adenilato ciclasi.
Le adenilato ciclasi sono delle ATP-pirofosfato liasi che convertono l’ATP in
adenosina monofosfato ciclico (cAMP) e pirofosfato. Al momento sono note 10
17
isoforme differenti di AC, di cui nove associate alla membrana (AC1-9), mentre la
decima è una proteina solubile (27). Tutte le isoforme associate alla membrana
sono attivate dal legame con la proteina Gs, mentre l’adenilato ciclasi solubile è
attivata da HCO3- (28) e da Ca2+ (29). La tabella 1.1 riporta alcune proprietà delle
AC di membrana.
Tabella 1.1: proprietà regolatorie delle adenilato ciclasi associate alla membrana (27). Le AC di
membrana sono regolate da molteplici effettori, tra i quali i principali sono: le proteine G (Gsα,
Gβγ, Gαi, z, o) la proteine chinasi A e C (PKA e PKC rispettivamente), la calmodulina (CaM) e le
chinasi dipendenti da questa (CaMK). Vi sono poi altri regolatori secondari, come: RGS2
(regolatore del signalling delle proteine G); PAM (proteina associata a Myc); Ric8a (fattore
scambiatore di nucleotidi guaninici per le proteine G); Snapin: (membro del complesso
SNAP25/Snare nei neuroni ippocampali); PP2a (proteina fosfatasi 2a).
Le adenilato ciclasi associate alla membrana possono essere classificate in quattro
gruppi principali, definiti in base alle loro proprietà regolatorie. Il gruppo I è
formato dalle AC stimolate dal Ca2+, il gruppo II comprende le AC stimolate dalle
subunità βγ delle proteine G; il gruppo III consiste delle AC inibite da Giα/Ca2+,
mentre l’ultimo gruppo è rappresentato dalla AC9, che è insensibile alla
forscolina, un attivatore generico delle altre AC.
Nei NSO è stata dimostrata la presenza delle adenilato ciclasi 2, 3 e 4 (30,31). Le
isoforme 2 e 4 sono espresse a livello delle cilia; dati recenti mostrano che l’ AC3
è espressa anche a livello dell’assone terminale ed in piccole concentrazioni,
appena rilevabili con saggi di immunoistochimica, nel soma (32). E’ stato
dimostrato che l’adenilato ciclasi 3 è accoppiata ai recettori olfattivi sulle cilia ed
è quindi fondamentale per la trasduzione del segnale olfattivo. Infatti, topi knock
18
out per l’AC3 non mostrano risposte elettrofisiologiche ad un’ampia gamma di
odori. Inoltre si è osservato che in queste linee di animali transgenici la mappa
sensoriale è fortemente alterata (vedi §1.11) questi risultati indicano che l’AC3 è
necessaria per la trasduzione del segnale odoroso a livello delle cilia ed inoltre
svolge un ruolo nella formazione della mappa sensoriale a livello dell’assone
terminale.
1.6.2 Le fosfodiesterasi.
Le fosfodiesterasi sono fosfoidrolasi che catalizzano l’idrolisi dei nucleotidi
ciclici cAMP e cGMP. Controllando la velocità di degradazione di questi
importanti secondi messaggeri, le PDE giocano un ruolo critico nel signalling
intracellulare. Esistono 11 famiglie differenti di PDE, ciascuna comprendente a
sua volta varie isoforme (33-35).
Tabella 1.2: classificazione delle 11 famiglie di fosfodiesterasi (34). Le PDE 4, 7 e 8 sono
specifiche per il cAMP, mentre le PDE 5, 6 e 9 idrolizzano il cGMP. Le PDE 1, 2, 3, 10 e 11
presentano infine una doppia specificità per entrambi i nucleotidi ciclici. Alcune famiglie sono
regolate dal cGMP, altre dal complesso Ca2+-calmodulina, altre ancora da PKA o PKG. IBMX è
un inibitore farmacologico generico delle PDE, il Rolipram è un inibitore specifico per le PDE4.
Le transducine sono proteine G specifiche della retina.
19
Come si può osservare in tabella 1.2, le famiglie di fosfodiesterasi presentano
specificità di substrato (cAMP e cGMP) diverse e differenti proprietà regolatorie.
Si possono distinguere tre gruppi principali, costituiti dalle famiglie di enzimi che
idrolizzano specificamente il cAMP, il cGMP o che presentano una doppia
specificità. Inoltre le PDE si differenziano per le loro proprietà regolatorie; tra i
principali modulatori della loro attività troviamo il cGMP stesso (che può fungere
sia da attivatore che da inibitore), il complesso Ca2+-calmodulina e le proteine
chinasi PKA e PKG.
Nei NSO sono stati identificati tre tipi di fosfodiesterasi:1, 2 e 4. Le PDE1
presentano alti livelli di espressione nelle cilia, mentre le PDE2 sono espresse
nelle cilia, nel soma e negli assoni; le PDE4, infine, localizzano nel knob
dendritico, nel soma e negli assoni (1,36).
Le fosfodiesterasi di tipo 1 sono regolate dal Ca2+ via calmodulina ed idrolizzano
sia il cAMP che il cGMP, anche se la specificità varia a seconda delle isoforme.
Nei NSO la forma predominante è la PDE1C2, che presenta affinità simili per
entrambi i nucleotidi ciclici (37).
Le PDE2 sono anch’essi enzimi a doppia specificità, che risultano stimolati
allostericamente dal legame del cGMP.
Le PDE4, infine, sono specifiche esclusivamente per il cAMP. E’ stato dimostrato
che la fosforilazione da parte di PKA aumenta l’attività delle PDE4 (38).
1.6.3 I canali associati a nucleotidi ciclici.
I canali associati a nucleotidi ciclici (CNG) sono stati identificati inizialmente nei
bastoncelli e nei coni della retina (39).
I CNG presenti nelle cilia dei NSO sono canali cationici permeabili sia al Ca2+ che
al Na+. Essi presentano un elevato grado di omologia con i CNG della retina (più
del 80% di identità di sequenza aminoacidica) e sono strutturalmente omologhi. I
CNG olfattivi sono tetrameri costituiti da tre diverse subunità: 2 A2, 1 A4 e 1
B1b. Le ultime due subunità svolgono un ruolo modulatorio: aumentano la
sensibilità al cAMP e permettono la regolazione via Ca2+-calmodulina (fig.5). La
struttura di ciascuna subunità è simile a quella dei canali cationici dipendenti dal
potenziale: esse presentano sei domini transmembrana, con un loop tra il quinto
20
ed il sesto dominio che forma il poro centrale del canale, e le regioni N- e Cterminali intracellulari (40,41).
Fig.5: rappresentazione dei canali CNG olfattivi e delle tre subunità che li formano. In blu sono
indicati i siti di legame per il cAMP, mentre in giallo ed in nero i siti di legame per la calmodulina
(40).
I CNG del sistema olfattivo presentano un’affinità maggiore per il cGMP rispetto
al cAMP come mostrano i valori di kM (concentrazione richiesta per raggiungere
metà dell’attivazione massima): 3 µM per il cAMP e 1.6 µM per il cGMP (42).
L’affinità per i ligandi è modulata dal legame Ca2+-CaM, da fosforilazione e da
cationi bivalenti (43,44).
1.7 Il codice combinatoriale
Abbiamo precedentemente detto che ogni NSO esprime un solo tipo di RO. Il
recettore espresso definisce il “molecular receptive range” del neurone, cioè
determina il range di odori che quel singolo NSO è in grado di legare e
riconoscere (5). Ogni recettore olfattivo riconosce una specifica caratteristica
strutturale della molecola odorosa (odotopo). Le due principali classi di odotopi
che differenziano i diversi odori sono: la struttura stereochimica della catena
idrocarbonica, il tipo e la posizione dei gruppi funzionali laterali. Poiché ogni
molecola odorosa può presentare al suo interno più odotopi, ciascun odore potrà
essere riconosciuto da diversi recettori olfattivi che riconoscono i diversi epitopi
di cui è costituito. Ogni singolo RO è però in grado di riconoscere molecole
odorose diverse che presentano lo stesso odotopo. In questo modo ciascun odore
sarà codificato dall’attivazione di una particolare combinazione di recettori
olfattivi, ma ogni singolo recettore può essere una delle componenti del codice di
21
diversi odori. Questo viene definito codice combinatoriale e permette al sistema
olfattivo di riconoscere un numero enorme di odori (fig.6).
Fig.6: modello di codice combinatoriale. A sinistra sono rappresentate varie molecole odorose con
i vari odotopi raffigurati in colori diversi. A destra sono rappresentati nei corrispondenti colori i
RO attivati da tali epitopi. L’identità dei diversi odori è codificata perciò da diverse combinazioni
di recettori. Tuttavia ciascun recettore può fungere da componente del codice combinatoriale per
odori diversi. Dato il numero immenso di possibili combinazioni di RO, questo schema permette
di discriminare un numero enorme di diversi odori (5).
1.8 Il bulbo olfattivo
Il bulbo olfattivo costituisce la prima stazione di ritrasmissione dell’informazione
olfattiva. Come altre aree cerebrali presenta tre tipi di neuroni: i neuroni afferenti,
i neuroni efferenti e gli interneuroni.
I neuroni sensoriali olfattivi costituiscono le afferenze sensoriali del bulbo
olfattivo. L’assone dei NSO è un processo sottile (0.2 µm di diametro),
amielinico, non ramificato, che dopo aver attraversato l’osso cribriforme
raggiunge il bulbo olfattivo (fig.7).
22
Fig.7: rappresentazione di una sezione sagittale di naso di topo (4). La freccia indica il punto di
ingresso dell’aria e delle molecole odorose. L’epitelio olfattivo (OE) riveste le strutture dei
turbinati ed è suddiviso in quattro zone, rappresentate nei diversi colori. Gli assoni dei neuroni
sensoriali olfattivi proiettano al bulbo olfattivo (MOB, main olfactory bulb), dove formano i
glomeruli.
Il bulbo olfattivo fa parte del prosencefalo ed è costituito da due strutture ovoidali
simmetriche che si trovano immediatamente sopra le cavità nasali. Esso ha una
caratteristica struttura laminare, suddivisa in: strato del nervo olfattivo (ONL),
strato glomerulare (GL), strato plessiforme esterno (EPL), strato delle cellule
mitrali (MCL), strato plessiforme interno (IPL) e strato delle cellule dei granuli
(GCL) (fig.8). Ognuno di tali strati contiene tipi cellulari diversi.
Fig.8 : immagine di sezione di bulbo olfattivo in seguito a colorazione di Nissl. E’ possibile
osservare i diversi strati che caratterizzano il bulbo olfattivo: lo strato dei glomeruli (GL); lo strato
plessiforme esterno (EPL), che contiene i corpi cellulari delle cellule tufted; lo strato delle cellule
mitrali (MCL); lo strato plessiforme interno (IPL) e lo strato delle cellule dei granuli (GCL).
23
Lo strato del nervo olfattivo è formato dagli assoni dei neuroni sensoriali olfattivi,
che attraversano la lamina basale dell’epitelio olfattivo e l’osso cribriforme per
raggiungere infine il bulbo olfattivo.
Nello strato glomerulare gli assoni dei NSO stabiliscono sinapsi eccitatorie,
glutamatergiche, con i dendriti delle cellule post-sinaptiche del bulbo olfattivo
(cellule mitrali, tufted e periglomerulari), dando origine ai glomeruli. I glomeruli
sono strutture sferiche di neuropilo, di dimensioni variabili tra gli 80 ed i 160 µm
di diametro (nel topo da 30 a 50 µm, nel coniglio e nel gatto da 100 a 200 µm),
disposte su tutta la superficie esterna dei bulbi. Sono costituiti dalla ramificazione
terminale degli assoni dei NSO, che contraggono sinapsi con i dendriti delle
cellule post-sinaptiche del bulbo, nonché con le cellule periglomerulari, poste
tutto intorno ai singoli glomeruli. I glomeruli sono inoltre circondati dalle cellule
gliali (45,46).
I neuroni post-sinaptici del bulbo sono rappresentati dalle cellule mitrali e dalle
cellule tufted. Queste cellule costituiscono la via efferente del bulbo olfattivo e
formano il tratto olfattivo laterale. Le cellule mitrali sono neuroni glutamatergici
che presentano i corpi cellulari disposti a formare un unico strato, lo strato delle
cellule mitrali. I corpi cellulari delle cellule tufted si trovano invece a diversi
livelli nello strato plessiforme esterno ed in base a tale posizione vengono distinte
in esterne, medie ed interne. Sia le cellule mitrali che le cellule tufted presentano
un unico dendrite primario, che penetra in un unico glomerulo nel quale forma
un’estesa arborizzazione. Presentano inoltre dendriti secondari che decorrono
orizzontalmente nell’EPL e contraggono sinapsi dendro-dendritiche con le cellule
dei granuli. Gli assoni delle cellule mitrali e delle cellule tufted si portano fuori
dal bulbo olfattivo, formando il tratto laterale olfattorio, che trasmette
l’informazione olfattiva ad altre stazioni cerebrali. Le cellule tufted esterne
rientrano negli interneuroni. Anche da un punto di vista biochimico, le cellule
tufted si presentano come una popolazione eterogenea, che si avvale di diversi
neurotrasmettitori quali la colecitochinina (CCK), il GABA, il fattore rilasciante
la corticotropina (CRF), il polipeptide intestinale vasoattivo (VIP) (45,46).
Al di sotto dello strato delle cellule mitrali si trova lo strato plessiforme interno,
molto sottile, costituito da pochi corpi cellulari, prevalentemente appartenenti a
cellule ad assone corto e cellule dei granuli. Gli interneuroni del bulbo sono
24
rappresentati dalle cellule periglomerulari e dalle cellule dei granuli. Le cellule
periglomerulari (PG) includono diversi tipi di cellule, quali le cellule
periglomerulari, le cellule ad assone corto e le cellule tufted esterne. Le cellule PG
contraggono sinapsi con i dendriti delle cellule mitrali e delle cellule tufted, oltre
che con i terminali assonali dei NSO. Le cellule PG sono una popolazione
eterogenea non solo morfologicamente, ma anche biochimicamente. Esse infatti
esprimono diversi neurotrasmettitori quali: il GABA, l’ossido nitrico, il NADPH,
le dopamine ed il neuropeptide Y. Pertanto, le sinapsi che le cellule PG formano
con le cellule pre- e post-sinaptiche del bulbo non hanno solamente funzione
inibitoria, ma in senso più ampio modulatoria.
Le cellule dei granuli sono neuroni gabaergici, il cui corpo cellulare si trova nello
strato dei granuli, che è immediatamente al di sotto dello strato plessiforme
interno, ed occupa la parte centrale del bulbo olfattivo. Esse formano sinapsi
inibitorie dendro-dendritiche con i dendriti secondari delle cellule mitrali e delle
cellule tufted. Mancano di assone (fig.9).
Le cellule PG e le cellule dei granuli rigenerano costantemente durante tutta la
vita dell’individuo. Esse originano dalla zona sub ventricolare (SVZ) intorno ai
ventricoli laterali e da qui si portano attraverso uno specifico processo di
migrazione lungo la rostral migratory stream (RMS). Una volta giunte al bulbo
olfattivo, migrano radicalmente e si differenziano in interneuroni bulbari (45,46).
25
Fig.9: diagramma dei circuiti base che riassume l’organizzazione sinaptica del bulbo olfattivo.
Due moduli (rosa e blu) rappresentano due diversi tipi di RO. Le cellule mitrali (M) e tufted (T)
sono i neuroni efferenti del bulbo, le cellule dei granuli (Gr) e le cellule periglomerulari (PG)
sono interneuroni locali (47).
1.9 Organizzazione dell’epitelio olfattivo e del bulbo olfattivo.
La specificità delle connessioni tra i neuroni pre- e post-sinaptici è essenziale per
il normale funzionamento del sistema nervoso centrale. Lo sviluppo ed il
mantenimento dei circuiti neuronali è regolato da molecole espresse in tempi e
sedi specifici nel corso dello sviluppo e da meccanismi dipendenti dall’attività
neuronale.
Nei sistemi sensoriali, i neuroni sensoriali presenti nelle strutture anatomiche più
periferiche sono ordinati in maniera tale che i neuroni che decodificano
caratteristiche simili dello stimolo sono disposti secondo un preciso ordine
spaziale. I neuroni sensoriali a loro volta indirizzano il loro assone in specifici
target del sistema nervoso centrale, mantenendo i rapporti spaziali presenti nelle
strutture periferiche, dando luogo a mappe topografiche. Le mappe topografiche
permettono di tradurre il mondo esterno in una rappresentazione interna, in
termini di rappresentazione neuronale.
26
Il sistema olfattivo si differenzia per varie ragioni da altri sistemi sensoriali.
Innanzitutto è privo di un’organizzazione spaziale periferica. Inoltre i neuroni
sensoriali rigenerano costantemente nel corso della vita dell’individuo, per cui i
circuiti neuronali si riformano continuamente.
1.9.1 Organizzazione dell’epitelio olfattivo.
Nell’epitelio olfattivo i neuroni sensoriali non sono disposti secondo un preciso
ordine spaziale, ma sono distribuiti in modo pressocchè casuale. Analisi della
distribuzione spaziale dei neuroni esprimenti recettori olfattivi diversi, tramite
ibridazione in situ, hanno rivelato che i NSO che esprimono un dato recettore
sono ristretti ad una delle quattro zone (in direzione dorso-ventrale) in cui viene
suddiviso l’epitelio olfattivo. Tuttavia, all’interno di ciascuna zona, i neuroni
esprimenti recettori diversi sono frammisti fra loro in modo casuale (14,48).
1.9.2 Organizzazione del bulbo olfattivo.
La situazione è completamente diversa a livello del bulbo olfattivo, dove è
presente una mappa topografica ben precisa ed altamente conservata. Le
proiezioni assonali di neuroni esprimenti lo stesso recettore convergono con
squisita precisione a formare glomeruli in specifiche posizioni sul lato mediale e
sul lato laterale di ciascun bulbo olfattivo.
Questo è stato dimostrato con metodiche di ibridazione in situ da Ressler (49) e da
Vassar (50). La disposizione dei glomeruli riceventi fibre esprimenti lo stesso RO
risulta simmetrica nei due bulbi ed è costante in animali diversi della stessa
specie. Fu avanzata quindi l’ipotesi che gli odori possano essere codificati da un
pattern specifico di glomeruli attivati. Questo dato sarà poi confermato da
esperimenti di imaging in vivo condotti in studi successivi (51,52).
Nel 1996 Mombaerts e coll. furono in grado di visualizzare i singoli neuroni
sensoriali olfattivi, e quindi la convergenza dei singoli assoni, grazie
all’elaborazione di una nuova strategia genetica (53).
Questa metodica molecolare prevede la modifica del locus del gene codificante
per i recettori olfattivi, in modo che tale locus codifichi per un RNA messaggero
bicistronico, per cui il RO è co-espresso con uno specifico marker (tau-LacZ). La
sostituizione del gene endogeno con il costrutto si ottiene mediante
27
ricombinazione omologa, per cui alla fine si ottengono linee di topi geneticamente
modificate, in cui uno specifico recettore è co-espresso con il marker. Un esempio
è dato dal recettore P2. Nei topi geneticamente modificati (P2-IRES-tau-LacZ) il
recettore P2 è co-espresso con tau-LacZ; pertanto, in seguito a colorazione con Xgal, i neuroni esprimenti P2 risultano marcati nella loro interezza, dalle cilia sino
alle ramificazioni dell’arborizzazione terminale assonica (fig.10). In questo modo
tali neuroni possono essere facilmente identificati nell’epitelio e nel bulbo. Inoltre
è possibile seguire il destino di ogni singola fibra esprimente un dato recettore
olfattivo e quindi visualizzare la mappa sensoriale del bulbo olfattivo. Questi
esperimenti hanno inoltre confermato quanto osservato precedentemente da
Ressler e Vassar tramite le ibridazioni in situ. I neuroni esprimenti il recettore P2
proiettano a due glomeruli posti uno sul lato mediale ed uno sul lato laterale di
ciascun bulbo olfattivo. I glomeruli P2 presentano una struttura omogenea,
risultando composti esclusivamente da fibre esprimenti solo il recettore P2.
Fig.10: immagine dei turbinati e del lato mediale del bulbo di un topo ingegnerizzato P2-IREStau-LacZ, dopo colorazione con X-gal. Tutti i neuroni sensoriali olfattivi esprimenti il recettore
olfattivo P2 esprimono anche LacZ ed in seguito a colorazione con X-gal risultano così marcati. Si
osserva come tutti questi neuroni convergano in un unico glomerulo mediale nel bulbo (53).
In studi successivi il marcatore LacZ è stato spesso sostituito con GFP, che offre il
vantaggio di essere immediatamente visualizzabile anche in animali in vivo
(54,55). Questa nuova tecnica ha permesso di analizzare nel dettaglio la
formazione dei glomeruli relativi a NSO che esprimono un dato recettore.
