ONCOGENI ed ONCOSOPPRESSORI
Nabissi2016
Iprodo0deglioncogenipossonoessereclassifica7inseigruppi:
1. Fa<oriditrascrizione
2. Rimodellatoridellacroma7na
3. Fa<oridicrescita
4. Rece<orideifa<oridicrescita
5. Trasdu<ridelsegnale
6. Regolatoridell’apoptosi
Nabissi2016
FATTORIDITRASCRIZIONE
Ifa<oriditrascrizione(TF)sonospessomembri
diunafamigliamul7genicachemostradei
doministru<uralicomuniemol7diques7fa<ori
perfunzionarerichiedonol’interazioneconaltre
proteine.IlTFregolanoposi7vamenteo
nega7vamentel’espressionedidetermina7geni,
legandosiaspecifichesequenzediDNA
(promoter,enhancer,silencer).EsempidiTF,
codifica7anchedaretrovirussonoc-myc,fosejun.
Nabissi2016
• c-myc
Ilgenemycèconservatointu<elespecieanimaliefu
inizialmentescopertoindiversiretrovirusresponsabili
nell’induzionedisarcomi,carcinomieleucemiemieloidi.
Laproteinacodificatadamycèunfa<oretrascrizionale
(p64myc)presentenellecelluleina0vaproliferazione
anchesemycsvolgeunruoloimportanteanche
nell’indurrel’apoptosi.Mycèclassificatotraigenedella
prontarispostainsiemeafosejun,indo0dafa<oridi
crescitamitogenici,conilruolodiindurrelaprogressione
dallafaseG0-G1allafaseSdelciclocellulare.
Nabissi2016
Nabissi2016
• Myceciclocellulare:
• ciclineD,EeA
• genichecodificanoperleCDK2eCDK4
• genedell’enzimaorni7na-decarbossilasi(ODC),essenzialenel
metabolismodegliacidinucleici
• reprimeigenidelleCDKI(inibitoridelleCDK)
• a0vaigenichecodificanoperE2F
• reprimeilgenedell’inibitoredelleCDK(p27KIP1).
Nabissi2016
Nabissi2016
ILIVELLIECCESSIVIDIMYCPOSSONOINDURRELACELLULA
ADATTIVAREILPROCESSODIAPOPTOSI.
fosejun
Ilfa<oreditrascrizionefos(p55)dimerizzaconilfa<oredi
trascrizionejun(p39)performareilfa<oredi
trascrizioneAP-1,ilqualeaumental’espressionedi
diversigenichecontrollanoladivisionecellulare.
L’a0vitàdiAP-1èindoGadafaGoridicrescita,
citochine,ormonipolipe7dici,neurotrasme0tori,
interazionecellula-matrice,infezioniviralieba<erichee
dadiversistressfisiciechimici.
Nabissi2016
Nabissi2016
AP-1 è un dimero composto da proteine della famiglia Jun (c-jun, JunB,
JunD),
Fos (c-Fos, FosB, Fra-1, Fra-2).
Riconosce sequenze del DNA TGAG/CTCA o CRE (TGACGTCA)
Nabissi2016
RECETTORIPERIFATTORIDICRESCITA
L’eccessiva s7molazione della proliferazione cellulare, nei tumori, dipende anche
dall’amplificazione o dalle mutazioni dei geni codifican7 per i rece<ori dei fa<ori di
crescita,conconseguenteaumentodell’espressionedeireceGoriolasintesidireceGori
struGuralmenteefunzionalmentemodificaL,conconseguentealterazionedelsegnalee
guadagnodifunzione.
Tali rece<ori possono essere suddivisi in base ai residui presen7 nella regione
citoplasma7cain:Lrosin-chinasicieserin/treonin-chinasici
Nell’ambitodeitumori,irece<ori7rosinchinasici(RTK)sonoquellimaggiormentemutaL
neitumoriesonotraigenicodifica7daglioncogenivirali.
Nabissi2016
IRTKsipossonosuddividerein:
7rosinchinasicitoplasma7che
rece<oricona0vità7rosinchinasica
IRTKsonosuddivisiindiversefamigliechesicara<erizzanoprincipalmenteperlaporzione
aminoacidicaextracellularein:
porzionericcadicisteinecomeEGFR,IR,IGFR
dominisimiliaquellidelleIg(Iglikemo7f),comePDGFReFGFR
dominiconsequenzedifibronecLnacombinateasequenzeIg-like,comeefrinaAR,
efrinaBR,angiotensina.1R
Nabissi2016
2+
dominichereagisconoconlecaderineinpresenzadiCa
ARvazionedeireceGori
RTK: Il sito catali7co, in posizione C-terminale ha a0vità proteinchinasica, ed è
cos7tuitodatreregioni:1)leganteATP,2)a0vitàfosfotransferasica,
3) contenente una proteinchinasi a0vata da fosforilazione dopo legame del
ligando al rece<ore. Gli RTK sono presen7 in stato ina0vo come monomeri e
dimerizzano dopo interazione con il ligando. La dimerizzazione induce un
cambiamento conformazionale che a0va il sito catali7co con a0vità
7rosinchinasica,cheinducefosforilazioneneisi7catali7cicheinaltriresidui.
Nabissi2016
Ilprimosegnale
dell’aRvazionedelreceGore,
mediatadalligando,e’la
fosforilazioneneldominio
citoplasmaLcodelreceGore
stesso,mediatadalla
dimerizzazionedelreceGore.
receptor-directedprotein-tyrosinephosphatases(PTPs)
Nabissi2016
Illegameligando-receGorereclutaproteineadaGatrici
contenenLSH2domain
L’autofosforilazione dei receGori genera siL
di fosforilazione i quali diventano siL
receRvi per proteine SH2 che includono
fosfolipasi C o GRB (growth factor receptor
bound). I domini SH3 agiscono poi come
domini di legame per domini ricchi in
prolina presenL in molecole effeGrici che
agisconocomepercorsimolecolariavalle.
Nabissi2016
Trasduzionedelsegnale
La posizione specifica delle 7rosine presen7 nei RTK è quella che rende che
condizionaleinterazioniconspecificheproteinecitoplasma7chedafosforilare.
Inpar7colareidominiSH2presen7suspecificheproteine(proteinecona0vità
enzima7ca) vengono fosforilate dai TRK o mediante proteine ada<atrici che
facilitano il conta<o con le proteine che sono substrato delle proteinchinasi. I
residuicarichiposi7vamente,presen7nelleregioniSH2reagisconoconigruppi
fosfato(carichinega7vamente)presen7sulleTKeperme<onolaformazionedi
un complesso che a0va la proteina contenente il dominio SH2. Altri domini
proteici che interagiscono con le 7rosine fosforilate sono i SH3 e PTB
(phosphotyrosinbinding).
Nabissi2016
FATTORIDICRESCITA
I fa<ori di crescita (GF) sono proteine di piccole dimensioni che s7molano la
proliferazione, il differenziamento, la sopravvivenza e la mo7lità cellulare. La
maggiorpartedeiGFhannoeffe<oposi7vosuques7fenomeni,anchesealcuni
come il TGF-β (trasforming growth factor beta) agiscono come segnale
inibitorio.Nell’ambitodellepatologietumoralimol7genichecodificaperGF,si
comportano da oncogeni, in quanto subiscono mutazioni tali da indurre la
cellula ad una sovra-produzione del GF con conseguente a0vazione di uno
s7moloprolifera7voautocrinooparacrino.
Nabissi2016
WNT SIGNALING
Attivazione della trascrizione di geni bersaglio che inducono proliferazione
cellulare e disassociazione delle E-caderine con conseguente perdita dei legami
cellula/cellula e stimolo della mobilità cellulare.
Nabissi2016
InsulinGrowthFactor(IGF)
Fraivarifa<oridicrescitachegiocanounruolodeterminanteneivariprocessiche
inducono proliferazione cellulare, sopravvivenza cellulare e resistenza ai
chemioterapicieradioterapiac’èilsistemaIGF(insulingrowthfactor).Ilsistema
IGFconsistedidueligandi,IGF-IeIGF-II,trerece<oridimembrana(IGF-IR,IGFIIR)ereceGoredell’insulina(IR)eseiproteinedilegameadaltaaffinità(IGFBP
1-6). I rece<ori IGF-IR una volta trado0 vengono glicosila7, dimerizzano e
processa7induesubunitàαeβchea<raversopon7disolfurovannoaformareil
tetramerofunzionante(β-α-α-β)chevienetrasportatoinmembrana.A<raverso
la subunità α che è extracellulare si ha il legame del IGF, con conseguente
trasduzionedelsegnalealivellointracellularecheconsistenellafosforilazionedi
residuidi7rosinaindominidiLrosinchinasi.