Nel bulbo olfattivo si possono distinguere due livelli di organizzazione
topografica: gli assoni di tutti i neuroni sensoriali olfattivi esprimenti lo stesso RO
28
convergono a formare lo stesso glomerulo ed i glomeruli occupano posizioni
specifiche simmetriche tra i due bulbi ed identiche tra animali diversi della stessa
specie.
Le quattro zone in cui è stato suddiviso l’epitelio olfattivo corrispondono
all’incirca a quattro zone nel bulbo olfattivo, in senso dorso-ventrale (47,50).
Pertanto gli assoni dei neuroni sensoriali localizzati in una data zona dell’epitelio
proiettano alla zona corrispondente nel bulbo olfattivo (fig.11). Per esempio i
neuroni che si trovano nella zona più dorsale dell’epitelio formano glomeruli nella
zona più dorsale del bulbo, mentre i neuroni localizzati più ventralmente
proiettano a formare glomeruli nella zona più ventrale del bulbo.
Fig.11: schema che illustra il pattern di convergenza assonale tra l’epitelio olfattivo ed il bulbo.
Nel topo l’epitelio olfattivo è diviso in quattro zone, definite dal pattern di espressione di
determinati RO. I NSO di una data zona proiettano a glomeruli localizzati nella zona
corrispondente del bulbo olfattivo. Assoni di NSO esprimenti lo stesso recettore (blu e rosso)
convergono in pochi glomeruli definiti (47).
Nonostante i NSO rigenerino costantemente nel corso della vita di un individuo,
la specificità della convergenza assonale, e quindi la mappa glomerulare, è
perfettamente conservata (56).
In base alla modalità di convergenza degli assoni dei NSO, il concetto di codice
combinatoriale può essere esteso anche al bulbo olfattivo. Infatti, poiché ciascun
odore viene codificato dall’attivazione di una particolare combinazione di
recettori olfattivi, a livello del bulbo questo si riflette nell’attivazione dei
corrispondenti glomeruli. Pertanto un odore è rappresentato da una specifica
combinazione spaziale di glomeruli attivati (mappa funzionale), che presenta una
simmetria bilaterale nei due bulbi dello stesso animale e rimane invariata in
29
animali diversi. Ciascun glomerulo può invece far parte di mappe diverse,
corrispondenti ad odori differenti (51).
1.9.3 Formazione dei glomeruli.
Innanzitutto è opportuno sottolineare che i glomeruli non rappresentano una
popolazione omogenea che si sviluppa in modo sincrono ed è influenzata nel
corso della sua formazione e del suo mantenimento dalle stesse molecole e nella
stessa misura dall’attività neuronale evocata o spontanea. Questi elementi
controllano la formazione dei glomeruli in modo diverso a seconda del recettore
espresso dai neuroni convergenti a formare quel dato glomerulo (57).
Si possono tuttavia rintracciare tappe comuni che portano alla formazione dei
glomeruli, sebbene siano appunto percorse in tempi diversi dai glomeruli
esprimenti recettori olfattivi differenti e influenzate da molteplici fattori.
Gli assoni dei NSO, una volta passato l’osso cribriforme, raggiungono il bulbo
olfattivo (intorno a E13), attraversano lo strato del nervo olfattivo e giungono
infine allo strato glomerulare (58,59). In un primo tempo i neuroni che esprimono
lo stesso recettore proiettano ad un’area del bulbo olfattivo. Progressivamente gli
assoni si riuniscono in un’area sempre più stretta e definita, in modo da formare
glomeruli veri e propri. Inizialmente per ogni lato, mediale e laterale, di ciascun
bulbo olfattivo si formano molteplici glomeruli connessi tra loro da fasci di
assoni. Con il procedere dello sviluppo le connessioni tra glomeruli omogenei
(formati da fibre esprimenti lo stesso recettore) vengono eliminate, come pure si
riduce il numero dei glomeruli soprannumerari, per arrivare al termine dello
sviluppo ad uno o due glomeruli per ciascun lato di ogni bulbo (60). In una prima
fase di sviluppo i glomeruli non sono omogenei, vale a dire innervati da neuroni
esprimenti solo lo stesso tipo di recettore, ma possono presentare innervazione da
parte di fibre esprimenti recettori diversi (eterogenei). Questi glomeruli misti
scompaiono con il progredire dello sviluppo per riarrangiamento ed eliminazione
delle “scorrette” proiezioni assonali (61).
Riportiamo due esempi relativi allo sviluppo di glomeruli formati da NSO
esprimenti recettori diversi.
Gli assoni dei neuroni sensoriali esprimenti il recettore P2 proiettano allo strato
del nervo olfattivo al giorno embrionale 14.5 (E14.5) e a E15.5 questi assoni
30
terminano nello strato glomerulare presuntivo. Nei cinque giorni successivi si
organizzano in strutture discrete simili ai glomeruli olfattivi. In molti casi si
osservano due glomeruli appaiati collegati inizialmente da fasci di assoni P2.
Questa connessione viene persa entro il giorno post-natale (PD) 7.5. Durante il
primo periodo post-natale si osservano assoni aberranti di NSO P2 entrare in
glomeruli non appropriati o continuare la crescita oltre lo strato glomerulare.
Questi neuroni non sono più visibili nell’animale adulto (58).
Gli assoni esprimenti M72 iniziano a formare proto-glomeruli in una fase molto
più tardiva, nei primi giorni di sviluppo post-natale, PD2-PD3. Fino a PD10
molteplici glomeruli sono presenti su ogni lato di ciascun bulbo. Tra PD10 e
PD40 si assiste ad un processo di progressiva eliminazione dei glomeruli
sopranumerari, cosicchè nella maggior parte dei casi da PD40 in poi solo singoli
glomeruli sono osservabili sul lato mediale e laterale di ciascun bulbo. E’
importante sottolineare che i glomeruli M72 inizialmente possono essere innervati
anche da fibre M71. La sequenza di questi due recettori, infatti, presenta un alto
grado di omologia (96%). Tuttavia, con il corso dello sviluppo i glomeruli misti
vengono eliminati (60). Per valutare l’effetto dell’attività neuronale evocata sulla
formazione dei glomeruli, lo sviluppo di glomeruli M71 e M72 è stato osservato
in animali in cui è stata attuata occlusione nasale al momento della nascita. In
questi animali, a differenza dei controlli, persiste la presenza di glomeruli multipli
su ogni lato del bulbo olfattivo. Ciascun bulbo presenta dunque un numero
maggiore di glomeruli rispetto ai controlli. Analizzando la struttura dei glomeruli
sopranumerari, si è visto che questi sono eterogenei. Sembra quindi che l’assenza
di attività evocata impedisca il processo di rifinitura delle proiezioni assonali ed in
particolare l’eliminazione di glomeruli eterogenei. La convergenza assonale non
sembra invece significativamente influenzata dall’assenza di attività evocata
(60,62).
1.9.4 Colonna olfattiva.
In ciascun glomerulo convergono gli assoni di NSO esprimenti un unico tipo di
recettore, che formano sinapsi con uno specifico gruppo di cellule mitrali e tufted.
Ogni glomerulo viene a definire un’unità funzionale detta “colonna olfattiva”, in
analogia alle colonne presenti nella corteccia visiva primaria. Ciascuna colonna
31
olfattiva è costituita da un gruppo specifico di cellule mitrali, tufted,
periglomerulari, che ricevono input da uno specifico gruppo di neuroni sensoriali,
e dalle cellule dei granuli che sono ad esse connesse. Le colonne olfattive
associate allo stesso recettore olfattivo sono dette isofunzionali o omologhe (45).
1.9.5 Ruolo del RO nella convergenza assonale e nella formazione dei
circuiti intra- ed inter-bulbari.
Come precedentemente detto, gli assoni dei neuroni sensoriali olfattivi esprimenti
lo stesso tipo di recettore proiettano, per ciascun bulbo olfattivo, a due glomeruli
distinti, uno in posizione mediale ed uno in posizione laterale. Il bulbo olfattivo
presenta pertanto due mappe speculari e simmetriche di glomeruli isofunzionali.
E’ stato dimostrato che queste due mappe di glomeruli omologhi presenti in ogni
bulbo non sono indipendenti, ma reciprocamente connesse in modo estremamente
preciso da un link inibitorio dovuto alle cellule tufted esterne (63). Gli assoni delle
cellule tufted esterne connesse ad un dato glomerulo formano sinapsi eccitatorie
con i dendriti delle cellule dei granuli nella parte opposta del bulbo. Queste
ultime, a loro volta, formano sinapsi inibitorie con le cellule tufted esterne del
glomerulo omologo (fig 12).
Queste connessioni sono reciproche. In tal modo le colonne odorose isofunzionali
vengono integrate in un’unica mappa funzionale.
32
Fig.12: rappresentazione schematica del circuito inibitorio che connette le colonne odorose
isofunzionali nelle due metà di ciascun bulbo. Gli assoni delle cellule tufted esterne (ETC),
connessi con un dato glomerulo, formano sinapsi eccitatorie (+) con i dendriti delle cellule dei
granuli (GC) in un’area ristretta dello strato plessiforme interno sul lato opposto del bulbo
olfattivo. Queste cellule dei granuli, a loro volta, formano sinapsi inibitorie (-) con le cellule tufted
esterne connesse al glomerulo omologo. Queste connessioni sono reciproche. OSN: neuroni
sensoriali olfattivi. GL: strato glomerulare. EPL: strato plessiforme esterno. GCL: strato delle
cellule dei granuli.
Non solo i due glomeruli omologhi appartenenti allo stesso bulbo sono
interconnessi, ma anche i glomeruli isofunzionali presenti nei due bulbi sono
collegati tra loro (64). E’ stato mostrato che le cellule mitrali e tufted associate ad
uno specifico glomerulo proiettano topograficamente ad una regione esterna dei
nuclei olfattivi anteriori (NOApE), dove i neuroni a loro volta proiettano alle
cellule dei granuli alla base dei glomeruli isofunzionali omologhi sull’altro bulbo.
Il bulbo olfattivo appare quindi come una struttura dotata di almeno tre livelli di
organizzazione topografica: i) NSO esprimenti lo stesso recettore olfattivo
convergono con i loro assoni a formare glomeruli simmetrici all’interno di ciascun
bulbo; ii) i due glomeruli omologhi appartenenti allo stesso BO sono collegati da
un circuito intrabulbare inibitorio; iii) i glomeruli isofunzionali presenti nei due
bulbi olfattivi sono collegati da un circuito interbulbare. Da quanto detto emerge
che l’elemento attorno a cui ruota l’organizzazione topografica del BO è il
recettore olfattivo.
33
Rimangono ancora da chiarire i meccanismi che sottendono alla formazione di tali
circuiti.
1.10 Axon guidance.
Il sistema nervoso è formato da circuiti neuronali molto complessi, per la
formazione dei quali è essenziale che i diversi neuroni proiettino i loro assoni
verso i corretti target. Il cono di crescita è una struttura localizzata all’estremità
terminale degli assoni, dotata di grande motilità, che permette di rilevare e
rispondere ai segnali dell’ambiente circostante che guidano gli assoni verso il
target appropriato. Il cono di crescita è costituito da lamellipodia, formati da un
network di filamenti di actina, e da filipodia, strutture tensili composte da fasci di
F-actina, che sondano l’ambiente extracellulare. Questo network di filamenti di
actina è associato a sua volta a microtubuli nella parte distale dell’assone.
L’avanzamento e la ritrazione del cono di crescita dipendono dalla
polimerizzazione e depolimerizzazione dell’actina nei filipodia e nei lamellipodia
e dalla velocità di flusso retrogrado della F-actina all’interno di queste strutture
(65). Le proteine Rho della famiglia delle GTPasi sono degli importanti regolatori
di tali processi di modificazione del citoscheletro (fig.13).
Il cono di crescita è in grado di rilevare molecole extracellulari, dette “segnali
guida”, che dirigono la crescita degli assoni. Tali segnali guida possono avere un
effetto attrattivo o repellente e ciascun tipo può agire a corto o lungo raggio (65). I
segnali guida a corto raggio sono molecole non diffusibili, che si trovano sulla
superficie delle cellule o nella matrice extracellulare, ed agiscono con meccanismi
mediati da contatto cellula-cellula. I segnali guida a lungo raggio sono invece
rappresentati da molecole secrete. Inoltre alcune molecole possono avere sia un
effetto repulsivo che attrattivo. Questa bifunzionalità dipende da diversi fattori, tra
cui lo stato intracellulare del cono di crescita, l’espressione differenziale di
complessi di recettori e l’interazione tra diverse cascate di signalling
intracellulare.
34
Fig.13: rappresentazione grafica del cono di crescita. I lamellipodia contengono un denso reticolo
di actina, i filipodia si estendono e si ritraggono attraverso la regolazione dei processi di
polimerizzazione e depolimerizzazione dei filamenti di F-actina (65).
Sono stati individuati diversi tipi di molecole guida a corto o lungo raggio.
Le molecole di adesione cellulare (CAM), i recettori tirosina chinasi (RPTK), le
efrine, i recettori tirosina fosfatasi (RPTP), le molecole della matrice
extracellulare (ECM) ed i loro recettori, rappresentano tutti segnali guida a corto
raggio.
Le netrine e le neurotrofine, invece, agiscono a lungo raggio. Le semaforine sono
una classe a parte, poiché comprendono sia molecole secrete che di superficie.
Le molecole di adesione cellulare si dividono in tre classi: la superfamiglia delle
immunoglobuline (Ig), delle caderine e delle integrine. Diversi membri delle
prime due famiglie possono mediare adesioni omofiliche, agendo quindi sia come
ligando che come recettore. Alcune proteine CAM, invece, formano legami
eterofilici, fungendo da ligandi o recettori per altre molecole di adesione o per
molecole della matrice extracellulare. Le Ig CAM meglio caratterizzate nel
sistema nervoso sono le CAM neurali (NCAM). Le caderine sono recettori di
adesione dipendenti dal Ca2+ e nel sistema nervoso si distinguono le N-caderine.
Alcune di queste molecole sono collegate all’actina citoscheletrica tramite la
proteina catenina. Infine le integrine sono recettori eterodimerici, che fungono da
legame tra le molecole della matrice extracellulare ed il citoscheletro.
35
I recettori tirosina chinasi comprendono la famiglia dei recettori Eph per le efrine
e dei recettori Trk per le neurotrofine.
I recettori Eph si dividono in due classi: i recettori EphA che legano le efrine-A,
ancorate alla membrana plasmatica cellulare attraverso il glicosilfosfatidil
inositolo (GPI), ed i recettori EphB che sono specifici per le efrine-B
transmembrana. Le interazioni tra le efrine ed i loro recettori richiedono quindi un
contatto
cellula-cellula.
I
complessi
Eph/efrina
trasducono
il
segnale
bidirezionalmente, sia nella cellula esprimente il recettore sia nella cellula
esprimente il ligando. Questo tipo di signalling gioca un ruolo critico durante lo
sviluppo del sistema nervoso.
I recettori Trk riconoscono e legano le diverse neurotrofine, quali NGF, BDNF,
NT-3 e NT-4. Questi fattori, oltre al loro ruolo chiave nella sopravvivenza e nel
differenziamento neuronale, svolgono anche una funzione di molecole guida per
la crescita degli assoni. L’effetto repulsivo o attrattivo delle neurotrofine è
influenzato dal livello di nucleotidi ciclici nel cono di crescita (66-68). Ad
esempio è stato dimostrato che il NGF ha un effetto attrattivo a basse
concentrazioni di cAMP, mentre il BDNF nelle stesse condizioni ha un effetto
repulsivo. Inoltre, le neurotrofine sono anche in grado di modulare la risposta del
cono di crescita assonale ad altre molecole guida, come le semaforine.
I recettori tirosina fosfatasi ed i loro ligandi sono i meno conosciuti. Si pensa che
alcuni tipi di RPTP leghino molecole CAM.
Le molecole della matrice extracellulare sono numerose: laminina, tenascina,
collagene, fibronectina, vibronectina e diversi proteoglicani. I recettori per le
ECM sono in genere integrine, membri della famiglia di recettori Ig e altri
proteoglicani.
Le netrine appartengono ad una piccola famiglia di proteine secrete ed hanno sia
una funzione attrattiva che repulsiva sulla crescita assonale. Questo duplice effetto
dipende sia dai recettori attivati sia da diverse cascate di signalling intracellulare
che regolano in modo differente i livelli citosolici di cAMP. E’ stato dimostrato
infatti che il recettore DCC/UNC-40 media la risposta attrattiva degli assoni alle
netrine, mentre il recettore UNC-5 determina l’effetto repulsivo (65). Inoltre
l’azione della Netrina-1 dipende dalla presenza di altri segnali che modulano i
livelli citosolici sia di cAMP che di Ca2+ (67,69). Il cAMP può dunque agire come
36
interruttore molecolare, andando ad attivare diversi target intracellulari e
determinando così diverse risposte del cono di crescita alle molecole guida.
La famiglia delle semaforine comprende sia proteine secrete che proteine
associate alla membrana cellulare e queste possono avere sia un effetto repulsivo
che attrattivo nell’axon guidance (65,70). Sono stati identificati almeno cinque
classi di semaforine. Tutti i membri di tale famiglia possiedono un dominio Sema
conservato nella regione extracellulare N-terminale. I recettori per le semaforine
sono in realtà dei complessi formati da più molecole. Le plexine sono proteine
transmembrana che si associano alla Neuropilina-1 o Neuroplina-2 (Npn-1 e Npn2), un altro tipo di proteine transmembrana che fungono da co-recettori, dal
momento che le loro corte code citoplasmatiche non hanno alcuna funzione
catalitica. Il complesso plexina-neuropilina può in tal modo legare le semaforine
della classe 3 (Sema-3). La risposta specifica dei neuroni ai diversi membri della
classe Sema-3 dipende dal pattern di espressione delle neuropiline, dal tipo di
neuropilina espressa (Npn-1 o Npn-2) ed anche dal pattern neuronale di
espressione delle plexine durante lo sviluppo del sistema nervoso. Inoltre, come
già detto precedentemente, la risposta dei neuroni alle semaforine è modulata
anche dalle neurotrofine. E’ stato provato che alte concentrazioni di NGF nel
terreno di crescita dei neuroni dei gangli della radice dorsale riducono l’effetto
inibitorio sulla crescita assonale della Sema-3A (71).
Come già accennato nel caso delle netrine e delle neurotrofine, varie prove
sperimentali hanno dimostrato l’importanza del cAMP e del Ca2+ nell’axon
guidance (67,72). Ad esempio, alcune molecole guida determinano nel cono di
crescita segnali Ca2+ che regolano l’attività di altre proteine, coinvolte
nell’organizzazione del citoscheletro e nei movimenti dei filamenti di actina. Il
Ca2+ può mediare sia una risposta di tipo attrattivo che repulsivo. Si pensa che la
polarità della risposta dipenda più dalla grandezza del gradiente Ca2+ che dalla sua
direzione. Ad esempio, un piccolo gradiente di Ca2+ induce repulsione nel cono di
crescita neuronale, mentre un gradiente maggiore induce un effetto attrattivo. E’
stato dimostrato che le vie di segnalazione del cAMP e del Ca2+ interagiscono
nella regolazione dell’axon guidance. L’inibizione della PKA determina un
cambiamento dell’azione attrattiva indotta dal Ca2+ in repulsione e viceversa.
37
Tuttavia i meccanismi alla base di queste interazioni cAMP-Ca2+ sono ancora
sconosciuti (67,72).
1.11 Formazione della mappa sensoriale nel sistema olfattivo.
Come è possibile passare da una mancanza di organizzazione spaziale dei diversi
neuroni olfattivi a livello dell’epitelio olfattivo ad una mappa topografica
estremamente precisa nel bulbo olfattivo? Non solo gli assoni dei neuroni
sensoriali olfattivi che esprimono il medesimo recettore convergono a formare gli
stessi glomeruli, ma anche la localizzazione di questi glomeruli è conservata in
individui della stessa specie.
E’ stato dimostrato che il recettore olfattivo gioca un ruolo istruttivo nella
convergenza dei neuroni sensoriali verso il corretto glomerulo target. Questo dato
è emerso da una serie di esperimenti genetici che hanno dimostrato come
modificazioni della sequenza codificante per i recettori olfattivi porti ad
un’alterazione della mappa sensoriale.
Nei neuroni sensoriali olfattivi esprimenti il recettore P2, l’eliminazione del gene
codificante tale recettore (knock down) impedisce la convergenza degli assoni di
tali neuroni a livello del bulbo olfattivo. In questa circostanza gli assoni si
dirigono in maniera casuale a vari glomeruli (73). Studi successivi hanno
dimostrato che l’eliminazione genetica di un dato recettore olfattivo porta
all’attivazione casuale di un gene codificante per un altro RO (74). Poiché la
scelta del recettore espresso è casuale per ciascun neurone, gli assoni convergono
in differenti glomeruli. Se invece si sostituisce il gene codificante per il recettore
olfattivo normalmente espresso con un diverso RO (swap genico), gli assoni di
tali neuroni convergono a formare glomeruli in una posizione diversa rispetto a
quella occupata dai glomeruli target dei neuroni esprimenti il RO sostituito e di
quelli esprimenti il RO sostitutivo. L’entità della distanza dipende da vari fattori:
dal grado di omologia di sequenza tra i geni codificanti i recettori considerati, dal
cluster di appartenenza dei due geni, dalla zona epiteliale in cui i recettori sono
espressi, dal livello di espressione del recettore. Anche nelle condizioni di
38
massima analogia dei vari parametri analizzati, e.g. stesso cluster sullo stesso
cromosoma, stessa zona epiteliale ed alto grado di omologia di sequenza, persiste
una certa distanza tra il glomerulo nuovo (risultato dello swap genetico) e quelli
originali. Per esempio, i recettori olfattivi P2 e P3 presentano il 75% di identità di
sequenza aminoacidica, i geni che li codificano risiedono sullo stesso cromosoma
e vengono espressi da NSO presenti nella stessa zona dell’epitelio olfattivo. In
esperimenti di swap genetico tra questi due RO (P3P2) i nuovi glomeruli sono
adiacenti ai glomeruli wild type P3 ed anteriori rispetto ai glomeruli wild type P2
(73). Questi dati indicano che il recettore olfattivo gioca un ruolo istruttivo nella
convergenza assonale, ma non è l’unico determinante. I dati ottenuti da questi
esprimenti di swap genetico suggeriscono inoltre che il recettore olfattivo
determini la posizione del glomerulo lungo l’asse antero-posteriore del bulbo
olfattivo, mentre la zona dell’epitelio in cui si trovano i neuroni esprimenti quel
dato RO determini la posizione del glomerulo lungo l’asse dorso-ventrale del BO
(53,73,75).
Studi successivi hanno messo in evidenza il coinvolgimento di varie molecole
guida nella formazione della mappa sensoriale nel bulbo olfattivo, tra cui le efrine,
le semaforine e le neuropiline (68,76,77). E’ stato dimostrato, ad esempio, che
neuroni esprimenti recettori olfattivi diversi esprimono diversi livelli di efrina-A3
ed efrina-A5 sui loro assoni, mentre le cellule post-sinaptiche corrispondenti
esprimono i recettori EphA (68). Alterazioni dei livelli di espressione di queste
molecole alterano la mappa glomerulare, in particolare modificano la posizione
dei glomeruli in senso antero-posteriore nel BO. Topi geneticamente modificati,
che non esprimono né efrina-A3 né efrina-A5, sono stati incrociati con topi P2IRES-tau-LacZ e con topi SR1-IRES-tau-LacZ, per poter visualizzare i NSO
esprimenti il recettore P2 o SR1 e valutare come la convergenza assonale di tali
neuroni a livello del bulbo sia influenzata dall’alterato livello di espressione delle
efrine. In topi mutanti, privi sia di efrina-A3 che di efrina-A5, neuroni che
esprimono il recettore olfattivo P2 o il recettore olfattivo SR1 mostrano una
posteriorizzazione nella formazione dei corrispondenti glomeruli. Invece,
l’aumento del livello di efrina-A5 risulta in un’ anteriorizzazione della posizione
dei glomeruli dei neuroni analizzati (P2-IRES-efrinaA5-IRES-tau-lacZ). I dati
ottenuti sembrano indicare che le efrine cooperano con il RO nella formazione
39
della mappa glomerulare ed in particolare regolano la posizione dei glomeruli
lungo l’asse antero-posteriore del bulbo olfattivo.
Serizawa e coll. hanno osservato che nei NSO esprimenti recettori diversi
vengono espresse molteplici molecole, non solo le efrine-A5, ma anche le EphA5
e le molecole di adesione omofilica Kirrel-2 e Kirrel-3 (76). L’espressione di tali
molecole si verifica in modo complementare: neuroni esprimenti un dato recettore
presentano alti livelli di Kirrel-2 e bassi livelli di Kirrel-3, mentre per neuroni
esprimenti recettori diversi il pattern di espressione è invertito, bassi livelli di
Kirrel-2 ed alti livelli di Kirrel-3. Lo stesso fenomeno avviene per efrina-A5 e
EphA5. Questa complementarietà è presente a livello dei neuroni sensoriali
dell’epitelio e nei corrispettivi glomeruli a livello del bulbo olfattivo. E’stato
dimostrato inoltre che la trascrizione di questi geni è attività- dipendente. Nei topi
knock out per la subunità A2 dei canali CNG, in cui l’influsso di Ca2+ attraverso
tali canali è abolito, l’espressione di Kirrel-2 ed EphA5 risulta ridotta, mentre
l’espressione di Kirrel-3 ed efrina-A5 è aumentata. L’espressione attivitàdipendente di questi geni è stata dimostrata anche in seguito ad occlusione nasale:
nei NSO esprimenti MOR28 l’espressione di efrina-A5 e Kirrel-3 risulta
incrementata in seguito ad occlusione nasale, mentre l’espressione di EphA5 e
Kirrel-2 è ridotta. In questo studio è stato dimostrato che i geni Kirrel-2, Kirrel-3,
efrina-A5, EphA5, i cui livelli di espressione sono correlati con il RO espresso,
sono trascritti in modo complementare ed attività-dipendente (76).
Tra le altre molecole coinvolte nella formazione di una corretta mappa
glomerulare ricordiamo anche la Semaforina-3A, che viene espressa nel bulbo
olfattivo. Un sottogruppo di NSO esprimono la Neuropilina-1, che rappresenta il
ligando della Semaforina-3A e la loro interazione è di tipo repulsivo. Gli assoni
dei neuroni sensoriali che esprimono la Neuropilina-1 formano glomeruli nella
regione mediale e laterale del BO, evitando selettivamente la regione di
espressione della Semaforina-3A. In topi knock out per questa molecola, gli
assoni di NSO esprimenti Neuropilina-1 proiettano anche nelle regioni del bulbo
che negli animali wild type presentano un’espressione della Semaforina-3A. (77).
L’insieme di questi esperimenti suggerisce che i neuroni esprimenti recettori
diversi co-esprimono molecole segnale diverse, che cooperano con il recettore
nella formazione della mappa glomerulare. Ancora oggi, tuttavia, rimangono da
40
chiarire i meccanismi molecolari che collegano l’identità di un dato RO con i
diversi tipi e/o livelli di espressione di molecole guida.
A rinforzare il ruolo del RO nel processo di convergenza assonale esperimenti di
immunoistochimica condotti da Barnea e coll. (78) e da Strotmann e coll. (57)
hanno dimostrato la presenza del recettore olfattivo sulla porzione più distale
dell’assone e sul cono di crescita. Il recettore olfattivo risulta quindi espresso non
solo sulle cilia ma anche sull’assone terminale-cono di crescita. Tuttavia i
meccanismi molecolari che sottendono al ruolo del recettore olfattivo sul cono di
crescita rimangono ancora da chiarire.
Ci si è chiesti quindi in quale modo elementi molecolari della via di signalling
intracellulare accoppiata al RO influenzino la trasduzione del segnale odoroso e la
formazione della mappa sensoriale. Per rispondere a questa domanda sono state
create una serie di linee di topi transgenici, in cui elementi del signalling
intracellulare associato al RO sono stati geneticamente modificati. I canali CNG
sono una componente centrale dell’apparato di trasduzione del segnale odoroso.
Topi knock out per tali canali sono anosmici, cioè non esibiscono risposte
elettrofisiologiche evocate dagli odori in risposta ad un ampio range di stimoli
diversi. L’incrocio di queste linee con topi P2-IRES-tau-LacZ e la successiva
analisi dei bulbi di questi animali hanno permesso di esaminare la posizione
relativa di glomeruli P2 e M50 (questi ultimi visualizzati con metodiche di
ibridazione in situ). Il relativo spaziamento e l’ordine dei glomeruli sono
mantenuti anche in topi knock out per CNG. Sembra perciò che l’attività evocata
non sia necessaria per la formazione di una corretta mappa glomerulare (79).
Questo dato è sostenuto anche dai risultati ottenuti in topi geneticamente
modificati mancanti di Golf (17). Questa proteina è associata al RO e permette la
trasduzione del segnale odoroso attivando l’adenilato ciclasi 3. Benchè Golf sia
molto più abbondante di Gs nell’epitelio olfattivo di topi adulti, entrambe le
molecole potrebbero contribuire alla trasduzione del segnale evocato dall’odore.
Topi knock out per Golf sono anosmici, mostrano una forte riduzione delle risposte
elettrofisiologiche associate alla somministrazione di un gran numero di odori.
Incrociando queste linee con topi P2-IRES-tau-LacZ è stato possibile osservare se
la convergenza dei NSO esprimenti P2 fosse alterata. Gli assoni di tali neuroni
sensoriali olfattivi convergono correttamente a formare un unico glomerulo P2
41
mediale e laterale. La posizione di entrambi i glomeruli è costante in tutti gli
animali analizzati. Questi studi sembrano quindi avvalorare l’ipotesi che l’attività
odore-evocata non sia necessaria per la formazione di una corretta mappa
topografica. Tali esperimenti sono stati condotti in condizioni in cui l’attività
odore-dipendente è inibita in tutta la popolazione di neuroni sensoriali olfattivi e
perciò non escludono un ruolo dell’attività evocata in un ambiente competitivo.
Zhao e Reed (80) hanno sfruttato la localizzazione del gene della subunità
OCNC1 dei canali CNG sul cromosoma X per creare un modello di studio del
ruolo dell’attività nello sviluppo del sistema olfattivo in un contesto competitivo.
Hanno generato due linee di topi in cui un allele OCNC1 è stato sostituito con il
reporter tau-LacZ o tau-EGFP, rendendo possibile la marcatura dei neuroni che
non esprimono quel gene. Nelle femmine eterozigoti, sfruttando il fenomeno
dell’inattivazione casuale del cromosoma X, si crea una situazione unica in cui
due diverse popolazioni cellulari coesistono. In queste linee di animali
geneticamente modificati, si assiste inizialmente ad una normale formazione dei
glomeruli e successivamente, nel corso della vita dell’animale, la popolazione di
neuroni OCNC1- viene eliminata dall’epitelio olfattivo. Questa eliminazione dei
neuroni “inattivi” dipende dall’esposizione agli odori. Infatti, in seguito ad
occlusione nasale, si osserva una drastica riduzione della perdita dei neuroni
sensoriali olfattivi OCNC1- sia a livello dell’epitelio olfattivo che a livello del
bulbo. Questi risultati indicano che la deplezione dei NSO OCNC1- dipende
dall’attività evocata dall’odore. Viene perciò suggerito che l’attività evocata
contribuisca alla sopravvivenza dei neuroni sensoriali olfattivi in un ambiente
competitivo. Nei bulbi olfattivi di questi animali, tuttavia, si osserva un pattern
inusuale di innervazione dei glomeruli da parte degli assoni OCNC1- e OCNC1+.
Nella maggior parte dei glomeruli la competizione è molto efficiente e nei
glomeruli vengono rinvenute solo le fibre “attive”. Tuttavia alcuni glomeruli
presentano una forte innervazione sia da parte delle cellule inattive sia da parte di
cellule attive. Inoltre la posizione di questi glomeruli rimane invariata tra
individui della stessa specie. Questi risultati indicano che l’attività evocata
conferisce un grosso vantaggio alle fibre attive nel processo di convergenza e di
mantenimento delle proiezioni assonali nella maggior parte, ma non in tutti, i
glomeruli. Questa ipotesi è avvalorata anche dai risultati ottenuti da Zheng e coll.
42
(81). Tramite mutagenesi specifica del gene OCNC1 hanno generato un’altra linea
di topi mancante dei canali CNG e pertanto priva di risposte evocate dagli odori.
Queste linee sono state incrociate con topi in cui neuroni che esprimono un dato
recettore esprimono anche LacZ o GFP. I neuroni marcati da LacZ o GFP sono
facilmente individuabili nel bulbo ed è possibile seguire la convergenza di questi
assoni. Gli assoni di NSO esprimenti M72 non convergono correttamente nel
bulbo olfattivo: già dopo la prima settimana post-natale si osserva la formazione
di glomeruli sopranumerari, disposti più ventralmente rispetto agli originari e
variabili per numero e grandezza. Gli assoni dei neuroni sensoriali esprimenti il
recettore P2, invece, convergono normalmente. Inoltre in questo studio hanno
utilizzato un triplo mutante (M72-IRES-tau-GFP-IRES-ONCN1 eterozigote,
M72-IRES-tau-LacZ
eterozigote,
OCNC1-
eterozigote)
che,
sfruttando
l’espressione monoallelica del recettore olfattivo e l’inattivazione casuale del
cromosoma X, presenta tre popolazioni di NSO che esprimono il recettore
olfattivo M72: GFP/OCNC1+, LacZ/OCNC1+, LacZ/OCNC1-. E’ stato osservato
che NSO esprimenti M72 proiettano i loro assoni in glomeruli separati, a seconda
che siano OCNC1+ o OCNC1-. Questi risultati suggeriscono che la formazione
della mappa glomerulare è influenzata in parte da meccanismi che si basano
sull’attività neuronale evocata. Come si può vedere dall’insieme dei dati raccolti,
finora la situazione è molto complessa ed il ruolo dell’attività evocata nella
formazione dei glomeruli continua ad essere oggetto di dibattito. Un’altra
molecola base nella cascata di trasduzione del segnale odoroso è l’adenilato
ciclasi 3 (AC3). Tramite saggi di immunoistochimica è stata osservata la sua
presenza non solo a livello delle cilia, ma anche lungo gli assoni dei NSO (32).
Topi knock out per questa proteina sono anosmici e presentano una mappa
glomerulare fortemente alterata. Le proiezioni assonali in questi animali sono
fortemente disorganizzate e spesso i glomeruli risultano innervati da assoni
esprimenti recettori diversi. Nella porzione dorso-caudale del bulbo si assiste alla
completa mancanza di glomeruli. Inoltre la formazione dei glomeruli è alterata in
modo variabile a seconda dei diversi recettori analizzati. Nel caso di P2, non si
osservano i glomeruli corrispondenti negli animali mutati. Il fenomeno si associa
ad una drastica riduzione dei neuroni esprimenti P2 nell’epitelio. Per quanto
concerne i NSO esprimenti M72, ai normali glomeruli si associano glomeruli
43
sopranumerari non presenti negli animali wild type di controllo. Si osserva inoltre
uno spostamento della normale posizione dei glomeruli lungo l’asse anteroposteriore del bulbo. Infine, la deplezione della ciclasi comporta la mancata
espressione di Neuropilina-1, una molecola segnale nei processi di allungamento
assonale (32,82). Per quanto concerne il ruolo dell’attività spontanea nella
formazione dei glomeruli, si è visto che essa svolge un ruolo chiave. In animali
sovraesprimenti i canali Kir 2.1 nei neuroni sensoriali, che risultano così
iperpolarizzati e privi di attività spontanea (ed evocata), la mappa glomerulare è
fortemente alterata. Si ha un forte ritardo dell’innervazione del bulbo ed i
glomeruli, anche quando si formano, sono abnormi per dimensioni e struttura.
Ancora una volta neuroni esprimenti recettori diversi sono affetti in maniera
diversa (83) ma tutti risultano comunque influenzare negativamente la formazione
della mappa glomerulare.
Come detto in precedenza, manipolazioni genetiche che aboliscono la sintesi di
cAMP hanno effetti drammatici sull’organizzazione topografica del bulbo
olfattivo, mentre manipolazioni che mantengono la sintesi di cAMP, quali
alterazioni dei canali CNG e di Golf, permettono la formazione di una normale
mappa glomerulare. Questi risultati hanno portato ad ipotizzare che il segnale
cAMP svolga un ruolo chiave nella formazione della mappa sensoriale.
Gli esperimenti di Imai e coll. (84) hanno dimostrato che il segnale cAMP
collegato al RO svolge un ruolo critico nella formazione dei glomeruli. E’ stato
creato il mutante I7-RDY, in cui il RO I7 è mutato nel sito di legame con le
proteina G (la tripletta DRY è modificata in RDY). A seguito di questa mutazione
il recettore non è in grado di associarsi con le proteine G, con conseguente
abolizione della sintesi di cAMP. Nei topi I7(RDY), che esprimono il recettore
mutato, gli assoni sensoriali non solo non convergono nella formazione di alcun
glomerulo, ma non riescono nemmeno a penetrare nello strato glomerulare del
bulbo. Il fenotipo viene completamente recuperato dall’espressione della proteina
Gs costitutivamente attiva nei neuroni I7-RDY(I7(RDY)-IRES-caGs). Il fenotipo
difettivo viene inoltre recuperato anche tramite l’espressione di PKA
costitutivamente attiva. Se però il costrutto I7(RDY) viene co-espresso con CREB
costitutivamente attivo (cAMP response element-binding protein), che è un fattore
di trascrizione attivato da PKA, gli assoni formano strutture glomerulari, ma non
44
si osserva una convergenza completa. E’ stato inoltre analizzato l’effetto
dell’espressione di Gs costitutivamente attiva in NSO esprimenti il recettore I7
wild type. In questo caso i neuroni presentano un eccesso di produzione di cAMP:
i glomeruli si formano in posizione posteriore rispetto ai wild type. L’espressione
di una PKA dominante negativa, invece, anteriorizza la posizione dei glomeruli
corrispondenti. Questi risultati dimostrano che il segnale cAMP collegato al RO è
necessario per la convergenza glomerulare ed inoltre l’aumento o la diminuzione
dei livelli di cAMP modifica la posizione dei glomeruli target dei NSO lungo
l’asse antero-posteriore del bulbo olfattivo. Infine, tramite analisi di microarray e
di RT-PCR sono stati determinati i geni i cui livelli di espressione sono correlati al
segnale cAMP. Uno dei geni così identificati codifica per la Neuropilina-1. Tale
proteina risulta essere espressa dai NSO che esprimono Gs costitutivamente attiva
e dunque presentano alti livelli di cAMP, mentre non è espressa nei neuroni
I7(RDY), dove il segnale cAMP è bloccato. Si è osservato precedentemente (32)
che l’espressione di questa molecola è abolita nei mutanti di AC3. Questi dati
suggeriscono che lo stesso segnale cAMP mediato da Gs regoli la trascrizione di
geni codificanti per proteine coinvolte nell’axon guidance, che a loro volta
partecipano al processo di formazione della mappa glomerulare. Questi risultati
sono convalidati anche dai dati ottenuti da Chesler e coll. (85). Questi autori
hanno utilizzato un approccio di gain-of-function basato su iniezioni di vettori
retrovirali nell’epitelio olfattivo embrionale. Hanno dimostrato che l’espressione
di un recettore funzionale è necessaria e sufficiente a indurre la formazione di
glomeruli. L’espressione ectopica del recettore OR17 (OR17-IRES-EGFP) si è
rivelata infatti sufficiente per la convergenza glomerulare, mentre l’espressione
dello stesso recettore mutato (MycOR17-IRES-EGFP) non funzionante non
permette la coalescenza degli assoni, che entrano in glomeruli diversi. Inoltre
hanno osservato che l’espressione ectopica di Golf costitutivamente attiva è
sufficiente perché gli assoni dei NSO che la esprimono convergano. I risultati
ottenuti hanno portato ad ipotizzare un modello in cui l’attivazione da parte del
RO di una proteina G stimolatoria induce la convergenza degli assoni. Questa
ipotesi è convalidata dai dati ottenuti da Feinstein e coll. (86). Sostituendo il
recettore olfattivo M71 con il recettore adrenergico β2 (β2AR-IRES-tau-LacZ),
che si accoppia a proteine Gs, hanno osservato la formazione di glomeruli
45
funzionali sia mediali che laterali. Se invece si sostituisce il RO con il recettore
vomeronasale V1rb2, che non si accoppia a proteine Gs, non si osserva la
formazione di glomeruli. Poiché topi knock out per la proteina Golf non mostrano
alterazioni della mappa glomerulare, Chesler e coll. (85) hanno ipotizzato che la
proteina G coinvolta nella formazione della corretta mappa sensoriale olfattiva sia
Gs. A sostegno di questa ipotesi c’è il diverso pattern di espressione delle due
proteine G. Gs viene espressa a livello embrionale dalle cellule progenitrici, dai
neuroni olfattivi immaturi e da quelli maturi. A PD5, quando molti glomeruli si
sono già formati, Gs è ancora fortemente espressa dalle cellule progenitrici, ma
solo debolmente da alcuni neuroni immaturi e non è rilevabile nei neuroni maturi.