Nabissi2016
Lacascatadia0vazionecomporta,principalmente,tre7pisisegnali:
1. a0vazionedelpathwaysdelfosfaLdilinositolo3-chinasi(PI-3K),Akt,mTORche
a0valasintesiproteicaeimeccanismian7apopto7cimedianteinaRvazione
(fosforilazione)diBad
2. a0vazionedelpathwaydiRas/Raf1/MEK/ERKcheinduceproliferazione
cellulare
3. a0vazionedialtrimeccanismicellularicheinduconol’espressionedifaGoridi
crescita,citochineeintegrine
Nabissi2016
Inoltre il IGF-IR svolge un ruolo nel facilitare :
1. il passaggio dalle fase G0 a G1 mediante attivazione della p70 S6K,
che fosforila la proteina ribosomiale S6 che stimola l’insieme di proteine
ribosomiali necessarie a entrare nel ciclo cellulare.
2. Inoltre promuove anche il passaggio dalla fase G1 a S mediante
l’induzione dell’espressione della ciclina D1 e della CDK4, comportando
fosforilazione del fattore Rb.
Nabissi2016
Nabissi2016
L’autofagiaèregolatanega7vamentedamTOR,unaserine/threoninekinase,viala
formazionedelcomplessomTORC1complex.
Nabissi2016
InibitoridiPI3KemTOResercitanounforteautophagy-inducingeffect
RAS
• RASèunoncogenecoinvoltonellaproliferazionecellulareed
ècompostodatremembri(H,K,N)-RAS.Sonoproteinelegate
nellatocitoplasmaLcodellacellulaedillororuoloèquellodi
legate,nellostatoinaRvoGDP.
• MutazionipunLformidelgeneRassonol’anomaliasingola
piu’comunedioncogenitrasmessecomecaraGereautosomico
dominante.
Nabissi2016
Queste proteine sono attivate dai RTK e passano continuamente da una
forma inattiva legante il GDP ad una forma attiva legante il GTP.
Le proteine ras sono state scoperte nei virus oncogeni :
K-ras-A, K-ras.B : sarcoma murino, il proto oncogene umano nel
cromosoma 12
H-ras: sarcoma murino, il proto oncogene umano nel cromosoma 11
N-ras: neuroblastoma e carcinomi umani, presente nel cromosoma 1
Il prodotto di questi geni distinti è molto simile e viene denominato p21ras
Nabissi2016
Nabissi2016
Mutazionineigenirashannocomeconseguenzaquelladiridurre
l’aRvità GTPasica, con conseguente mantenimento del GTP e
quindi prodo<o a0vo o mutazioni che facilitano il distacco del
GDP e conseguentemente ras a0vo. Il risultato finale è che ras
rimane piu’ a lungo o cos7tu7vamente a0vo, s7molando la
proliferazionecellularedellecelluletumorali.
Nabissi2016
MUTAZIONE DI KRAS
Inactive
KRAS
active
KRAS
GTP
GAP
Inactive
KRAS
GTP
GDP
Mutant
KRAS
active
GAP
GDP
+
+
Downstream signal transduction cascade
Nabissi2016
mutazioniaRvanLRAS
mutaLoniRASsiriscontranonel30%deitumori
HRAS1
KRAS2
NRAS
R-RAS
Ras-mut=GAP-refractoryGTPase
Un’altaincidenzadimutazionidiK-RASsiriscontrano
neitumorialpancreas(70–100%)
Nabissi2016
Meccanismoditrasduzionediras
Nel citoplasma delle cellule è presente una proteina denominato GRB-2 con
domini SH2/3 che gli perme<ono di formare un complesso con una proteina
denominataSOS(cheèunfa<orediscambiodinucleo7diguanidilici).Quando
lacellulariceveunsegnaledaunfa<oredicrescitachelegaRTK,ilcomplesso
GRB-2 SOS interagisce mediante i domini SH2/3 o PTB (phosphotyrosin
binding)airesiduifosforila7delRTK.Questoconta<operme<eilreclutamento
da parte di SOS della proteina ras (che lega il GDP), questo perme<e lo
scambioconilGTPerassia0va.
Nabissi2016
Questaa0vazionecomportalafosforilazionedellaserina/treoninachinasiraf-1cheasua
voltafosforilaunaMAPKK(MEK)(MitogenAc7vatedProteinKinaseKinase)cheasuavolta
fosforilaunaMAPK(ERK1/2)chetraslocanelnucleo,fosforilafa<oriditrascrizione,come
c-fos,mycea0valaproteinaribosomialeS6,chea0vanolatrascrizionedigenicoinvol7
nell’indurre la proliferazione cellulare o contribuiscono alla crescita cellulare. Quando ras
legailGTPassumeuncambiamentoconformazionalechecomportaancheilreclutamento
delle GTP-asi ac7va7ng protein (GAPs) come la p120GAP e la neurofibromina1 (NF1) che
riportanorasnellaformaleganteilGDP.
NF-1
Nabissi2016
Nabissi2016
PI3-K pathways
Nabissi2016
I pathways RAS, PI(3)K e mTOR formano una rete biochimica
intersecata che, quando mutata, guida la crescita cellulare in
maniera indipendente dall’ambiente circostante. Ques7 pathways
guidano la tumorigenesi a<raverso la fosforilazione coordinata di
proteine che a0vano la progressione del ciclo cellulare, la
regolazione della sintesi di proteine e di fa<ori trascrizionali che
regolanol’espressionedigenicoinvol7inques7processi.
Nabissi2016
Il pathway PI(3)-chinasi.
PI(3)K è attivato sia da RAS che da RTK e di seguito attiva diversi
pathways a valle attraverso la generazione del secondo messaggero
lipidico fosfatidil-inositol-3,4,5-trifosfato (PtdIns(3,4,5)P3. In particolare la
famiglia di chinasi serina-treonina AKT è considerata come il principale
bersaglio di PI(3)K nei tumori.
Esistono tre classi di PI(3)Ks, ma sembra che solo il sottogruppo IA-PI(3)K
sia coinvolto nei tumori. IA-PI(3)K sono eterodimeri composti da una
famiglia di subunità regolatorie (p85) e di subunità catalitiche (p110).
Nabissi2016
Sono capaci di fosforilate l’anello inositolico del lipide di membrana fosfa7dilinositolo 4,5 bifosfato per generare il secondo messaggero PtdIns(3, 4, 5)P3.
Inoltrelesubunitàp85contengonodeidominicheleganospecificiresiduinella
regione citoplasma7ca dei RTKs, questo legame perme<e di generare un pool
locale di PtdIns(3, 4, 5)P3. Le subunità p110 possono legarsi dire<amente ed
a0vareRAS.
Una delle famiglie maggiormente a0vate da PtdIns(3, 4, 5)P3 sono le serine/
treoninechinasiAKT.AKTcontrollailciclocellulare,lasopravvivenzaelacrescita
cellularea<raversolafosforilazionedidiversisubstra7.
Nabissi2016
Nabissi2016
Nabissi2016
Regolazione del pathway PI3K
Nabissi2016
Per i tumori con lesioni genetiche definite,
l’abilità di superare un dato bersaglio
terapeutico è data dalla capacità di acquisire
altre mutazioni che superano l’effetto del
farmaco o effettori biochimici del pathway mutato
o di up-regolare il pathway disregolato.
a) l’attivazione del fattore IF4E e HIF,
inizialmente determinato dal pathway Ras/Akt e
bloccata con Rapamicina risulta inefficiente
causa attivazione di pathways alternativi.
Nabissi2016
b) In tumori con lesioni iniziali in PI(3)K o altri regolatori di mTOR non hanno vie
alternative per attivare IF4E and HIF. Questi tumori rispondono a Rapamicina
Nabissi2016
c) In tumori che esprimono EGFR e ERBB3 inibitori dell’eterodimero
bloccano la via delle IP3K.
d) Nei tumori dove ERBB3 non è espresso la via IP3K è regolata da altre vie
e gli inibitori non funzionano
Nabissi2016
PTENèunafosfatasichelegaedistruggePIP3
convertendoloinPI(4,5)P2oPI(3,4)P2
Nabissi2016
mutazioni
PTEN
Prosta7ccarcinomas–PTENmutatoneglistadiavanza7.