Golf invece presenta un diverso pattern temporale di espressione: sembra infatti
essere espressa solo dai neuroni maturi, perciò a E15 viene espresso dai pochi
neuroni maturi presenti e dunque la sua espressione totale a livello dell’epitelio è
molto bassa, mentre a PD5 la sua espressione è molto più consistente in quanto
gran parte dei neuroni sono maturi. Gs sembra pertanto essere espressa durante lo
sviluppo embrionale e dai neuroni che stanno rigenerando, mentre Golf viene
espressa soltanto dai neuroni maturi (esprimenti OMP) (16,17). L’insieme di
questi risultati suggerisce che Golf sia associata al recettore olfattivo nella
trasduzione del segnale odoroso, mentre Gs sembra essere associata al recettore
nella trasduzione del segnale alla base dell’allungamento e nella convergenza
assonale.
L’insieme di tutti i dati ottenuti finora indica che il recettore olfattivo svolge una
duplice funzione nei NSO: media la cascata di trasduzione del segnale odoroso e
svolge un ruolo istruttivo nella convergenza assonale insieme a specifiche
molecole guida. Questa ipotesi sembra corroborata dall’espressione del recettore
olfattivo sull’assone terminale-cono di crescita dei NSO. Tuttavia i meccanismi
molecolari che sottendono al ruolo del RO espresso sul cono rimangono oscuri.
Non è noto se il recettore olfattivo in questa posizione sia attivo e quale sia la via
di signalling ad esso associato, in particolare se agisca attraverso cAMP e Ca2+.
Del resto tutti gli studi condotti finora si sono avvalsi di tecniche molecolari,
mancano quindi informazioni funzionali sul recettore e sulla via di signalling ad
esso associata.
46
Rispondere a tali domande risulta essenziale per poter procedere oltre
nell’indagine sul ruolo del recettore nella formazione della mappa sensoriale.
1.12 Il progetto di ricerca.
L’elemento caratterizzante l’organizzazione topografica del sistema olfattivo è il
doppio ruolo del RO. Esso è coinvolto nella trasduzione del segnale odoroso ma
svolge anche un ruolo istruttivo nella convergenza degli assoni dei neuroni
sensoriali nel corretto glomerulo target a livello del bulbo olfattivo, insieme ad
altre molecole guida (49,50). Questa duplice funzione è corroborata dalla duplice
localizzazione di tale recettore: esso è infatti presente sia sulle cilia dei NSO sia
sull’assone terminale-cono di crescita (57,78). Mentre la cascata di trasduzione
del segnale attivata dal RO a livello delle cilia è nota e si basa sul cAMP e sul
Ca2+ come secondi messaggeri (18-20), i meccanismi molecolari che sottendono
al ruolo del recettore olfattivo sul cono di crescita rimangono tuttora sconosciuti.
Non è noto se il RO in questa posizione sia attivo e quale sia la via di signalling
ad esso associata, in particolare se agisca tramite cAMP e Ca2+.
In questo progetto di dottorato ci siamo proposti di analizzare le proprietà
funzionali ed il signalling intracellulare associato al recettore olfattivo sull’assone
terminale-cono di crescita dei neuroni sensoriali olfattivi.
Rispondere a questa domanda è condizione necessaria per investigare
ulteriormente il modo con cui il recettore sul cono di crescita può guidare l’assone
al suo target glomerulare.
Per studiare le proprietà funzionali ed il signalling intracellulare associato al
recettore olfattivo sul cono di crescita abbiamo utilizzato sonde geneticamente
codificate per il cAMP, basate su FRET ed indicatori del Ca2+, quali il FURA-2AM ed il Calcium Green 1 dextran.
47
1.13 Sensori per il cAMP.
Per valutare le dinamiche spazio-temporali del segnale cAMP nei neuroni
sensoriali olfattivi in coltura ci siamo avvalsi di sonde geneticamente codificate,
che si basano sul fenomeno di FRET.
Tali sensori presentano un’alta risoluzione spaziale (<1 µm) e temporale (<
100ms), risultando quindi particolarmente adatte a valutare cambiamenti di cAMP
in vari compartimenti cellulari e di conseguenza a capire i meccanismi di
signalling intracellulare (87).
Il meccanismo di FRET (Fluorescence Resonance Energy Transfer) sfrutta la
presenza di due molecole fluorescenti, dette donatore ed accettore. I due fluorofori
vengono scelti in modo tale che lo spettro di emissione del donatore si
sovrapponga allo spettro di eccitazione dell’accettore (fig.14). Di conseguenza
l’eccitazione del fluoroforo donatore porta ad un’emissione di energia, che a sua
volta induce l’eccitazione dell’accettore. Tale processo avviene in modo ottimale
solo se le due molecole sono a distanza ristretta (<100Å) (88).
Fig.14: spettri di assorbimento e di emissione dei fluorofori YFP e CFP. In grigio è evidenziata
l’area di sovrapposizione tra l’emissione di CFP e l’assorbimento di YFP.
L’efficienza di questo processo (E) dipende dalla distanza che intercorre tra i due
fluorofori. L'efficienza (E) di FRET è infatti definita dall’equazione di Förster:
La distanza tra i fluorofori è indicata con r. R0 rappresenta invece la distanza a cui
metà dell’energia è trasferita. In base all’equazione di Förster, una distanza doppia
48
tra accettore e donatore, per esempio da R0 a 2R0, comporta una diminuzione
dell’efficienza del trasferimento di energia da E = 50% a E = 1,5%. Perciò FRET
è una tecnica che misura in modo molto sensibile le distanze intramolecolari ed i
cambiamenti conformazionali.
Nel nostro progetto abbiamo utilizzato due tipi di sonde: la prima si basa sulla
proteina chinasi A (PKA) e la seconda su Epac. Entrambe le proteine sono target
del cAMP. La PKA è un tetramero formato da due subunità catalitiche (C) e da
due subunità regolatorie (R). La sonda è stata geneticamente modificata fondendo
la subunità catalitica con YFP e la subunità regolatoria con CFP (87,89,90).
In condizioni di basse concentrazioni di cAMP, la PKA si trova nel suo stato
olotetramerico e FRET è massima. Infatti stimolando il fluoroforo CFP alla sua
propria lunghezza d’onda (430 nm), parte della sua energia di emissione (480 nm)
viene trasferita a YFP, che emette alla sua corrispondente lunghezza d’onda (545
nm).
Quando la concentrazione di cAMP all’interno della cellula aumenta, il cAMP si
lega alle subunità regolatorie di PKA, inducendone un cambiamento
conformazionale, a seguito del quale le subunità catalitiche si dissociano. Di
conseguenza i due fluorofori CFP e YFP si allontanano e FRET è abolito (fig.15).
FRET può essere facilmente misurato come il rapporto tra le intensità di
emissione del donatore (480 nm) sull’accettore (545 nm). Cambiamenti nel
rapporto 480 nm/545 nm sono direttamente correlati al grado di associazione tra le
subunità C e R della sonda basata su PKA e quindi ai livelli di cAMP.
49
Fig.15: rappresentazione schematica del sensore basato su PKA. Le subunità R e C della PKA
sono state fuse rispettivamente con CFP e YFP. A bassa concentrazione di cAMP le subunità sono
associate ed avviene il trasferimento di energia. Ad alte concentrazioni di cAMP le subunità si
separano, il trasferimento di energia non avviene più, l’emissione a 545 nm cala e perciò il
rapporto delle emissioni CFP/YFP aumenta (87).
Il secondo tipo di sonda da noi utilizzato si basa su Epac (91). Epac è un fattore
scambiatore di GTP-GDP (guanine exchange factor, GEF) che agisce su Rap1 e
Rap2. Le proteine Rap sono delle GTPasi che ciclicamente passano dalla forma
inattiva che lega GDP a quella attiva che lega GTP, attraverso l’azione di proteine
GEF che mediano lo scambio tra GDP e GTP. A partire dall’N-terminale, Epac è
costituita da un dominio regolatorio contenente un sito di legame per il cAMP
simile a quello presente nelle subunità R della PKA ed un dominio DEP che
media il legame della proteina stessa alla membrana cellulare. A questo dominio
segue un sito catalitico caratteristico della famiglia di proteine GTPasi Ras. A
bassi livelli di cAMP, Epac è ripiegata in una conformazione inattiva in cui
l’ingombro sterico impedisce il suo legame a Rap. Quando il cAMP si lega ad
Epac, questa proteina assume una conformazione estesa, associandosi così a Rap.
La sonda Epac da noi utilizzata è stata ottenuta fondendo l’ammino-terminale ed il
carbossi-terminale di Epac1 rispettivamente a CFP e a YFP (fig.16).
50
Fig.16: rappresentazione schematica del sensore basato su Epac1. I fluorofori CFP e YFP sono
stati fusi rispettivamente alle estremità N- e C-terminale di Epac1. A basse concentrazioni di
cAMP la proteina si trova nella forma inattiva ed i due fluorofori sono sufficientemente vicini
perché si abbia FRET. In seguito al legame del cAMP, Epac1 assume una conformazione estesa,
CFP e YFP si allontanano e FRET è abolita (91).
1.14 Espressione di Gs e Golf.
Ciascun neurone sensoriale olfattivo esprime diverse proteine G, in particolare
due tipi di proteine G stimolatorie, Gs e Golf, che portano all’attivazione delle
adenilato ciclasi. Golf è strettamente associata al recettore olfattivo nel processo di
trasduzione del segale odoroso. Essa è infatti prevalentemente espressa nei NSO
maturi. L’espressione di Gs, al contrario, si verifica principalmente nel corso dello
sviluppo embrionale e nei neuroni immaturi, inducendo ad ipotizzare un suo ruolo
nei processi di crescita assonale (16,17). I dati finora disponibili relativi alla
sintesi di Gs nei NSO sono limitati al periodo embrionale ed ai primi giorni di vita
post-natale. Per determinare l’espressione di tale proteina a livello del bulbo
olfattivo e valutare se essa possa svolgere un ruolo nell’axon guidance durante il
processo di rigenerazione dei neuroni sensoriali olfattivi, è stata analizzata la sua
espressione sia in ratti che in topi, non solo neonati, ma anche adulti (>PD20)
tramite saggi di immunoistochimica.
51
52
2. Materiali e metodi
2.1 Colture primarie di neuroni sensoriali olfattivi.
Colture primarie di neuroni sono ottenute da turbinati di feti di ratti Sprague
Dawley di 18-20 giorni, come descritto da Ronnett (92). La ratta viene uccisa con
eccesso di CO2 e successiva dislocazione cervicale; vengono prelevati i feti ed in
seguito decapitati.
Il cranio è inciso sagittalmente dalla regione occipitale verso quella frontale,
ottenendo due emiteste. Da queste si isolano i turbinati, utilizzando un
microscopio da dissezione (Leica MZ16F).
Tutte queste fasi di dissezione sono eseguite in HBSS (Invitrogen) a 4°C.
I turbinati sono digeriti con tripsina 2.5% a 37°C per 15 minuti. La reazione è
successivamente bloccata per aggiunta di FBS. Di seguito, si eseguono tre lavaggi
con HBSS, per rimuovere l’enzima. I turbinati sono centrifugati due volte a 4000
rpm per 5 min. Infine i tessuti sono dissociati meccanicamente con una pipetta
pasteur con diametro ridotto. Le cellule sono poi piastrate a densità variabile tra
300.000 e 600.000 cellule/vetrino su vetrini copri-oggetto da 24 mm (VWR)
trattati precedentemente con Poli-L-lisina (10µg/ml) (Sigma). I neuroni vengono
mantenuti in terreno di coltura come descritto in Ronnett (92).
Tutte le cellule utilizzate in questo studio sono chiaramente identificabili come
neuroni sensoriali olfattivi in base alla loro caratteristica morfologia bipolare, dal
momento che presentano un singolo dendrite più spesso, terminante con
un’espansione (knob), da cui protrudono le cilia, ed un assone molto lungo e
sottile. I NSO maturi in coltura esprimono il marker specifico olfactory marker
protein (OMP) (11,12).
53
2.2 Trasfezione di neuroni sensoriali olfattivi.
Il sensore per cAMP è interamente codificato geneticamente, quindi la sua
espressione nei NSO si ottiene tramite trasfezione del DNA codificante le
subunità chimeriche di PKA o Epac. Il sensore PKA è una forma modificata della
sonda originale (87), presentando una maggiore affinità per il cAMP ed un più
ampio range dinamico in risposta al cAMP (Zamparo and Zaccolo, unpublished
data).
Nei plasmidi la trascrizione dei cDNA di PKA ed Epac è sotto il controllo di un
promotore CMV che assicura un livello di espressione sufficientemente alto tale
da permettere l’esecuzione di esperimenti nella maggior parte dei tipi cellulari
utilizzati, sia in colture cellulari primarie che in linee cellulari.
Dopo 24 ore, le colture sono trasfettate utilizzando il reagente lipidico trasfettina
(BioRad), seguendo il protocollo fornito dalla ditta stessa. Per ciascun vetrino
sono utilizzati 4 µg di DNA e 5 µl di trasfettina. Dopo un’ora di incubazione a
37˚C a 5% di CO2 i neuroni sono lavati con DMEM privo di Rosso Fenolo
(Invitrogen) riscaldato a 37°C e riposti nel terreno di coltura originale arricchito
con 50% di mezzo fresco.
Le cellule così trasfettate sono utilizzate il giorno successivo per condurre gli
esperimenti di imaging di cAMP.
2.3 Imaging di cAMP in neuroni sensoriali olfattivi isolati.
Il vetrino viene montato in un’apposita cameretta termostatata a 37°C,
ermeticamente chiusa (Warner Instruments), e mantenuto in 800-900 µl di
soluzione Ringer. Le soluzioni saline contengono HEPES, per mantenere costante
il pH. Gli stimoli sono aggiunti direttamente nella soluzione salina o localmente
applicati all’assone terminale-cono di crescita per iniezione pressurizzata
(Pneumatic pico-pump, WPI) con una micropipetta (3-5 µm di diametro, durata
del puff 3 s). La preparazione è continuamente perfusa con soluzione Ringer (1.5
ml/min). La concentrazione della mix odorosa nella micropipetta è 1 mM.
54
Gli esperimenti sono condotti su un microscopio invertito (IX81, Olympus) con
obiettivo a immersione in olio 60X NA 1.4. Il microscopio è equipaggiato con una
CCD camera (SIS F-View), con un sistema di illuminazione MT20 (Olympus) e
con un beam-splitter optical device (Multispec Microimager; Optical Insights). Le
immagini sono acquisite utilizzando il software Cell^R. Per ogni esperimento si
ottengono una serie di immagini acquisite con frequenza impostata. Ad ogni
acquisizione si ottengono due immagini: una relativa al canale CFP ed una
relativa al canale YFP. Queste vengono elaborate tramite un apposito software
offline sviluppato in MATLAB 7.3. Questo programma permette di sottrarre il
rumore di fondo a ciascuna immagine, calcolare il rapporto, pixel per pixel, per
ciascun punto nel tempo tra l’immagine del canale CFP e l’immagine del canale
YFP, ed infine mettere in grafico i valori del rapporto di intensità di fluorescenza
CFP/YFP nel tempo. I dati sono ottenuti dalla media delle intensità dei pixel
all’interno di una definita area della cellula. Tracciando una regione di interesse
(Region Of Interest, ROI) in corrispondenza di aree specifiche, possono essere
stimate le cinetiche delle variazioni di cAMP nei diversi compartimenti cellulari:
cilia-dendrite, soma, assone terminale-cono di crescita. I cambiamenti di FRET
sono perciò misurati come cambiamenti nell’intensità di emissione di
fluorescenza a 480 nm/545 nm (dopo aver sottratto il background) in seguito ad
eccitazione a 430 nm ed espressi come R/R0, dove R è il rapporto al tempo t e R0
è il rapporto al tempo t0 in cui viene applicato lo stimolo. La risposta in
percentuale è valutata come ∆R/R0 x 100, con ∆R=R- R0. Il tempo per il
raggiungimento di metà della risposta massimale (t1/2) è valutato come il tempo,
dopo l’applicazione dello stimolo, al quale viene raggiunta la metà della risposta
massimale, considerando metà della risposta massimale= [(R-R0)/2] +R0 e t=t-t0.
Per l’analisi statistica dei dati è stato utilizzato il programma Excel di Microsoft
Office. Tutti i dati sono presentati come la media aritmetica±errore standard
(standard error mean, s.e.m.). Per calcolare la significatività statistica dei dati si è
effettuato un test di Student (t-test) di tipo accoppiato a due code. I valori di
significatività sono denotati come segue: *0.01<p≤0.05; **0.001<p≤0.01;
***p≤0.001. Il numero di cellule è indicato con “n”.
55
2.4 Traslocazione nucleare della subunità catalitica di PKA.
Per seguire la traslocazione nucleare della subunità catalitica della sonda PKA
associata al fluoroforo YFP, abbiamo valutato i cambiamenti in fluorescenza di
YFP (filtro di eccitazione 500 nm, filtro di emissione 545 nm) a livello nucleare in
NSO trasfettati con il sensore basato su PKA prima e dopo l’applicazione dello
stimolo (applicato al bagnetto o tramite puff). Le immagini, acquisite ogni 10
minuti, sono poi processate con il programma ImageJ (National Institute of
Health) per rilevare eventuali cambiamenti nella fluorescenza a livello del nucleo.
2.5 Ca2+ imaging in neuroni sensoriali olfattivi isolati.
Dopo 24 ore in coltura i vetrini sono trattati per caricare i neuroni sensoriali
olfattivi con l’indicatore del Ca2+ raziometrico FURA-2-AM (Invitrogen). Le
cellule vengono incubate per 35 minuti in terreno contenente FURA-2-AM
(concentrazione finale 5 µM), acido pluronico 10%, (Invitrogen) e sulfinil
pirazone (Sigma-Aldrich) (concentrazione finale 10 mM) a 37°C e 5% di CO2. I
vetrini vengono poi sciacquati con DMEM senza Rosso Fenolo per eliminare
l’eccesso di FURA-2-AM e montati nell’apposita cameretta per condurre gli
esperimenti di imaging del Ca2+. L’indicatore FURA-2-AM è caratterizzato dal
cambiamento dello spettro di assorbimento in seguito al legame con il Ca2+. Il
picco di assorbimento, infatti, si sposta da 380 nm a 340 nm. Il campione viene
perciò eccitato a 340 nm e 380 nm ed il rapporto delle emissioni a tali lunghezze
d’onda è direttamente correlato alla quantità di Ca2+ intracellulare. Tutti gli
esperimenti sono condotti in presenza di TTX (Latoxan) (concentrazione finale
4 µM), per evitare il contributo dei potenziali d’azione alla misurazione del
segnale Ca2+. E’ stato utilizzato un microscopio invertito (IX81, Olympus) con
obiettivo ad immersione in olio 40X. I cambiamenti nella concentrazione di Ca2+
intracellulare sono visualizzati utilizzando i filtri di eccitazione a 380 nm e 340
nm ed i filtri di emissione rispettivamente a 510 nm e 540 nm. Le immagini,
acquisite ogni secondo, sono poi processate tramite il programma ImageJ
(National Institute of Health). I cambiamenti nella fluorescenza (340 nm/380 nm)
56
sono espressi come R/R0, dove R è il rapporto al tempo t e R0 è il rapporto al
tempo t0 di applicazione dello stimolo. La percentuale di risposta è valutata come
∆R/R0 x 100, con ∆R=R- R0.
Tutti i dati sono presentati come la media aritmetica±errore standard (standard
error mean, s.e.m.).
Per calcolare la significatività statistica dei dati si è effettuato un test di Student (ttest) di tipo accoppiato a due code. I valori di significatività sono denotati come
segue: *0.01<p≤0.05; **0.001<p≤0.01; ***p≤0.001. Il numero di cellule è
indicato con “n”.
2.6 Ca2+ imaging ex vivo.
Gli esperimenti sono condotti su topi C57/BL6 (Charles River) tra il 8-22 giorno
post-natale (PD8-PD22). I neuroni sensoriali olfattivi vengono caricati in vivo con
l’indicatore Calcium Green dextran, 10 KDa MW (Molecular Probes), seguendo il
protocollo adattato da Wachowiack e Cohen (52). Tre-quattro giorni dopo il
caricamento dell’indicatore, i topi sono anestetizzati con Zoletil 100 (una
combinazione di Zolazapam e Tiletilamine, 1:1, 10 mg/kg, Laboratoire Virbac) e
Xilor (Xilazine 2%, 0.06 ml/kg, Bio98), decapitati e la testa è tagliata
sagittalmente (preparati di espianti di emiteste). Un’emitesta viene poi posizionata
in un’apposita cameretta per imaging, in ACSF (in mM: 119 NaCl, 1.3 MgSO4,
2.5 CaCl2·2H2O, 2.5 KCl, 1 KH2PO4, 26.2 NaHCO3, 11 glucosio) e ossigenata
con 95% O2 e 5% CO2. Un sistema confocale a scansione laser (Bio-Rad
Radiance 2100) collegato ad un microscopio Nikon Eclipse E 600-FN è utilizzato
per visualizzare i cambiamenti in fluorescenza del Calcium Green. Il sistema
confocale è dotato di un laser Argon. L’obiettivo utilizzato per gli esperimenti è
un 40X NA 0.80 (Nikon, Fluor) con immersione ad acqua. L’illuminazione di
eccitazione è di 514 nm e la fluorescenza in emissione è raccolta da un filtro HQ
545 nm/540 nm. Le immagini sono acquisite con una risoluzione di 512 X 512
pixel sotto il controllo di un software Bio-Rad-LaserSharp 2000. Tutti i dati sono
processati utilizzando un apposito software offline sviluppato in MATLAB 7.3.
57
I dati sono presentati come la media aritmetica±errore standard (standard error
mean, s.e.m.).
Per calcolare la significatività statistica dei dati si è effettuato un test di Student (ttest) di tipo accoppiato a due code. I valori di significatività sono denotati come
segue: 0.01<p≤0.05; **0.001<p≤0.01; ***p≤0.001. Il numero di animali e/o di
glomeruli è indicato con “n”.
2.7 Immunocitochimica per OMP.
Dopo 48 ore in coltura le cellule sono fissate in metanolo 100% freddo per 20 min
a -20°C. Le cellule vengono poi fatte reagire con un anticorpo policlonale
specifico per OMP (goat, anti-OMP, Wako Chemicals, USA). Il legame
dell’anticorpo primario è visualizzato utilizzando un anticorpo secondario antigoat IgG coniugato con Cy3 (Jackson Laboratories).
2.8 Immunoistochimica per Gs.
Gli animali sono perfusi con soluzione salina NaCl 0.9% ed in seguito
paraformaldeide 4% in PBS. Le teste vengono dissezionate ed i cervelli sono
mantenuti in paraformaldeide 4% over-night. In seguito i cervelli sono posti in
saccarosio 30% in PBS per 24 ore per la crioprotezione. Sezioni coronali (20µm)
del bulbo olfattivo si ottengono con un criostato microtomo (Leica CM1850) e
sono montate su vetrini Superfrost Plus (VWR). A questo punto si procede con il
saggio di immunoistochimica.Le sezioni di bulbo olfattivo sono incubate per 1 ora
in una soluzione bloccante-permeabilizzante contenente 5% donkey serum
(Jackson
Laboratories),
0.5%
Tryton
X-100
(Applichem)
in
successivamente si effettuano le incubazioni con i seguenti anticorpi:
-
anticorpo primario anti-Gsα (rabbit) (Millipore) over-night a 4°C;
58
PBS
e
-
anticorpo
secondario
(donkey
anti-rabbit)
coniugato
Cy2
(Jackson
Laboratories) per 2 ore a temperatura ambiente;
-
anticorpo primario anti-OMP (goat) (Wako Chemicals) 2 ore a temperatura
ambiente, per visualizzare i NSO nell’epitelio olfattivo;
-
anticorpo
secondario
(donkey
anti-goat)
coniugato
Cy3
(Jackson
Laboratories) per 2 ore a temperatura ambiente;
-
DAPI (Invitrogen) per la visualizzazione dei nuclei cellulari.
Le immagini vengono acquisite al sistema confocale Leica TCS SP5. L’obiettivo
utilizzato è un 40X ad immersione in olio. I laser di eccitazione utilizzati per Cy2
e Cy3 sono rispettivamente 488 nm e 561 nm, mentre i range di emissione usati
sono rispettivamente 495 nm-545 nm e 565 nm-624 nm. Il merged è stato eseguito
utilizzando il programma ImageJ (National Institute of Health).
2.9 Stimoli su neuroni sensoriali olfattivi isolati.
Gli stimoli farmacologici: forscolina (3 µM e 25 µM) e isobutil-metil-xantina
(IBMX, 100 µM), forniti da Sigma-Aldrich, sono preparati in stock e diluiti alla
concentrazione finale nel bagnetto. Lo stimolo odoroso è rappresentato dalla
mistura di diversi componenti: acetofenone, citralva, S- e R-carvone, citronella,
geraniolo, eugenolo, mentone (tutti acquistati da Sigma-Aldrich) preparati in
concentrazione 1 mM in Ringer e diluiti poi alla concentrazione finale di 200 µM
nel bagnetto (48). Prima dell’utilizzo la miscela di odori viene sonicata per
assicurare che tutti i componenti siano in soluzione. Nel caso della stimolazione
odorosa focale a livello dell’assone terminale-cono di crescita dei NSO la
concentrazione della mix di odori nella micropipetta è pari a 1 mM.
Gli inibitori SQ22536 (30 µM) e MgCl2 (10 mM) sono forniti rispettivamente da
Sigma-Aldrich e da Biomol. Internat.
59
2.10 Stimoli su preparati di emiteste.
Per evitare il contributo di potenziali d’azione al segnale Ca2+ misurato, TTX 4
µM è stata aggiunta alla soluzione di ACSF nella cameretta, almeno 5 min prima
dell’inizio dell’esperimento. Gli stimoli sono somministrati per iniezione
pressurizzata (Pneumatic pico-pump, WPI) con una micropipetta (3-5 µM di
diametro) posizionata sopra i glomeruli (distanza ~40 µm) nel bulbo olfattivo. La
concentrazione di stimolo in pipetta è: 250 µM per la forscolina, 1 mM per la mix
di odori, 500 µM per l’inibitore delle AC, SQ22536. La durata del puff è di 60 s.
L’incremento in [Ca2+] indotto da forscolina od odori inizia dopo 1-3 s
dall’applicazione del puff sul BO e quindi esclude che questo effetto sia dovuto
alla diffusione della mix odorosa verso le cilia dei NSO, che sono localizzati a
circa 1 cm di distanza.
2.11 Terreni e soluzioni utilizzate.
I tessuti, durante la preparazione delle colture cellulari sono mantenuti in HBSS
1X (HBSS, HEPES 0.3M, Pen-Strep 100U/ml).
Le cellule sono mantenute in terreno di coltura come descritto in Ronnett (92)
(MEM- D-valina; FBS 10%; NU Serum 5%; PenStrep L-glutamine 100 U/ml,
AraC 10 µM, NGF 25 ng/ml) a 37˚C a 5% di CO2.
Gli esperimenti di imaging sono stati condotti incubando le cellule in soluzione
Ringer (in mM: 140 NaCl, 5 KCl, 1 CaCl2·H2O, 1 MgCl2, 10 glucosio, 1 sodio
piruvato, 10 HEPES; pH 7.2). Per gli esperimenti condotti in assenza di calcio
esterno, si è utilizzata una soluzione Ringer senza calcio (in mM: 140 NaCl, 5
Kcl, 1 MgCl2, 10 glucosio, 1 sodio piruvato, 10 HEPES; pH 7.2) e con EGTA
1mM.
Il MEM, il DMEM senza Rosso Fenolo, il FBS provengono dall’Invitrogen
Corporation (Paisley, UK). Il NGF e il NU Serum sono acquistati dalla BD
Bioscience. L’AraC proviene dalla Sigma-Aldrich.
60
Il PBS è stato preparato in concentrazione 10X (in mM: 137 NaCl, 27 KCl, 101.4
Na2HPO4, 17.6 KH2PO4; pH 7.4) e filtrato. Tutti i componenti sono acquistati da
Sigma-Aldrich.
La paraformaldeide 4% (Sigma-Aldrich) è stata sciolta in PBS 1X, pH 7.4.
61
62
3. Risultati
3.1 Progetto di ricerca.
Il lavoro oggetto di questa tesi è volto a determinare le proprietà funzionali del
recettore olfattivo espresso sul cono di crescita-assone terminale dei neuroni
sensoriali olfattivi e a stabilire la cascata di signalling intracellulare associata a
tale recettore. Lo studio è stato condotto inizialmente in vitro su colture primarie
di neuroni sensoriali olfattivi.
3.2 Colture primarie di neuroni sensoriali olfattivi.
Colture primarie di neuroni sensoriali olfattivi sono state ottenute da turbinati di
feti di ratto Sprague Dawley. Abbiamo verificato che per ottenere una coltura
ottimale per numero e morfologia di NSO, gli embrioni devono avere un’età
compresa tra i 18 ed i 20 giorni. Al di sotto di tale età si osservava nelle colture di
neuroni sensoriali olfattivi una maggior contaminazione da parte di altri tipi di
cellule epiteliali, mentre le preparazioni cellulari ottenute da feti di età superiore ai
20 giorni (e quindi anche da ratti neonati) risultavano più resistenti alla trasfezione
con i sensori per il cAMP. Per verificare che le cellule messe in coltura fossero
neuroni sensoriali olfattivi maturi, abbiamo eseguito saggi di immunocitochimica
(fig.17), utilizzando come marker specifico per il nostro tipo cellulare la proteina
OMP (olfactory marker protein) (vedi §1.4).
63
Fig. 17:
immagine di un neurone sensoriale olfattivo in coltura marcato per OMP. Si può
osservare la tipica morfologia bipolare.
3.3 Trasfezione di neuroni sensoriali olfattivi in coltura.
Per valutare le dinamiche spazio-temporali del cAMP, i neuroni sensoriali
olfattivi sono stati trasfettati con le sonde per il cAMP.
Inizialmente abbiamo testato due tipi di sensori, il primo basato sulla proteina
chinasi A (PKA) ed il secondo sulla proteina Epac (vedi §1.13). Tuttavia la
minore efficienza di trasfezione nel nostro modello cellulare e l’inferiore
sensibilità per il cAMP presentate dalla sonda basata su Epac rispetto alla sonda
basata su PKA (la Kd per il cAMP è rispettivamente di ~50 µM vs ~300 nM) (91,
Zamparo & Zaccolo, unpublished data), ci hanno portato a scegliere quest’ultima.
Tutti gli esperimenti descritti di seguito sono stati pertanto condotti con la sonda
basata su PKA.
Come si può osservare dalla fig.18 i neuroni sensoriali olfattivi trasfettati con il
sensore basato su PKA mantengono la normale morfologia bipolare. La
fluorescenza della sonda è omogeneamente distribuita nel citosol, mentre è
esclusa dal nucleo.
64
Fig.18: neurone sensoriale olfattivo in coltura trasfettato con la sonda basata su PKA. La
fluorescenza è distribuita lungo tutto il citoplasma con esclusione del nucleo.
3.4 Dinamiche spazio-temporali di cAMP in neuroni sensoriali
olfattivi isolati.
Una volta trasfettate, le cellule venivano mantenute in condizioni standard di
coltura per 24 ore ed in seguito utilizzate in esperimenti di real time imaging per
valutare le dinamiche spazio-temporali del cAMP.
3.4.1 Effetto dello ione Cl- sull’emissione di fluorescenza di YFP e
CFP.
Inizialmente negli esperimenti di real time imaging abbiamo testato due tipi
diversi di soluzioni saline extracellulari, Ringer e HBSS-gluconato, che si
differenziavano rispettivamente per la presenza o l’assenza di Cl-. E’ noto infatti
che tale anione può provocare un quenching del fluoroforo YFP in alcuni tipi
cellulari (93).
Abbiamo pertanto effettuato dei controlli preliminari per verificare che la
presenza di tale ione nella soluzione Ringer non interferisse con l’emissione di
YFP. I NSO sono stati trasfettati contemporaneamente con due plasmidi,
codificanti esclusivamente per i fluorofori YFP e CFP, non coniugati con le
subunità catalitica e regolatoria della PKA. Dopo 24 ore, le cellule sono state
utilizzate per gli esperimenti di real time imaging. I NSO sono stati trattati con
stimoli farmacologici in grado di aumentare la concentrazione di cAMP, quali la
65
forscolina (FRSK), un attivatore generico delle adenilato ciclasi, e l’isobutilmentil-xantina (IBMX), un inibitore aspecifico delle fosfodiesterasi. Gli stimoli
farmacologici sono stati aggiunti direttamente al bagnetto sperimentale. Le
emissioni del canale CFP e del canale YFP (480 nm/545 nm) sono state
monitorate a livello di: cilia-dendrite, soma, assone terminale-cono di crescita per
tutta la durata dell’esperimento (10-15 minuti). Dopo il trattamento con forscolina
l’intensità delle emissioni dei fluorofori YFP e CFP (e pertanto il loro rapporto) si
sono mantenute stabili nell’intero NSO. La successiva somministrazione di IBMX
non ha prodotto alcuna variazione del segnale (fig.19). Anche quando i neuroni
sensoriali olfattivi sono stati stimolati in primo luogo con IBMX e
successivamente con forscolina, il rapporto dell’intensità delle emissioni dei due
fluorofori è rimasto costante.
Fig.19: cinetica dei cambiamenti del rapporto di intensità di emissione di CFP e YFP (480 nm/545
nm) in un NSO stimolato con forscolina (25 µM) e successivamente con IBMX (100 µM) in
soluzione Ringer.
Questi risultati indicavano che nel nostro modello cellulare il Cl- non provocava
un quenching della fluorescenza di YFP. Tale controllo preliminare è risultato
fondamentale per escludere che nei successivi esperimenti di real time imaging
delle dinamiche di cAMP, le variazioni del rapporto dell’intensità di emissione tra
CFP e YFP fossero dovute ad alterazioni della fluorescenza dei singoli fluorofori,
indipendenti da effettivi cambiamenti di concentrazione di cAMP. Tutti gli
esperimenti riportati in seguito sono stati pertanto condotti utilizzando la
soluzione Ringer, che, rispetto alla soluzione HBSS-gluconato priva di Cl-,
presenta maggiori caratteristiche fisiologiche per i NSO.
66
3.4.2 Dinamiche spazio-temporali di CAMP in neuroni sensoriali
olfattivi trattati con stimoli farmacologici.
Per valutare le dinamiche spazio-temporali di cAMP nell’intero NSO, abbiamo
trattato le cellule con stimoli in grado di aumentare la concentrazione di cAMP,
quali FRSK e IBMX. Gli stimoli sono stati aggiunti direttamente nel bagnetto
sperimentale. L’incremento della concentrazione intracellulare di cAMP in
seguito a stimolazione è stato valutato come aumento del rapporto dell’intensità di
emissione del canale CFP e del canale YFP (480 nm /545 nm). Tali variazioni
sono state esaminate a livello di: cilia-dendrite, soma, assone terminale-cono di
crescita.
In seguito al trattamento con forscolina (3 µM) abbiamo rilevato un aumento di
cAMP in tutto il neurone sensoriale olfattivo, compreso l’assone terminale-cono
di crescita. Abbiamo quindi analizzato le dinamiche temporali dell’incremento di
cAMP (t1/2, tempo necessario per raggiungere la metà della risposta massimale
misurata) a livello di cilia-dendrite, soma, assone terminale-cono di crescita nei
NSO. Come si può osservare dal grafico riportato in fig.20, a seguito di
stimolazione con forscolina l’aumento di cAMP si verificava più rapidamente
nelle cilia-dendrite e nell’assone terminale-cono di crescita rispetto al soma (
n=13, t1/2 in s: cilia-dendrite 46.8±13.0; soma 92.2±14.5; assone terminale-cono di
crescita 41.7±5.8; t-test: cilia-dendrite/soma p=0.004**; assone terminale-cono di
crescita/soma p=0.002**; cilia-dendrite/cono di crescita p=0.15). Gli stessi
neuroni sensoriali olfattivi trattati con forscolina sono stati trattati in un secondo
momento con IBMX (100 µM). Il secondo stimolo farmacologico non ha prodotto
ulteriori incrementi di cAMP. Questo risultato potrebbe essere ascrivibile ad una
saturazione della sonda PKA.
67
a
b
Fig.20: a Cinetica dei cambiamenti del rapporto di intensità di emissione (480 nm/545 nm)
registrati in cilia-dendrite (blu), soma (viola), assone terminale-cono di crescita (verde), in un NSO
stimolato con forscolina (3 µM) e successivamente con IBMX (100 µM).. b Media dei tempi di
risposta emimassimale (t½) calcolati per le cellule trattate con FRSK (n=13, dendrite-soma
p=0.004**, cono di crescita-soma p=0.002**); barre di errore=s.e.m. *0.01<p≤0.05;
**0.001<p≤0.01; ***p≤0.001. La regione di interesse, ROI, è stata tracciata sulla parte distale
dell’assone e del dendrite e sul soma. Le cilia sono state incluse nella ROI sul dendrite, ma
risultavano troppo sottili per essere chiaramente identificabili all’ingrandimento utilizzato.
Quando i neuroni sensoriali olfattivi sono stati trattati con IBMX (100 µM) come
primo stimolo si è rilevato un incremento della concentrazione di cAMP in tutto il
NSO, incluso l’assone terminale, e nuovamente tali variazioni si verificavano più
velocemente a livello delle cilia-dendrite e dell’assone terminale-cono di crescita
rispetto al corpo cellulare (n=10; t1/2 in s: cilia-dendrite 174.4±46.3; soma
228.8±38.5; assone terminale-cono di crescita 133.6±35.7; t-test: ciliadendrite/soma p=0.04*; assone terminale-cono di crescita/soma p=0.007**; ciliadendrite/assone terminale-cono di crescita p=0.17) (fig.21). In circa un terzo delle
cellule stimolate con IBMX, la successiva somministrazione di forscolina (25
µM) ha indotto un’ulteriore aumento del segnale (fig.21a).
a
b
Fig.21 :a Cinetica dei cambiamenti del rapporto di intensità di emissione (480 nm/545 nm) in un
NSO stimolato con IBMX (100 µM). b Media dei t ½ calcolati per le cellule trattate con IBMX
(n=10; dendrite-soma p=0.04*, cono di crescita-soma p=0.007**). Barre=s.e.m.
Da ultimo, i neuroni sensoriali olfattivi sono stati trattati con FRSK e IBMX
somministrati nello stesso momento. Anche in questo caso si è osservato un
68
aumento di cAMP in tutto il neurone sensoriale, incluso l’assone terminale-cono
di crescita. Le dinamiche temporali in queste condizioni sperimentali si sono
mantenute invariate: il segnale di cAMP aumentava più velocemente ai neuriti,
cilia-dendrite e assone terminale-cono di crescita, rispetto al soma (n=10; t1/2 in s:
cilia-dendrite 56.1±6.4; soma 87.0±14.5; assone terminale-cono di crescita
51.3±11.4; t-test: cilia-dendrite/soma p=0.02*; assone terminale-cono di
crescita/soma p=0.001**; cilia-dendrite/assone terminale-cono di crescita p=0.6).
I dati ottenuti indicano quindi che:
1) l’aumento di cAMP a livello dell’assone terminale-cono di crescita dei NSO è
dovuto alla sintesi locale di cAMP e non può essere attribuito ad una eventuale
diffusione del secondo messaggero dal dendrite apicale o dal soma;
2) le adenilato ciclasi nei NSO presentano un’attività basale costitutiva e la
concentrazione basale di cAMP viene mantenuta bassa dall’attività delle
fosfodiesterasi;
3) le fosfodiesterasi non sembrano essere coinvolte nelle diversità dei t1/2
riscontrate nei vari compartimenti dei NSO.
Questi risultati dimostrano che gli elementi molecolari necessari per la sintesi e
l’idrolisi del cAMP sono presenti e funzionali non solo a livello delle cilia, come
noto, ma anche a livello dell’assone terminale-cono di crescita.
3.4.3 Dinamiche spazio-temporali di cAMP in neuroni sensoriali
olfattivi in seguito a stimolazione odorosa.
Abbiamo quindi studiato se il recettore olfattivo sull’assone terminale-cono di
crescita è funzionalmente accoppiato alla sintesi di cAMP. A tale scopo, i neuroni
sensoriali olfattivi sono stati stimolati con una soluzione contenente molteplici
molecole odorose (mix odorosa). Ciascun componente della mix odorosa
presentava gruppi funzionali differenti, in modo da aumentare le probabilità di
evocare risposte nei neuroni sensoriali testati, tenendo conto della specificità
molecolare di ogni recettore.
E’ importante sottolineare che non era ancora noto se i RO sul cono di crescita
fossero in grado di legare gli odori ed in caso affermativo quale fosse la via di
signalling intracellulare ad essi associata.
69
Inizialmente abbiamo valutato la concentrazione minima della mix odorosa
necessaria per ottenere delle risposte che mostrassero un buon rapporto
segnale/rumore negli esperimenti di imaging sia del cAMP che del Ca2+. A tal fine
abbiamo eseguito esperimenti preliminari di imaging del Ca2+ (caricando i NSO
con il FURA-2-AM) e successivamente di FRET imaging, testando un ampio
range di concentrazioni (1 mM-10 µM). La concentrazione di 200 µM è risultata
essere la concentrazione più bassa a cui si potevano ottenere risposte riproducibili
sia negli esperimenti di imaging del Ca2+ (fig.22) che in quelli di FRET imaging.