LecellulePTEN-nega7vecellsdiventanoandrigeniresisten7
Etendonoametas7izzare
carcinomamammario:ina0vato15-30%
GLIOMA:mutazionidiPTENsonoindicediaggressività
Carcinomafollicolaredella7roide(25%),
Nabissi2016
EGF RECEPTOR
Nabissi2016
EGF-R
EGF-R ha un ruolo importante nell’ambito di diverse patologie tumorali in quanto
la sua attivazione costitutiva lo rende responsabile dell’attivazione di meccanismi
di proliferazione cellulare. L’omologo del protoncogene EGF-R è stato scoperto
nel virus dell’eritroblastosi aviaria e denominato v-erb. L’analisi della sequenza
aminoacidica di v-erb ha dimostrato che questo oncogene virale codifica per una
proteina (gp65) che è simile ad una parte del EGFR, quindi è un recettore
troncato della porzione extracellulare responsabile del legame a EGF, ma
contenente la porzione citoplasmatica con attività tirosinchinasica che è
costitutivamente attiva. I pathways principalmente attivati dall’attivazione di
EGF-R sono quello di Ras/MAPchinasi e PI3 chinasi/AKT, che agiscono entrambi
nella mitogenesi e sopravvivenza cellulare.
Nabissi2016
ATTIVAZIONE DEI SEGNALI INTRACELLULARI EGFR-DIPENDENTI
1) Legame del ligando specifico nella porzione extracellulare del EGFR
2) Formazione dell’omodimero o eterodimero che causa la fosforilazione
ATP-dipendente di specifici residui di tirosina nel dominio intracellulare di
EGFR
3) Attivazione dei pathways intracellulari:
1) Ras-Raf-MEK-MAPK che controllano la progressione della fase
G1/S
2) PI3K-Akt che attiva una cascata antiapoptotica e di sopravvivenza
Nabissi2016
EGFR è un recettore appartenente ad una famiglia di quattro recettori.
Nabissi2016
Trediversestrategieterapeu7cheperinteragirecontalesistemaeinibirelatrasduzionedei
segnaliprolifera7vi:
• Usodian7corpimonoclonalian7rece<orechesileganoallaporzioneextracellularedella
molecolarece<orialeeimpedisconoillegameconilligando
• Usodifarmaciinibitorisele0vidella7rosinchinasiassociataalrece<ore
• Uso di farmaci che agiscono sulle fasi piu’ avanzata della cascata della trasduzione del
segnalecomeifarmacicheinteragisconoconleproteinedellafamigliaRAS(inibitoridella
farnesiltrasferasi)
Nabissi2016
Meccanismo d’azione dei farmaci Anti-EGFR
Le conseguenze sono:
- Arresto delle cellule in fase G1 con incremento di p27 (inibizione delle CDK)
- Inibizione dell’angiogenesi tramite il blocco della sintesi di TGFa, VEGF, IL-8,
FGF.
- inibizione dell’invasività e delle metastasi
Nabissi2016
Meccanismo dei Anti-EGFR Monoclonal Antibodies.
L’anti-EGFR cetuximab, che è una immunoglobulina IgG1 induce immunorisposta
anticorpo dipendente, citotossicità cellulo-mediata (Panello A). Inoltre gli anticorpi
anti-EGFR inducono internalizzazione del recettore e down-regolazione
incrementando la sua degradazione (Panello B).
Nabissi2016
Nella FISH il centromero del cromosoma 17 e' marcato con un segnale
fluorescente verde e il gene HER-2 con un segnale fluorescente arancio. In
breve si contano circa 60 nuclei cellulari e in ognuno di essi si fa il rapporto tra
segnale verde e segnale arancio.Il tumore e'definito "positivo" per l'amplificazione
genica quando il rapporto gene/cromosoma e'>2
Nabissi2016
Percentuali di tumori EGFR+
TipodiNeoplasia
%espressioneEGFR
Vescica
31-48%
Mammella
14-91%
Utero
90%
Colon
25-77%
Esofago
43-88%
Stomaco
4-33%
Glioma
40-63%
Testa-collo
80-100%
Ovaio
35-70%
Pancreas
30-89%
Prostata
40-85%
Rene
50-90%
Polmone
Nabissi2016
40-80%
INIBITORI DELLE TIROSINCHINASI
Nabissi2016
REGOLAZIONE DELL’ATTIVITA DELLE TK
TKs sono divise in due classi, recettori TKs transmembrana con dominio attivatore
extracellulare e dominio catalitico intracellulare e TKs citoplasmatiche che mancano
del dominio extracellulare che si trovano nel citoplasma, nucleo e membrane
interne. L’attività delle TK è regolata in modo tale che in condizioni non proliferative
sono defosforilate, perchè è assente il ligando.