Tale valore di concentrazione, inoltre, rientra ampiamente nel range normalmente
utilizzato in esperimenti su NSO isolati (48).
a
b
Fig.22: cinetiche dei cambiamenti del rapporto di intensità di emissione (340 nm/380 nm) in un
NSO caricato con l’indicatore del Ca2+ FURA-2-AM e stimolato con mix odorosa: (a) 1mM; (b)
200 µM. In questo caso è stata disegnata un’unica ROI sull’intero NSO.
Nei NSO trasfettati con la sonda per il cAMP e trattati con mix odorosa (aggiunta
direttamente al bagnetto) si osservava un consistente aumento di cAMP in tutto il
neurone sensoriale olfattivo, incluso l’assone terminale (fig.23). Tale incremento
si verificava con dinamiche temporali analoghe a quelle osservate nei NSO trattati
con gli stimoli farmacologici. L’incremento di cAMP avveniva infatti più
rapidamente nelle cilia-dendrite e nel cono di crescita-assone terminale rispetto al
soma (n=12; t1/2 in s: cilia-dendrite 201.5±33.3; soma 337.1±54.7; assone
terminale-cono di crescita 168.3±29.7; t-test: cilia-dendrite/soma p=0.02*; assone
terminale-cono di crescita/soma p=0.006**; cilia-dendrite/assone terminale-cono
di crescita p=0.31) (fig.23).
70
a
b
Fig.23: a cinetica dei cambiamenti del rapporto di intensità di emissione (480 nm/545 nm) in un
NSO stimolato con mix odorosa (200 µM). b Media dei t ½ calcolati per le cellule trattate con mix
odorosa (n=12; dendrite-soma p=0.02*, cono di crescita-soma p=0.006**). Barre=s.e.m.
Per escludere che l’aumento di cAMP a livello del cono di crescita potesse essere
una conseguenza dell’attivazione Ca2+-dipendente delle adenilato ciclasi ivi
presenti, dovuto all’incremento della concentrazione intracellulare di Ca2+
prodotto dall’attivazione del RO sulle cilia, abbiamo stimolato i neuroni sensoriali
olfattivi con la mix odorosa utilizzando una soluzione salina extracellulare priva
di Ca2+ (Ringer Ca2+-free).
Anche in queste condizioni sperimentali si è potuto rilevare un sensibile aumento
di cAMP in tutto il NSO, incluso l’assone terminale-cono di crescita. Le
dinamiche temporali registrate in presenza di soluzione Ringer senza Ca2+ sono
risultate analoghe a quelle osservate per i neuroni immersi in soluzione Ringer
normale. Infatti, le variazioni di cAMP si verificavano prima ed in tempi
comparabili a livello di cilia-dendrite e cono di crescita-assone terminale, e
successivamente a livello del soma (n=10; t1/2 in s: cilia-dendrite 250.5±43.9;
soma 341.8±39.7; assone terminale-cono di crescita 218.5±57.2; t-test: ciliadendrite/soma p=0.004**; assone terminale-cono di crescita/soma p=0.002**;
cilia-dendrite/assone terminale-cono di crescita p=0.2) (fig.24).
71
a
b
Fig. 24: a Cinetica dei cambiamenti del rapporto di intensità di emissione (480 nm/545 nm) in un
NSO stimolato con mix odorosa (200 µM) in soluzione Ringer priva di Ca2+. b Media dei t ½
calcolati per le cellule trattate con mix odorosa in soluzione Ringer Ca2+-free (n=10; dendrite-soma
p=0.004**, cono di crescita-soma p=0.002**). Barre=s.e.m.
I dati ottenuti dimostrano che il recettore olfattivo espresso al cono di crescita dei
NSO è un recettore funzionale, in grado di legare gli odori e accoppiato a sintesi
locale di cAMP.
15
∆F
20 µm
0
Fig.25: sequenza di immagini in pseudocolori, raffiguranti le dinamiche spazio-temporali di
incremento di cAMP in un NSO stimolato con mix odorosa (200µM). La freccia bianca in alto a
sinistra indica l’assone terminale-cono di crescita del NSO, il prolungamento in basso a destra è il
dendrite. La barra sul lato destro della sequenza di immagini rappresenta la scala di pseudocolori
della differenza di intensità di fluorescenza (∆F, in unità arbitrarie), calcolata tra il valore
massimale della risposta raggiunto dopo lo stimolo e le condizioni basali.
Per valutare in modo più diretto la funzionalità del recettore olfattivo a livello del
cono di crescita, abbiamo applicato direttamente la mix odorosa (1 mM) al
terminale assonico di NSO con una micropipetta di vetro. In tali condizioni
72
sperimentali
l’incremento
della
concentrazione
di
cAMP
era
presente
esclusivamente all’assone terminale-cono di crescita (n=10; t1/2 in s: 67.5±18.3)
(fig.26a). Negli altri compartimenti subcellulari non si osservavano apprezzabili
cambiamenti nel segnale.
a
b
Fig.26: a Cinetica dei cambiamenti del rapporto di intensità di emissione (480 nm/545 nm) in un
NSO stimolato con un puff di mix odorosa (1 mM in micropipetta) localizzato all’assone
terminale-cono di crescita. b Esempio di un NSO non responsivo ad un puff odoroso applicato
all’assone terminale, ma responsivo alla forscolina (25 µM), aggiunta al bagnetto sperimentale.
E’ importante sottolineare che, mentre tutti i neuroni trattati con gli stimoli
farmacologici mostravano un incremento della concentrazione di cAMP, la
risposta alla mix odorosa era osservabile solamente in una subpopolazione di
neuroni (circa il 30% delle cellule stimolate), come ci si attendeva considerata la
specificità del RO espresso da ogni singolo NSO (fig.26b).
3.5 Dinamiche spazio-temporali di Ca2+ in neuroni sensoriali
olfattivi isolati.
L’esposizione delle cilia agli odori causa un incremento locale di cAMP seguito
da un influsso di Ca2+ attraverso i canali CNG attivati. Per valutare se questa
stessa sequenza di eventi molecolari si verificasse anche a livello del cono di
crescita, abbiamo effettuato esprimenti di Ca2+ imaging su neuroni sensoriali
olfattivi caricati con l’indicatore del Ca2+ FURA-2-AM. Lo stimolo odoroso è
stato somministrato direttamente all’assone terminale-cono di crescita tramite una
micropipetta di vetro. Tutti gli esperimenti di imaging del calcio sono stati
73
condotti in presenza di TTX, per escludere il contributo del potenziale d’azione al
segnale Ca2+ misurato.
Abbiamo osservato che un “puff” di odori applicato localmente all’assone
terminale-cono
di
crescita
induceva
un
significativo
incremento
della
concentrazione di Ca2+ esclusivamente al cono di crescita (n=15; ∆R/R0 (%)=
102.6±16.7) (fig27).
Fig.27: cinetica dei cambiamenti del rapporto di intensità di emissione (340 nm/380 nm) in un
NSO caricato con l’indicatore del Ca2+ FURA-2-AM e stimolato con un puff di mix odorosa (1
mM in micropipetta) localizzato all’assone terminale-cono di crescita.
Per verificare se l’influsso di Ca2+ registrato fosse dovuto all’apertura dei canali
CNG indotta dal cAMP, gli esperimenti sono stati ripetuti in presenza di un
bloccante di tali canali, quale il MgCl2 (94).
Il protocollo seguito per tali esperimenti è il seguente: un NSO era inizialmente
stimolato con un puff odoroso per verificare la responsività della cellula allo
stimolo. In caso di risposta, la cellula veniva abbondantemente sciacquata con
soluzione Ringer e poi si attendevano 15 minuti prima di stimolare nuovamente lo
stesso NSO, questa volta in presenza di MgCl2 (10 mM), aggiunto almeno 10
minuti prima dell’inizio del secondo esperimento.
Come si può osservare dalla fig.28, in seguito al trattamento con MgCl2, si ha
l’abolizione dell’incremento di Ca2+ associato alla presentazione dello stimolo
odoroso (n=10; p=0.002**).
74
a
b
c
Fig.28: cinetica dei cambiamenti del rapporto di intensità di emissione (340 nm/380 nm) in un
NSO caricato con FURA-2-AM e stimolato localmente sul cono di crescita con puff odoroso (a).
Lo stesso neurone è stato sciacquato e stimolato nuovamente con un puff odoroso in presenza del
bloccante dei canali CNG MgCl2 (10 mM) (b). c. Media della percentuale di risposta alla mix
odorosa delle cellule calcolata prima (in nero) e dopo (in rosso) l’applicazione dell’inibitore MgCl2
dei canali CNG (n=10; p=0.002**). Barre=s.e.m.
Dal momento che il MgCl2 non rappresentava un inibitore selettivo dei canali
CNG (e non sono disponibili bloccanti altamente selettivi per i canali CNG),
abbiamo deciso di eseguire un controllo più stringente per verificare il ruolo di tali
canali sull’influsso di Ca2+ rilevato dopo stimolazione odorosa del cono di
crescita. A questo scopo abbiamo deciso di bloccare a monte l’attivazione dei
canali CNG, inibendo la sintesi di cAMP con l’inibitore delle adenilato ciclasi
SQ22536. Il protocollo seguito è analogo a quello descritto in precedenza: dopo
aver verificato la responsività di un NSO con un puff di odori, la cellula veniva
abbondantemente sciacquata e dopo un’attesa di 15 minuti lo stesso neurone era
stimolato una seconda volta in presenza di SQ22536 (30 µM), aggiunto almeno 10
minuti prima dell’inizio del nuovo esperimento.
Come si può osservare in fig.29, bloccando l’attività delle adenilato ciclasi, si ha
l’abolizione dell’influsso di Ca2+ accoppiato alla stimolazione odorosa (n=6;
p=0.001**).
a
b
c
Fig.29: cinetica dei cambiamenti del rapporto di intensità di emissione in un NSO caricato con
FURA-2-AM e stimolato localmente sul cono di crescita con puff odoroso (a). Lo stesso neurone
è stato sciacquato e stimolato nuovamente con un puff odoroso in presenza dell’inibitore delle AC
SQ22536 (b). c. Media della percentuale di risposta alla mix odorosa delle cellule calcolata prima
(in nero) e dopo (in rosso) l’applicazione dell’inibitore SQ22536 (n=6; p=0.001**). Barre=s.e.m.
75
Per escludere che la mancata risposta allo stimolo fosse dovuta ad una
desensitizzazione del recettore olfattivo abbiamo effettuato due tipi di controlli: i)
in un primo caso abbiamo stimolato un NSO con un puff di odori,
successivamente la stessa cellula è stata sciacquata abbondantemente e dopo 15
minuti a riposo si è eseguita una nuova stimolazione. Abbiamo visto che la
seconda risposta era sostanzialmente simile alla prima per dinamiche temporali ed
ampiezza del segnale. Questo risultato è stato confermato anche in seguito
all’applicazione di più di due puff odorosi allo stesso cono di crescita. ii) In un
secondo controllo abbiamo stimolato il terminale assonico-cono di crescita con la
mix odorosa in presenza di inibitore. Nessun incremento del segnale Ca2+ si è
verificato in queste condizioni sperimentali. Lo stesso neurone è stato quindi
abbondantemente sciacquato, lasciato a riposo per almeno 15 minuti e stimolato
nuovamente con un puff odoroso in assenza di inibitore. In questo caso si
riscontrava un incremento del segnale Ca2+ (in una percentuale di cellule
comparabile a quella ottenuta con stimoli odorosi, intorno al 30%) (dati non
mostrati). Va inoltre notato che, a differenza del segnale cAMP, il segnale Ca2+
raggiunge il massimo incremento pochi secondi dopo l’aggiunta degli odori e si
inattiva rapidamente (vedi Discussione, cap.4).
L’insieme di questi risultati indica che, a seguito di stimolazione con odori
applicati focalmente al cono di crescita, si assiste ad un incremento della
concentrazione di Ca2+ esclusivamente al terminale assonico. Questo segnale Ca2+
è dovuto all’ingresso del catione in seguito all’apertura dei canali CNG, regolata
dall’aumento dei cAMP all’interno del cono di crescita. Infatti l’inibizione della
sintesi di cAMP a seguito dell’applicazione di un bloccante dell’adenilato ciclasi
(SQ22536) abolisce l’incremento di Ca2+ associato alla risposta agli odori.
3.6 Traslocazione nucleare della subunità catalitica di PKA.
I recettori olfattivi giocano un ruolo chiave nella convergenza assonale dei NSO
verso i glomeruli target a livello del bulbo olfattivo, ma non sono gli unici
determinanti. E’ stato dimostrato, infatti, che NSO esprimenti RO differenti co-
76
esprimono diversi tipi e/o diversi livelli di molecole guida, i.e. i recettori Eph e
l’efrina-5A (68). Tuttavia, la connessione tra l’identità del recettore olfattivo
presente al cono di crescita dei NSO e l’espressione di specifiche molecole guida
rimane oscuro. Recenti studi hanno mostrato che il segnale cAMP derivante
dall’attivazione del RO è in grado di modulare l’espressione di geni codificanti
per molecole guida nella convergenza assonale (84).
Per regolare l’espressione genica, il segnale cAMP associato al RO deve in
qualche modo raggiungere il nucleo dei NSO, ad esempio attraverso la
traslocazione della subunità catalitica della PKA, suo principale target.
Per valutare se l’incremento di cAMP derivante dall’attivazione del recettore
olfattivo fosse seguito dalla traslocazione nucleare di PKA-C, abbiamo monitorato
la distribuzione della subunità catalitica C-YFP della sonda PKA trasfettata nei
NSO a seguito di stimoli in grado di aumentare la concentrazione di cAMP, quali
la forscolina.
Come si può osservare in fig.30, mentre prima della stimolazione la subunità
catalitica della PKA era chiaramente esclusa dal nucleo, dopo 10 minuti
dall’aggiunta di forscolina (25 µM) nel bagnetto sperimentale una frazione di
PKA-C traslocava nel nucleo dei NSO.
Fig.30: traslocazione nucleare della subunità catalitica C-YFP della sonda PKA: condizioni basali
(t0), e dopo 10 min e 20min dall’aggiunta di forscolina (25 µM) al bagnetto sperimentale. Le
lunghezze d’onda di eccitazione e di emissione sono rispettivamente 500 nm e 545 nm. A queste
lunghezze d’onda solo YFP viene eccitato; la fluorescenza, quindi, è sensibile solamente alla
concentrazione/distribuzione di YFP ed indipendente da cAMP.
77
In un secondo set di esperimenti i NSO sono stati trattati con la miscela odorosa
(200 µM), aggiunta al bagnetto sperimentale. Abbiamo osservato che 10 minuti
dopo l’aggiunta di odori al NSO, la subunità catalitica della PKA era presente nel
nucleo del NSO (dati non mostrati).
Per valutare se l’attivazione selettiva del recettore olfattivo all’assone terminalecono di crescita fosse seguita da traslocazione nucleare della PKA-C abbiamo
stimolato direttamente il terminale assonico di NSO con un puff di odori (1 mM
nella micropipetta). Analogamente a quanto osservato in precedenza, si è rilevata
una traslocazione nucleare della subunità catalitica della PKA dopo 10 minuti
dalla somministrazione del puff odoroso (fig.31).
Fig.31: traslocazione nucleare della subunità catalitica C-YFP della sonda PKA: condizioni basali
(t0), e dopo 10 min e 20min dall’applicazione focale di un puff odoroso (1 mM in micropipetta)
all’assone terminale-cono di crescita di un NSO. Sono visibili alcuni aggregati di PKA-C a livello
dei neuriti, come osservato in circa il 40% delle cellule trasfettate.
Per verificare che i cambiamenti di fluorescenza osservati nel nucleo non fossero
ascrivibili ad artefatti, abbiamo eseguito due tipi di controlli: i) in un primo caso è
stato aggiunto al bagnetto sperimentale un volume di soluzione Ringer pari al
volume di mix odorosa (stimolo) normalmente utilizzata negli esperimenti
precedenti; ii) nel secondo controllo, un puff di soluzione Ringer è stato applicato
direttamente all’assone terminale di NSO (fig.32). In entrambi i casi non si sono
rilevati cambiamenti significativi della fluorescenza a livello del nucleo, per tutta
la durata dell’esperimento (almeno 30 minuti).
78
Fig.32: controllo della traslocazione nucleare della subunità catalitica C-YFP della sonda PKA:
condizioni basali (t0), dopo 10 min e 20 min dall’applicazione di un puff di Ringer all’assone
terminale-cono di crescita di un NSO.
I risultati ottenuti indicano che l’attivazione del RO sul cono di crescita è
associata a traslocazione nucleare di PKA-C.
3.7 Dinamiche spazio-temporali del Ca2+ ex vivo.
Gli esperimenti condotti su colture di neuroni sensoriali olfattivi hanno dimostrato
che il recettore olfattivo a livello del cono di crescita è funzionale, capace di
legare gli odori e di attivare una cascata di signalling intracellulare locale, che
coinvolge il cAMP ed il Ca2+.
Per valutare se i risultati ottenuti in vitro non fossero dovuti alle condizioni di
coltura ed all’isolamento dei neuroni, ma fossero presenti nel bulbo olfattivo
intatto, abbiamo effettuato esperimenti di imaging del Ca2+ sul sistema olfattivo
integro, utilizzando particolari preparati, definiti espianti di emiteste. In questi
preparati la testa di un topo è stata tagliata sagittalmente e le connessioni tra
l’epitelio olfattivo, il bulbo ed il cervello sono state mantenute lungo l’asse
mediale. I neuroni sensoriali olfattivi sono stati caricati con gli indicatori del Ca2+
(Calcium Green dextran) per semplice inalazione in topi vivi anestetizzati. Gli
animali erano quindi mantenuti in condizioni standard di stabulazione per 2-3
giorni, per permettere il completo caricamento dei NSO. I neuroni così caricati
presentavano una forte ed omogenea fluorescenza dalle cilia fino all’assone
79
terminale, per cui gli stessi glomeruli risultavano fortemente marcati. Il giorno
dell’esperimento i topi venivano decapitati e la testa era tagliata sagittalmente
(emitesta). L’emitesta era quindi posizionata in una cameretta da imaging fatta su
misura, immersa in ACSF e costantemente ossigenata con 95%O2 e 5% CO2. Tutti
gli esperimenti sono stati condotti in presenza di TTX (4 µM nel bagnetto) per
evitare il contributo del potenziale d’azione alla misura del Ca2+ intracellulare. Gli
stimoli sono stati somministrati localmente sopra i glomeruli marcati con una
micropipetta di vetro. L’aumento di concentrazione di Ca2+ è stato valutato come
aumento dell’intensità di emissione dei glomeruli a 545 nm.
Analogamente alla procedura seguita negli esperimenti in vitro, abbiamo
dapprima stimolato i glomeruli con la forscolina. Un puff di forscolina applicato
direttamente ai glomeruli con una micropipetta di vetro induceva un evidente
incremento di fluorescenza (indicante un aumento della concentrazione di Ca2+)
dei glomeruli caricati (topi n=4, glomeruli n=20; ∆F/F0=0.38±0.03) (fig.33a).
80
a
b
c
d
e
f
Fig. 33: cinetiche del cambiamento di fluorescenza (∆F/F0) nei terminali assonici di neuroni
sensoriali olfattivi in preparati di emiteste in seguito alla stimolazione con forscolina (a) e con mix
odorosa (b). In (c) e (d) immagini di glomeruli caricati con il Calcium Green: prima (a sinistra) e
dopo (a destra) applicazione di frsk (c) e odori (d). In (e) e (f) le immagini sono presentate in
pseudocolori . La barra sulla destra dei pannelli indica la scala di pseudocolori della differenza di
intensità di fluorescenza (∆F) (in unità arbitrarie) tra il picco della risposta e le condizioni basali
(e) forscolina e (f) mix odorosa. Le frecce indicano la posizione della micropipetta; i cerchi e le
linee colorate indicano i glomeruli.
Per valutare in vivo l’accoppiamento funzionale del recettore olfattivo
all’incremento della concentrazione di cAMP, abbiamo voluto verificare se la
stimolazione odorosa dei terminali assonici di NSO applicata direttamente a
livello dei glomeruli era in grado di generare un incremento di Ca2+.
Stimolando i glomeruli con un puff di odori si è rilevato un rapido incremento di
Ca2+ nei terminali pre-sinaptici a livello dei glomeruli (topi n=9, glomeruli n=23,
∆F/F0=0.44±0.06) (fig.33b).