Le TK-non recettori associate sono mantenute in uno stato inattivo da inibitori
cellulari e stimoli di diversa natura dissociano questi inibitori e inducono la
fosforilazione mediante altre chinasi. L’azione delle chinasi è terminata dalle
fosfatasi che idrolizzando il fosfato presente sulla tirosina ed inducono gli inibitori.
Nabissi2016
Nabissi2016
Un meccanismo comune nei tumori ematologici è l’attivazione delle TK
mediante la fusione di un recettore o non recettore TK con una proteina
partner, come conseguenza di una traslocazione cromosomica. Un aspetto
frequente della proteina partner è un dominio che causa una
oligomerizzazione costitutiva della TK in assenza di ligando, promovendo
autofosforilazione ed attivazione.
BCR-ABL nella CML (leucemia mielodie cronica), dove si genera una
proteina chimerica che costituisce una TK costitutivamente attiva.
Mutazioni della chinasi FLT3 nella leucemia mieloide acuta (AML) che
rende la TK attiva in assenza di ligando.
Nabissi2016
Leucemia Mieloide Cronica (LMC)
IlcromosomaPhiladelphia
1
6
2
7
3
8
13
14
19
20
4
9
15
10
16
21
22
5
11
12
17
18
x
Y
Presenteinoltreil95%deipazien7affe0daLMC
Nabissi2016
Rappresentazione schematica
del cromosoma Philadelphia (Ph)
Leucemia Mieloide Cronica
Latraslocazionet(9;22)
Cromosoma
Cromosoma
9
9+
Cromosoma
Cromosoma
22
Ph
bcr
Bcr-Abl
abl
Proteinadifusionecon
elevataaRvità
Lrosina-chinasica
La fusione BCR-ABL durante la traslocazione e’ associata a un notevole
aumento dell’attività tirosinchinasica di ABL e da’ a quest’ultima la
capacità di legare il sito
SH2 di attacco a GRB2.
Nabissi2016
Nabissi2016
Meccanismidiresistenza
secondariaall’ImaLnib
• MutazionidiBCR-ABL
• Aumentatoefflussodallacellula(glicoproteina
MDR)
• IperespressionediBCR-ABL
• Alterazionigene7cheaggiun7ve
Nabissi2016
Nabissi2016
ONCOSOPPRESSORI
Nabissi2016
Genioncosoppressori
GenilacuimancataaRvitàall’internodellacellulafavoriscelacrescitatumorale.
L’alterazionenellastruGuraonell'espressionediquesLgeni(inaRvazione)seèacarico
dientrambigliallelideterminaperditadifunzioneefenoLpotumorale.
●Inibitoridellacrescita(TGF-beta)
●ReceGorediinibitore(TGF-betaReceptor)
●InibitoreditrasduGoredelsegnale(NF1,PTEN)
●InibitorideifaGoreditrascrizione(Rb)
●Inibitoridelciclocellulare(CDKI,p16,p21)
●FaGoriapoptoLci(Bad,Bax)
Nabissi2016
●Funzionedicontrollodell’integritàdelgenoma(p53)
CorreGaduplicazionedelgenoma
RiparazionedelDNA
•BRCA-1Integritàdelgenoma(stabilitàcromosomiriparazioneDNA)
•BRCA-2Integritàdelgenoma(stabilitàcromosomi,riparazioneDNA)
•NERIntegritàdelgenoma(riparazioneDNAperescissionedeinucleoLdi)
•MSH2 Integritàdelgenoma(riparazioneDNAdaerratoappaiamento)
ATMIntegritàdelgenomaaRvasistemiriparazione,aRvap53,rallentaciclo
mitoLcoinfaseG2
Nabissi2016
MeccanismidiinaRvazionedigenioncosoppressori
PerditadelprodoGooformazionediprodoGoinaRvo
(troncato)acausadi
●Delezione(perditagene,perditainterocromosoma)
●Mutazione(nonsenso,disenso,frameshiq)
●EpigeneLca(esme7lazione)(ipermeLlazionedel
promotorediMLH1impedisceilmismatchrepair)
●ProdoRvirali
Nabissi2016
TP53
Nabissi2016
TP53
p53 funziona principalmente come
fa<ore trascrizionale e controlla
l’espressionediunampiospe<rodi
geni, coinvol7 nelle piu’ disparate
funzioni.
p53èspessomutatoneitumori.