Per verificare se tale segnale Ca2+ fosse dovuto all’apertura dei canali CNG
indotta dal cAMP, abbiamo bloccato la produzione di cAMP con l’inibitore delle
adenilato ciclasi SQ22536. Analogamente alla procedura seguita negli esperimenti
su NSO isolati, abbiamo dapprima verificato la responsività dei glomeruli testati
con un puff odoroso; in caso di risposta, i glomeruli venivano abbondantemente
81
sciacquati e dopo un attesa di 15 minuti un puff odoroso era nuovamente applicato
agli stessi glomeruli, questa volta in presenza dell’inibitore delle AC (aggiunto
almeno 10 minuti prima dell’inizio dell’esperimento). In presenza del bloccante
l’incremento della concentrazione intracellulare di Ca2+ era abolito (p=0.01**)
(fig.34). Per escludere che la mancata risposta allo stimolo fosse dovuta ad una
desensitizzazione del recettore olfattivo abbiamo effettuato due tipi di controlli: i)
in un primo caso abbiamo stimolato i glomeruli con un puff odoroso, dopodiché si
è effettuato un lavaggio e si sono attesi circa 15 minuti prima di eseguire una
seconda stimolazione. Abbiamo verificato che la seconda risposta era
sostanzialmente simile alla prima per dinamiche temporali ed ampiezza del
segnale. ii) In un secondo controllo, abbiamo applicato ai glomeruli dapprima un
puff di odori in presenza di inibitore e, dopo il lavaggio ed un’attesa di 15 minuti,
abbiamo ripetuto la stimolazione in assenza di SQ22536. Anche in questo caso, la
risposta Ca2+ era visibile solamente senza il bloccante.
A causa della struttura molto compatta dei glomeruli all’interno del bulbo
olfattivo, un puff odoroso può raggiungere diversi glomeruli adiacenti. Tuttavia,
data la specificità molecolare del RO di ciascun glomerulo, circa il 30% dei
glomeruli stimolati mostrava un incremento della concentrazione di Ca2+. Al
contrario, in seguito al trattamento con forscolina, in tutti i glomeruli stimolati si
rilevava un aumento del segnale Ca2+.
82
a
c
b
e
d
Fig.34 : cinetiche dei cambiamenti nell’intesità di emissione (∆F/F0) nei terminali assonici di NSO
in preparati di emiteste in seguito all’applicazione di un puff odoroso sui glomeruli (a). In (b)
cinetica dei cambiamenti di fluorescenza dello stesso glomerulo sciacquato e stimolato di nuovo
con mix odorosa in presenza del bloccante dell’adenilato ciclasi SQ22536. In (c) e (d) immagini in
pseudocolori che mostrano lo stesso glomerulo di cui la cinetica è mostrata in (a), prima (a
sinistra) e dopo (a destra) applicazione dello stimolo odoroso (c) e dello stimolo odoroso in
presenza di bloccante SQ22536 (d). In (e) media della percentuale di risposta delle cellule allo
stimolo odoroso, calcolata prima (in nero) e dopo (in rosso) l’aggiunta del bloccante dell’adenilato
ciclasi (p=0.01**). Barre= s.e.m.
Questi esperimenti condotti sulla struttura olfattiva integra confermano i dati
esposti finora: il recettore olfattivo sul cono di crescita è funzionante, è in grado di
legare gli odori ed è associato ad un aumento locale di Ca2+ attraverso i canali
CNG. Bloccando la sintesi di cAMP, infatti, l’influsso di Ca2+ attraverso tali
canali è abolito.
3.8 Espressione di Gs.
I dati finora disponibili riguardo l’espressione di Gs nei NSO (vedi §1.14) sono
relativi alla vita embrionale ed ai primi giorni di vita post-natale (fino a PD5)
(16,17). Per capire se Gs continuasse a svolgere un ruolo nell’axon guidance
durante la vita adulta, abbiamo deciso di analizzare l’espressione di Gs non solo
nei neonati, ma anche in animali adulti. Si sono eseguiti saggi di
immunoistochimica per visualizzare la presenza e la distribuzione della proteina
Gs lungo l’assone dei NSO. Abbiamo analizzato l’espressione di Gs in stadi
precoci dello sviluppo post-natale (PD2-PD5) e in età adulta (quale PD20). Per
83
essere in grado di determinare se la marcatura che si osservava per Gs era
effettivamente localizzata sui neuroni sensoriali olfattivi è stata effettuata una
doppia marcatura con OMP. La marcatura con OMP, poiché il protocollo era
ottimizzato, è stata utilizzata come controllo sia dell’integrità delle fettine che
dell’effettiva riuscita del saggio.
Inizialmente i saggi sono stati eseguiti sul ratto. Come si può osservare dalla
fig.35 era presente una forte marcatura di Gs a livello dei glomeruli e nello strato
del nervo olfattivo, costituito dalle estremità distali degli assoni dei NSO. La
marcatura di Gs colocalizzava con la marcatura di OMP. Non si osservavano
sostanziali differenze nella distribuzione della marcatura di Gs in sezioni di bulbo
di ratti PD2-PD5 e di ratti PD20 (fig.35).
Fig.35: immunoistochimica per Gs in sezioni coronali di bulbo olfattivo di ratti PD2 (a) e PD20
(b). Nella colonna a sinistra l’immunofluorescenza di OMP, al centro di Gs e a destra il merged.
Lo stesso studio è stato effettuato sul topo e come si può osservare in fig.36 i
risultati sono sostanzialmente simili: si osservava una forte marcatura di Gs in
corrispondenza dei glomeruli, ma in parte anche nello strato del nervo olfattivo.
84
La distribuzione della marcatura osservata in topi di età PD2-PD5 era
paragonabile alla distribuzione ed ai livelli di marcatura osservati in animali
PD20.
Fig.36: immunoistochimica per Gs in sezioni coronali di bulbi olfattivi di topi PD2 (a) e PD20 (b).
Nella colonna a sinistra l’immunofluorescenza per OMP, al centro per Gs e a destra il merged.
Questi dati indicano che la proteina è presente sul terminale assonico non solo nei
primi giorni di vita post-natale, ma anche in età adulta. La presenza di Gs nei
glomeruli di animali adulti è consistente con la natura costantemente rigenerante
dell’epitelio olfattivo. Non possiamo escludere che Gs persista nei terminali
assonici anche oltre il periodo di allungamento assonale, svolgendo un ruolo nel
mantenimento del contatto sinaptico tra elementi pre- e post-sinaptici.
85
86
4. Discussione
I risultati ottenuti durante il mio dottorato dimostrano, per la prima volta, che il
recettore olfattivo localizzato sull’assone terminale-cono di crescita dei neuroni
sensoriali olfattivi è funzionante ed è capace di riconoscere e legare le molecole
odorose.
L’attivazione del RO sul cono di crescita è associata ad un aumento locale della
concentrazione intracellulare di cAMP. Analizzando le dinamiche di variazione di
cAMP nelle cilia-dendrite, soma ed assone terminale-cono di crescita di neuroni
sensoriali olfattivi in coltura, trasfettati con un sensore per il cAMP abbiamo
riscontrato che a seguito di stimoli sia farmacologici (forscolina, un generico
attivatore delle adenilato ciclasi, e IBMX, un inibitore aspecifico delle
fosfodiesterasi) sia fisiologici (odori) si osserva un aumento di cAMP in tutto il
neurone, incluso l’assone terminale-cono di crescita. Le dinamiche temporali di
questo incremento di cAMP sono le stesse nei NSO trattati con stimoli
farmacologici o fisiologici. Infatti, indipendentemente dal tipo di stimolo,
l’aumento di cAMP si verifica più velocemente a livello di dendrite distale ed
assone terminale rispetto al soma. Al momento non possiamo escludere che
l’incremento di cAMP a livello del corpo cellulare sia dovuto a diffusione dai
processi piuttosto che a sintesi locale.
I dati ottenuti trattando i neuroni con IBMX indicano la presenza di attività
costitutiva dell’adenilato ciclasi ed i livelli intracellulari di cAMP sono mantenuti
bassi dall’azione delle fosfodiesterasi.
Inoltre, abbiamo osservato che le differenze nelle dinamiche temporali registrate
nei vari compartimenti cellulari dei neuroni sensoriali olfattivi non sono ascrivibili
ad un ruolo delle fosfodiesterasi, dato che tali differenze permangono anche in
seguito all’inattivazione delle PDE in concomitanza con l’attivazione delle
adenilato ciclasi.
I dati ottenuti indicano quindi che le strutture molecolari necessarie per la sintesi e
la degradazione del cAMP sono presenti ed attive non solo nelle cilia, come noto,
ma anche a livello del terminale assonico-cono di crescita.
87
I nostri dati sono in accordo con la distribuzione delle adenilato ciclasi, come è
stato dimostrato da saggi di immunoistochimica (32). Il livello di questa proteina,
infatti, è elevato nel terminale dendritico e nella proiezione assonale, inclusi lo
strato del nervo olfattivo ed i glomeruli, mentre è pressocchè non rilevabile nel
soma.
Il dato più rilevante è che la produzione di cAMP a seguito di stimolazione
odorosa è dovuta all’attivazione del recettore olfattivo espresso a questo livello e
non è dovuta a diffusione di cAMP dalle cilia-dendrite o dal soma, né ad
attivazione Ca2+-dipendente dell’adenilato ciclasi presente nell’assone distale.
La produzione locale di cAMP è stata ulteriormente comprovata dagli esperimenti
in cui il recettore olfattivo al terminale assonico-cono di crescita viene stimolato
direttamente con un puff odoroso applicato tramite una micropipetta. In tali
condizioni sperimentali l’incremento di cAMP è presente esclusivamente
all’assone terminale e non si notano apprezzabili cambiamenti nel segnale negli
altri compartimenti del NSO.
Abbiamo inoltre dimostrato che l’attivazione da parte di stimoli fisiologici (odori)
del recettore olfattivo a livello del terminale assonico-cono di crescita è associata
ad un influsso di Ca2+ attraverso i canali CNG. Infatti l’incremento di Ca2+ è
abolito in presenza di bloccanti dei canali CNG (MgCl2) ed in presenza di inibitori
delle adenilato ciclasi (SQ22536).
Il confronto tra le cinetiche dell’incremento di cAMP e di Ca2+ mostra, tuttavia,
che questi due segnali presentano un pattern temporale molto diverso: i)
l’aumento di [Ca2+]] è rapido (con la nostra risoluzione temporale, 1
immagine/secondo, non si osserva un’apprezzabile fase di latenza tra l’aggiunta
della mix di odori e l’inizio del segnale Ca2+) e transiente (il picco è raggiunto in
circa 3-5 s e la [Ca2+] ritorna circa a livello basale in 30-60 s); ii) l’aumento di
cAMP, al contrario, è più lento e sostenuto, i.e. non tende a diminuire per almeno
5-10 min dopo l’applicazione degli odori. La spiegazione più semplice per queste
differenti cinetiche temporali è un’estrema sensibilità dei canali CNG per la
concentrazione locale di cAMP in stretta prossimità della membrana plasmatica.
In seguito al legame dell’odore al RO, si ha l’attivazione di una proteina G.
L’adenilato ciclasi che a sua volta viene attivata è localizzata in membrana, in
prossimità dei canali CNG. L’aumento di cAMP è altamente circoscritto alla
88
membrana plasmatica, in vicinanza dei canali CNG, cosicchè anche minimi
incrementi di cAMP possano raggiungere ed attivare tali canali. Una spiegazione
alternativa, i.e. una maggiore affinità per il cAMP dei canali CNG rispetto al
sensore basato su PKA, sembra da escludere, dal momento che l’affinità per il
cAMP della subunità regolatoria della PKA (e quindi del nostro sensore) è più
elevata di quella dei canali CNG (kM 0.2-0.3 µM vs 3 µM rispettivamente per
PKA e canali CNG) (42,95). La natura transiente del segnale Ca2+ e la durata
prolungata dell’incremento di cAMP suggeriscono che i canali CNG si inattivino
più velocemente (o in modo maggiore) rispetto al RO, per lo meno nelle attuali
condizioni sperimentali.
Esperimenti ex vivo, condotti su preparati di emiteste, escludono che i dati finora
esposti, ottenuti su NSO in coltura, siano influenzati dalla procedura di isolamento
e/o dalle condizioni di coltura. Caricando in vivo i NSO con un indicatore del Ca2+
fluorescente, abbiamo potuto dimostrare che l’applicazione locale a livello dei
glomeruli di forscolina o, più importante, di odori risulta in un rapido e transiente
incremento di [Ca2+] a livello dei terminali pre-sinaptici, con cinetiche molto
simili a quelle ottenute sui neuroni sensoriali olfattivi in coltura. Inoltre, come
atteso per un recettore olfattivo accoppiato ad attivazione di adenilato ciclasi,
l’incremento di Ca2+ in seguito a stimolazione odorosa è completamente abolito
dall’inibitore delle AC, SQ22536.
La domanda che sorge a questo punto è quale sia il ruolo funzionale degli
incrementi di cAMP e Ca2+ generati in seguito all’attivazione del recettore
olfattivo sull’assone terminale-cono di crescita. La funzione più ovvia, associabile
al RO sul cono di crescita, è che esso sia coinvolto nel processo di crescita
direzionata degli assoni verso il corretto target a livello del bulbo olfattivo. Questo
è un fenomeno molto complesso, che avviene in diverse tappe. Nel sistema
olfattivo si possono individuare almeno due eventi: la convergenza iniziale degli
assoni a formare glomeruli e la disposizione dei glomeruli in specifici siti
all’interno del BO. E’ stato proposto che il segnale cAMP derivante dal recettore
olfattivo giochi un ruolo chiave nella convergenza assonale, modulando
l’espressione di molecole guida. In questo modello la posizione del RO ed il sito
di produzione del cAMP possono essere irrilevanti. Tuttavia, topi knock out per
Golf sono anosmici, ma presentano una mappa sensoriale normale a livello del
89
bulbo olfattivo (17), portando quindi ad escludere che il cAMP generato alle cilia
sia coinvolto nella convergenza assonale e nel posizionamento dei glomeruli. Al
contrario, i nostri dati dimostrano che il RO all’assone terminale-cono di crescita
è funzionale e questo sembra in accordo con l’ipotesi che il recettore al terminale
assonale agisca come molecola guida, attraverso la produzione locale di cAMP e
Ca2+. Il ruolo di tali secondi messaggeri nell’axon guidance sul cono di crescita è
stato ampiamente studiato in altri sistemi ed è stato dimostrato che i livelli locali
di cAMP e Ca2+ possono modulare il comportamento del cono di crescita in
risposta a molecole guida esterne (67,72). In particolare, in Xenopus laevis si è
osservato che il cAMP prodotto localmente al cono di crescita determina
l’orientamento della crescita assonale in risposta a molecole segnale esterne
implicate nell’axon guidance. Diversi livelli di cAMP modificano il tipo di
risposta (da attrattiva a repulsiva o viceversa) alla stessa molecola guida.
Benchè i canali CNG non sembrino giocare un ruolo chiave nella formazione
della mappa sensoriale, i nostri dati indicano che tali canali sono associati al
recettore olfattivo sul terminale assonico-cono di crescita e possono avere un
ruolo modulatorio nella formazione della mappa sensoriale. E’ stato dimostrato
che livelli di nucleotidi ciclici all’assone terminale modulano la neurotrasmissione
attraverso i canali CNG e questo può avere un ruolo regolatorio nell’axon
guidance. Inoltre il Ca2+ che entra attraverso tali canali può interagire con il
cAMP e determinare infine la direzione di crescita dell’assone. La regolazione
reciproca tra questi due secondi messaggeri nel processo di axon guidance è stata
dimostrata in altri sistemi (67,72).
I dati ottenuti suggeriscono quindi che il cAMP prodotto a seguito dell’attivazione
del RO sul cono di crescita abbia la potenzialità di giocare un ruolo chiave nella
convergenza assonale. Questo può spiegare i risultati di esperimenti genetici che
talora potevano apparire contraddittori. Negli animali mancanti di CNG, le
risposte evocate dagli odori sono drasticamente ridotte, gli animali sono infatti
anosmici, ma la mappa glomerulare a livello del bulbo olfattivo è pressocchè
inalterata (81). Una situazione analoga, come già detto, si verifica per gli animali
knock out per Golf (17). Nei topi in cui l’espressione dell’adenilato ciclasi 3 è stata
geneticamente abolita, non si registra alcuna risposta agli odori e, più importante,
la mappa sensoriale è drammaticamente modificata, con mancata formazione della
90
maggior parte dei glomeruli ed alterata struttura di quelli presenti (82). I risultati
ottenuti nei knock out di Golf potrebbero non essere chiari, se non venissero
analizzati alla luce del fatto che nei neuroni sensoriali olfattivi le proteine G
stimolatorie sono due: Gs e Golf. E’ stato dimostrato che Golf è associata al
recettore olfattivo durante il processo di trasduzione del segnale odoroso (21),
mentre è stato solo ipotizzato che Gs sia associata al RO durante il processo di
crescita assonale. A sostegno di tale ipotesi, abbiamo osservato che Gs è espressa
non solo nelle prime fasi di sviluppo post-natale (PD2-PD5), come già dimostrato,
ma anche in età adulta (>PD20), nello strato del nervo olfattivo e nello strato
glomerulare, in accordo con la natura continuamente rigenerante dell’epitelio
olfattivo (16). Non possiamo tuttavia escludere un ruolo di Gs nel mantenimento
dei terminali pre-sinaptici a livello dei glomeruli. L’elemento che emerge da
questi esperimenti è che Golf ed i canali CNG sono indispensabili per la
trasduzione del segnale odoroso, mentre Gs e cAMP possono giocare un ruolo
chiave nella formazione della mappa sensoriale (84).
Abbiamo inoltre dimostrato che la selettiva attivazione del RO al cono di crescita
è seguita dalla traslocazione nucleare della subunità catalitica della sonda PKA.
Quindi il cAMP prodotto dopo l’attivazione del RO al terminale assonico-cono di
crescita agisce non solo localmente, ma anche a livello nucleare, dove potrebbe
modulare l’espressione di molecole coinvolte nell’axon guidance. Mentre l’azione
locale del CAMP è rapida e può determinare il comportamento del cono di
crescita in risposta a molecole locali, l’espressione genica dipendente da PKA è
un processo a lungo termine, che potrebbe intervenire dopo un intervallo di tempo
maggiore, e.g. per determinare la posizione dei glomeruli nel bulbo olfattivo.
Sono state proposte altre ipotesi per spiegare il ruolo funzionale del recettore
olfattivo espresso al cono di crescita. Ad esempio è stato suggerito che la
convergenza assonale dipenda da interazioni omofiliche tra i RO (76,86). Anche
se nel nostro progetto sperimentale non ci siamo soffermati su questo punto, la
dimostrazione che i recettori olfattivi all’assone terminale sono funzionali ed
accoppiati all’attivazione di adenilato ciclasi e canali CNG appare maggiormente
in accordo con l’idea che gli assoni di neuroni esprimenti lo stesso RO siano
guidati da molecole segnale che attivano localmente i loro RO sul cono di
crescita.
91
La domanda che rimane aperta è quale siano i ligandi naturali in grado di attivare i
RO sul cono di crescita. I nostri risultati dimostrano che gli odori legano ed
attivano il recettore olfattivo sul terminale assonale di neuroni in coltura e, più
importante, in situ. Questo suggerisce che gli odori potrebbero rappresentare i
ligandi naturali del RO anche sul cono di crescita. Essi infatti rimangono al
momento le sole molecole conosciute in grado di attivare in modo specifico e
selettivo
più
di
mille
recettori
olfattivi,
strutturalmente
identici
ma
molecolarmente distinti. Per quanto riguarda la modalità con cui gli odori possono
raggiungere i recettori olfattivi sul cono di crescita, si possono immaginare diversi
scenari: attraverso vescicole, mediante trasporto lungo le membrane ed anche
attraverso il flusso ematico. Al momento non possiamo certamente escludere altri
possibili meccanismi di attivazione del RO all’assone terminale, come ad esempio
un’attivazione costitutiva del recettore stesso o l’esistenza di molecole guida nel
bulbo olfattivo in grado di legare ed attivare i RO.
Qualunque possa essere il meccanismo di attivazione del recettore olfattivo sul
terminale assonico, i nostri dati sui meccanismi molecolari sottesi al RO espresso
al cono di crescita costituiscono un elemento fondamentale ed imprescindibile per
capire la funzione del recettore olfattivo sull’assone terminale.
92
Bibliografia
1) Ronnett GV, Moon C. G proteins and olfactory signal transduction.
Annu.Rev.Physiol. 2002;64:189-222.
(2) Lledo PM, Gheusi G, Vincent JD. Information processing in the
mammalian olfactory system. Physiol.Rev. 2005 Jan;85(1):281-317.
(3) Menini A. Calcium signalling and regulation in olfactory neurons.
Curr.Opin.Neurobiol. 1999 Aug;9(4):419-426.
(4) Firestein S. How the olfactory system makes sense of scents. Nature 2001
Sep 13;413(6852):211-218.
(5) Malnic B, Hirono J, Sato T, Buck LB. Combinatorial receptor codes for
odors. Cell 1999 Mar 5;96(5):713-723.
(6) Buck L, Axel R. A novel multigene family may encode odorant receptors:
a molecular basis for odor recognition. Cell 1991 Apr 5;65(1):175-187.
(7) Godfrey PA, Malnic B, Buck LB. The mouse olfactory receptor gene
family. Proc.Natl.Acad.Sci.U.S.A. 2004 Feb 17;101(7):2156-2161.
(8) Zhang X, Firestein S. The olfactory receptor gene superfamily of the
mouse. Nat.Neurosci. 2002 Feb;5(2):124-133.
(9) Malnic B, Godfrey PA, Buck LB. The human olfactory receptor gene
family. Proc.Natl.Acad.Sci.U.S.A. 2004 Feb 24;101(8):2584-2589.
(10) Young JM, Trask BJ. The sense of smell: genomics of vertebrate odorant
receptors. Hum.Mol.Genet. 2002 May 15;11(10):1153-1160.
93
(11) Monti-Graziadei
GA, Margolis FL,
Immunocytochemistry
of
the
Harding JW, Graziadei PP.
olfactory
marker
protein.
J.Histochem.Cytochem. 1977 Dec;25(12):1311-1316.
(12) Monti-Graziadei GA, Stanley RS, Graziadei PPC The olfactory marker
protein in the olfactory system of the mouse during development.
Neurosci.1980 Jul;5(7):1239-1252.
(13) Nef P, Hermans-Borgmeyer I, Artieres-Pin H, Beasley L, Dionne VE,
Heinemann SF. Spatial pattern of receptor expression in the olfactory
epithelium. Proc.Natl.Acad.Sci.U.S.A. 1992 Oct 1;89(19):8948-8952.
(14) Ressler KJ, Sullivan SL, Buck LB. A zonal organization of odorant
receptor gene expression in the olfactory epithelium. Cell 1993 May
7;73(3):597-609.
(15) Chess A, Simon I, Cedar H, Axel R. Allelic inactivation regulates olfactory
receptor gene expression. Cell 1994 Sep 9;78(5):823-834.
(16) Jones DT, Masters SB, Bourne HR, Reed RR. Biochemical characterization
of three stimulatory GTP-binding proteins. The large and small forms of Gs
and
the
olfactory-specific
G-protein,
Golf.
J.Biol.Chem.
1990
Feb
15;265(5):2671-2676.
(17) Belluscio L, Gold GH, Nemes A, Axel R. Mice deficient in G(olf) are
anosmic. Neuron 1998 Jan;20(1):69-81.
(18) Pace U, Hanski E, Salomon Y, Lancet D. Odorant-sensitive adenylate
cyclase
may
mediate
olfactory
reception.
Nature
1985
Jul
18-
24;316(6025):255-258.
(19) Sklar PB, Anholt RR, Snyder SH. The odorant-sensitive adenylate cyclase
of olfactory receptor cells. Differential stimulation by distinct classes of
odorants. J.Biol.Chem. 1986 Nov 25;261(33):15538-15543.
94
(20) Nakamura T, Gold GH. A cyclic nucleotide-gated conductance in olfactory
receptor cilia. Nature 1987 Jan 29-Feb 4;325(6103):442-444.
(21) Jones DT, Reed RR. Golf: an olfactory neuron specific-G protein involved
in odorant signal transduction. Science 1989 May 19;244(4906):790-795.
(22) Bakalyar HA, Reed RR. Identification of a specialized adenylyl cyclase
that may mediate odorant detection. Science 1990 Dec 7;250(4986):1403-1406.
(23) Kleene SJ. Origin of the chloride current in olfactory transduction.
Neuron 1993 Jul;11(1):123-132.
(24) Lowe G, Gold GH. Nonlinear amplification by calcium-dependent
chloride
channels
in
olfactory
receptor
cells.
Nature
1993
Nov
18;366(6452):283-286.
(25) Liu M, Chen TY, Ahamed B, Li J, Yau KW. Calcium-calmodulin
modulation of the olfactory cyclic nucleotide-gated cation channel. Science
1994 Nov 25;266(5189):1348-1354.
(26) Pifferi S, Boccaccio A, Menini A. Cyclic nucleotide-gated ion channels in
sensory transduction. FEBS Lett. 2006 May 22;580(12):2853-2859.
(27) Sadana R, Dessauer CW. Physiological Roles for G Protein-Regulated
Adenylyl Cyclase Isoforms: Insights from Knockout and Overexpression
Studies. Neurosignals 2008 Oct 24;17(1):5-22.
(28) Chen Y, Cann MJ, Litvin TN, Iourgenko V, Sinclair ML, Levin LR, et al.
Soluble adenylyl cyclase as an evolutionarily conserved bicarbonate sensor.
Science 2000 Jul 28;289(5479):625-628.
(29) Litvin TN, Kamenetsky M, Zarifyan A, Buck J, Levin LR. Kinetic
properties of "soluble" adenylyl cyclase. Synergism between calcium and
bicarbonate. J.Biol.Chem. 2003 May 2;278(18):15922-15926.
95
(30) Bakalyar HA, Reed RR. The second messenger cascade in olfactory
receptor neurons. Curr.Opin.Neurobiol. 1991 Aug;1(2):204-208.
(31) Wong ST, Trinh K, Hacker B, Chan GC, Lowe G, Gaggar A, et al.
Disruption of the type III adenylyl cyclase gene leads to peripheral and
behavioral anosmia in transgenic mice. Neuron 2000 Sep;27(3):487-497.
(32) Col JA, Matsuo T, Storm DR, Rodriguez I. Adenylyl cyclase-dependent
axonal targeting in the olfactory system. Development 2007 Jul;134(13):24812489.
(33) Beavo JA. Cyclic nucleotide phosphodiesterases: functional implications
of multiple isoforms. Physiol.Rev. 1995 Oct;75(4):725-748.
(34) Lugnier C. Cyclic nucleotide phosphodiesterase (PDE) superfamily: a
new
target
for
the
development
of
specific
therapeutic
agents.
Pharmacol.Ther. 2006 Mar;109(3):366-398.
(35) Bender AT, Beavo JA. Cyclic nucleotide phosphodiesterases: molecular
regulation to clinical use. Pharmacol.Rev. 2006 Sep;58(3):488-520.
(36) Moon C, Simpson PJ, Tu Y, Cho H, Ronnett GV. Regulation of
intracellular cyclic GMP levels in olfactory sensory neurons. J.Neurochem.
2005 Oct;95(1):200-209.
(37) Borisy FF, Ronnett GV, Cunningham AM, Juilfs D, Beavo J, Snyder SH.
Calcium/calmodulin-activated phosphodiesterase expressed in olfactory
receptor neurons. J.Neurosci. 1992 Mar;12(3):915-923.
(38) Sette C, Iona S, Conti M. The short term activation of a rolipramsensitive, cAMP-specific phosphodiesterase by thyroid-stimulating hormone
in thyroid FRTL-5 cells is mediated by a cAMP-dependent phosphorilation.
J.Biol.Chem. 1994 Mar;269(12):9245-52.
96
(39) Fesenko EE, Kolesnikov SS, Lyubarsky AL. Induction by cyclic GMP of
cationic conductance in plasma membrane of retinal rod outer segment.
Nature 1985 Jan 24-30;313(6000):310-313.
(40) Bradley J, Reisert J, Frings S. Regulation of cyclic nucleotide-gated
channels. Curr.Opin.Neurobiol. 2005 Jun;15(3):343-349.
(41) Zagotta WN, Siegelbaum SA. Structure and function of cyclic nucleotidegated channels. Annu.Rev.Neurosci. 1996;19:235-263.
(42) Nakamura T. Cellular and molecular constituents of olfactory sensation
in
vertebrates.
Comp.Biochem.Physiol.A.Mol.Integr.Physiol.
2000
May;126(1):17-32.
(43) Gordon SE, Brautigan DL, Zimmerman AL. Protein phosphatases
modulate the apparent agonist affinity of the light-regulated ion channel in
retinal rods. Neuron 1992 Oct;9(4):739-748.
(44) Gordon SE, Zagotta WN. Subunit interactions in coordination of Ni2+ in
cyclic
nucleotide-gated
channels.
Proc.Natl.Acad.Sci.U.S.A.
1995
Oct
24;92(22):10222-10226.
(45) Shepherd G. The synaptic organization of the brain. Oxford: Oxford
University Press 1998.
(46) Shipley MT, Ennis M. Functional organization of olfactory system.
J.Neurobiol. 1996 May;30(1):123-176.
(47) Mori K, Nagao H, Yoshihara Y. The olfactory bulb: coding and processing
of odor molecule information. Science 1999 Oct 22;286(5440):711-715.
(48) Bozza TC, Kauer JS. Odorant response properties of convergent olfactory
receptor neurons. J.Neurosci. 1998 Jun 15;18(12):4560-4569.
(49) Ressler KJ, Sullivan SL, Buck LB. Information coding in the olfactory
system: evidence for a stereotyped and highly organized epitope map in the
olfactory bulb. Cell 1994 Dec 30;79(7):1245-1255.
97
(50) Vassar R, Chao SK, Sitcheran R, Nunez JM, Vosshall LB, Axel R.
Topographic organization of sensory projections to the olfactory bulb. Cell
1994 Dec 16;79(6):981-991.
(51) Belluscio L, Katz LC. Symmetry, stereotypy, and topography of odorant
representations in mouse olfactory bulbs. J.Neurosci. 2001 Mar 15;21(6):21132122.
(52) Wachowiak M, Cohen LB. Representation of odorants by receptor neuron
input to the mouse olfactory bulb. Neuron 2001 Nov 20;32(4):723-735.
(53) Mombaerts P, Wang F, Dulac C, Chao SK, Nemes A, Mendelsohn M, et al.
Visualizing an olfactory sensory map. Cell 1996 Nov 15;87(4):675-686.
(54) Potter SM, Zheng C, Koos DS, Feinstein P, Fraser SE, Mombaerts P.
Structure and emergence of specific olfactory glomeruli in the mouse.
J.Neurosci. 2001 Dec 15;21(24):9713-9723.
(55) Treloar HB, Feinstein P, Mombaerts P, Greer CA. Specificity of glomerular
targeting by olfactory sensory axons. J.Neurosci. 2002 Apr 1;22(7):2469-2477.
(56) John JA, Key B. Axon mis-targeting in the olfactory bulb during
regeneration of olfactory neuroepithelium. Chem.Senses 2003 Nov;28(9):773779.
(57) Strotmann J, Levai O, Fleischer J, Schwarzenbacher K, Breer H. Olfactory
receptor proteins in axonal processes of chemosensory neurons. J.Neurosci.
2004 Sep 1;24(35):7754-7756.
(58) Royal SJ, Key B. Development of P2 olfactory glomeruli in P2-internal
ribosome entry site-tau-LacZ transgenic mice. J.Neurosci. 1999 Nov
15;19(22):9856-9864.
(59) Conzelmann S, Malun D, Breer H, Strotmann J. Brain targeting and
glomerulus formation of two olfactory neuron populations expressing related
receptor types. Eur.J.Neurosci. 2001 Nov;14(10):1623-1632.
98
(60) Zou DJ, Feinstein P, Rivers AL, Mathews GA, Kim A, Greer CA, et al.
Postnatal refinement of peripheral olfactory projections. Science 2004 Jun
25;304(5679):1976-1979.
(61) Strotmann J, Breer H. Formation of glomerular maps in the olfactory
system. Semin.Cell Dev.Biol. 2006 Aug;17(4):402-410.
(62) Nakatani H, Serizawa S, Nakajima M, Imai T, Sakano H. Developmental
elimination of ectopic projection sites for the transgenic OR gene that has lost
zone
specificity
in
the
olfactory
epithelium.
Eur.J.Neurosci.
2003
Nov;18(9):2425-2432.
(63) Lodovichi C, Belluscio L, Katz LC. Functional topography of connections
linking mirror-symmetric maps in the mouse olfactory bulb. Neuron 2003 Apr
24;38(2):265-276.
(64) Yan Z, Tan J, Qin C, Lu Y, Ding C, Luo M. Precise circuitry links
bilaterally symmetric olfactory maps. Neuron 2008 May 22;58(4):613-624.
(65) Huber AB, Kolodkin AL, Ginty DD, Cloutier JF. Signaling at the growth
cone: ligand-receptor complexes and the control of axon growth and
guidance. Annu.Rev.Neurosci. 2003;26:509-563.
(66) Song HJ, Ming GL, Poo MM. cAMP-induced switching in turning
direction of nerve growth cones. Nature 1997 Jul 17;388(6639):275-279.
(67) Song HJ, Poo MM. Signal transduction underlying growth cone guidance
by diffusible factors. Curr.Opin.Neurobiol. 1999 Jun;9(3):355-363.
(68) Cutforth T, Moring L, Mendelsohn M, Nemes A, Shah NM, Kim MM, et al.
Axonal ephrin-As and odorant receptors: coordinate determination of the
olfactory sensory map. Cell 2003 Aug 8;114(3):311-322.
(69) Ming GL, Song HJ, Berninger B, Holt CE, Tessier-Lavigne M, Poo MM.
cAMP-dependent growth
cone guidance by
Dec;19(6):1225-1235.
99
netrin-1.
Neuron
1997
(70) Williams-Hogarth LC, Puche AC, Torrey C, Cai X, Song I, Kolodkin AL, et
al. Expression of semaphorins in developing and regenerating olfactory
epithelium. J.Comp.Neurol. 2000 Aug 7;423(4):565-578.
(71) Dontchev VD, Letourneau PC. Nerve growth factor and semaphorin 3A
signaling pathways interact in regulating sensory neuronal growth cone
motility. J.Neurosci. 2002 Aug 1;22(15):6659-6669.
(72) Gomez TM, Zheng JQ. The molecular basis for calcium-dependent axon
pathfinding. Nat.Rev.Neurosci. 2006 Feb;7(2):115-125.
(73) Wang F, Nemes A, Mendelsohn M, Axel R. Odorant receptors govern the
formation of a precise topographic map. Cell 1998 Apr 3;93(1):47-60.
(74) Serizawa S, Miyamichi K, Nakatani H, Suzuki M, Saito M, Yoshihara Y, et
al. Negative feedback regulation ensures the one receptor-one olfactory
neuron rule in mouse. Science 2003 Dec 19;302(5653):2088-2094.
(75) Feinstein P, Mombaerts P. A contextual model for axonal sorting into
glomeruli in the mouse olfactory system. Cell 2004 Jun 11;117(6):817-831.
(76) Serizawa S, Miyamichi K, Takeuchi H, Yamagishi Y, Suzuki M, Sakano H.
A neuronal identity code for the odorant receptor-specific and activitydependent axon sorting. Cell 2006 Dec 1;127(5):1057-1069.
(77) Taniguchi M, Nagao H, Takahashi YK, Yamaguchi M, Mitsui S, Yagi T, et
al. Distorted odor maps in the olfactory bulb of semaphorin 3A-deficient
mice. J.Neurosci. 2003 Feb 15;23(4):1390-1397.
(78) Barnea G, O'Donnell S, Mancia F, Sun X, Nemes A, Mendelsohn M, et al.
Odorant receptors on axon termini in the brain. Science 2004 Jun
4;304(5676):1468.
(79) Lin DM, Wang F, Lowe G, Gold GH, Axel R, Ngai J, et al. Formation of
precise connections in the olfactory bulb occurs in the absence of odorantevoked neuronal activity. Neuron 2000 Apr;26(1):69-80.
100
(80) Zhao H, Reed RR. X inactivation of the OCNC1 channel gene reveals a
role for activity-dependent competition in the olfactory system. Cell 2001 Mar
9;104(5):651-660.
(81) Zheng C, Feinstein P, Bozza T, Rodriguez I, Mombaerts P. Peripheral
olfactory projections are differentially affected in mice deficient in a cyclic
nucleotide-gated channel subunit. Neuron 2000 Apr;26(1):81-91.
(82) Zou DJ, Chesler AT, Le Pichon CE, Kuznetsov A, Pei X, Hwang EL,
Firestein S. Absence of adenylyl cyclase 3 perturbs peripheral projections in
mice. J.Neurosci. 2007 Jun;27(25):6675-83.
(83) Yu CR, Power J, Barnea G, O'Donnell S, Brown HE, Osborne J, et al.
Spontaneous neural activity is required for the establishment and
maintenance of the olfactory sensory map. Neuron 2004 May 27;42(4):553566.
(84) Imai T, Suzuki M, Sakano H. Odorant receptor-derived cAMP signals
direct axonal targeting. Science 2006 Oct 27;314(5799):657-661.
(85) Chesler AT, Zou DJ, Le Pichon CE, Peterlin ZA, Matthews GA, Pei X, et al.
A G protein/cAMP signal cascade is required for axonal convergence into
olfactory glomeruli. Proc.Natl.Acad.Sci.U.S.A. 2007 Jan 16;104(3):1039-1044.
(86) Feinstein P, Bozza T, Rodriguez I, Vassalli A, Mombaerts P. Axon guidance
of mouse olfactory sensory neurons by odorant receptors and the beta2
adrenergic receptor. Cell 2004 Jun 11;117(6):833-846.
(87) Zaccolo M, De Giorgi F, Cho CY, Feng L, Knapp T, Negulescu PA, et al. A
genetically encoded, fluorescent indicator for cyclic AMP in living cells.
Nat.Cell Biol. 2000 Jan;2(1):25-29.
(88) Miyawaki A, Tsien RY. Monitoring protein conformations and
interactions by fluorescence resonance energy transfer between mutants of
green fluorescent protein. Methods Enzymol. 2000;327:472-500.
101
(89) Zaccolo M, Pozzan T. Discrete microdomains with high concentration of
cAMP in stimulated rat neonatal cardiac myocytes. Science 2002 Mar
1;295(5560):1711-1715.
(90) Lissandron V, Terrin A, Collini M, D'alfonso L, Chirico G, Pantano S, et al.
Improvement of a FRET-based indicator for cAMP by linker design and
stabilization of donor-acceptor interaction. J.Mol.Biol. 2005 Dec 2;354(3):546555.
(91) Ponsioen B, Zhao J, Riedl J, Zwartkruis F, van der Krogt G, Zaccolo M, et al.
Detecting cAMP-induced Epac activation by fluorescence resonance energy
transfer: Epac as a novel cAMP indicator. EMBO Rep. 2004 Dec;5(12):11761180.
(92) Ronnett GV, Hester LD, Snyder SH. Primary culture of neonatal rat
olfactory neurons. J.Neurosci. 1991 May;11(5):1243-1255.
(93) Slemmer JE, Matsushita S, De Zeew CL, Weber JT, Knopfel T. Glutamateinduced elevations in intracellular chloride concentration in hippocampal cell
cultures
derived
from
EYFP-expressing
mice.
Eur.J.Neurosci.
2004
Jun;19(11):2915-2922.
(94) Lee HM, Park YS, Kim W, Park CS. Elettrophysiological characteristic of
rat gustatory cyclic nucleotide-gated channels expressed in Xenopus oocytes.
J.Neurophysiol. 2001 Jun;85(6):2335-2348.
(95) Beavo JA, Betchel PJ, Krebs EG. Activation of protein kinase by
physiological concentrations of cyclic AMP. Proc.Natl.Acad.Sci. U S A 1974
Sep;71(9):3580-3583.
102
Ringraziamenti
Desidero innanzitutto ringraziare la Dott.ssa Claudia Lodovichi per questo
progetto e per il costante aiuto e l'incoraggiamento nei momenti di difficoltà di
questi tre anni di dottorato.
Ringrazio la Dott.ssa Manuela Zaccolo e tutti ragazzi del suo laboratorio per
averci gentilmente fornito i sensori per il cAMP e per i preziosi suggerimenti.
Grazie al Professor Tullio Pozzan, per l'interessamento che ha dimostrato per
questo progetto di ricerca e per i suoi consigli.
Uno speciale ringraziamento ai miei colleghi Giovanni Monaco, per la parte di
Ca2+ imaging ex vivo, e Raffaella Flaibani per gli esperimenti su Gs. Il loro
contributo a questo lavoro è stato fondamentale!
Un grazie enorme alla mia collega e compagna Mara Pietrobon, per l'importante
cooperazione con cui abbiamo sempre lavorato.
Un grazie meno formale a tutti i miei compagni di laboratorio, che mi hanno
permesso di svolgere questo dottorato in un clima di amicizia. Grazie Giovanni
per la tua simpatia che ci ha aiutato a sdramatizzare e a sorridere anche quando
non ne avevamo tanta voglia! Grazie Mara per la tua amicizia, la tua
organizzazione e la straordinaria intesa (telepatia?)! Grazie Raffaella per le
lunghe chiacchierate, l'aiuto e la disponibilità, la passione per Liga... Grazie
Federica per l'amicizia, i viaggi, le cene etniche. Grazie a Giulia per essermi
sempre stata vicina, soprattutto all'inizio. Insomma grazie a tutti, perchè ho avuto
l'opportunità di conoscere delle persone veramente speciali.
Infine un ringraziamento speciale alla mia famiglia, in particolare i miei genitori,
che mi hanno sempre sostenuto nelle mie scelte e mi hanno incoraggiato nei
momenti difficili, spronandomi ad andare avanti. Senza di loro non sarei arrivata
al traguardo!
103
104