Mutazioni che producono un p53
troncato o malfunzionante o
mutazioni che interferiscono con il
D N A B i n d i n g D o m a i n s o n o
predizionediunapessimadiagnosi.
Nabissi2016
Perquantolaperditaomutazioniinp53siaassociateadunincrementodellasusce0bilità
a sviluppare un tumore, topi -/- per p53 sviluppano normalmente e altre osservazioni
indirizzano p53 anche verso un ruolo nella normale fisiologia cellulare. Ques7 studi
includono il ruolo di p53 nel regolare la longevità e l’invecchiamento, la glicolisi e la
risposta apopto7ca dopo ischemia, la sopravvivenza cellulare dopo danni genotossici o
stressossida7vo,angiogenesi,rimodellamentoosseoedautofagia.
Nabissi2016
TP53 Mutazioni Germinali predispongono
a diversi tipi di cancro
Tumor spectrum in TP53 mutation carriers Breast
28.9
16 SonLssues
>80%
15.4
Brain Bones
12.8
6.8
Adrenalgland Lung Leuk/Lymph.
Stomach 3.4
3.2
2.5
1.7
Skin 1.5
Ovary 1.5
Other
5.9
0 Colorectum
5 10 Nabissi2016
15 20 25 30 TP53 è un oncosoppressore
% topi con tumore
p53+/+
1% at 18 mesi
p53+/-
2% at 9 mesi
p53-/-
75% at 6 mesi
Nabissi2016
Mutazioniinp53
Nabissi2016
• TADN-terminal[aa1-42]
TADregolazionenegaLvaaGraversointerazioneconMDM2
Domino transaRvante (TAD): a0va geni che codificano per prodo0 che esercitano un
ruoloinibitoriosullaprogressionedelciclocellulare(p21/WAF),edinibisconol’espressione
digenicoinvol7nellaprogressionedellafaseG1/S(genidellecicline).
La regolazione dell’espressione di ques7 geni avviene quando p53 trasloca nel nucleo,
previa fosforilazione del residuo di serina 316 (ser316), svolta da specifiche chinasi.
ConLeneancheilsitodilegameperMDM2
Nabissi2016
• Pro-richregionfraTADeDBD
– PxxPpresen75zonenellaregione61-94
– DelezionedellaP-richregion➨riducelarispostaapopto7caeilbloccodel
ciclocellulare,manonlanormalerispostatrascrizionale
– Con7eneresiduichediventanofosforila7dopolarispostaapopto7ca
Regione ricca in prolina: regione ricca di aminoacidi basici, che a0va7 in seguito a
fosforilazioneperme<eillegamealDNAtramiteilDNABindingDomain
Nabissi2016
p53DBD
2“α-helicalloops”checonta<anoilDNA
Altrestru<uredeputate
AllegamealDNA
Nabissi2016
MutazioniallaDBD
• Lamaggiorpartedelle
mutazionichecausanoil
tumoresitrovanonella
DBD
– Destabilizzandole
interazioni.
– p53legailDNAcome
tetramero
Nabissi2016
Dominioditetramerizzazione
2β+2αstruGura
LegataallaDBDvia37aa
Nabissi2016
C-terminal
allostericdomain
DiversievenLnellaregioneC-terminale
possonoriaRvarelaregioneDBD
DominioleganteilDNA:perme<el’interazioneconilDNA
Nabissi2016
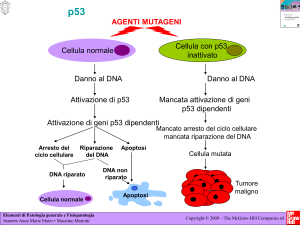
ATTIVAZIONE di p53
Ilfunzionamentodip53èregolatodaalcuneproteineeffeGrici:ATMeATR,
le
qualiriconosconoilDNAdanneggiato.
ATMeATRaRvanolechinasiCHK2eCHK1.
CHK1aGraversounaseriediprocessiinibisceleCDKs,mentreCHK2come
ATMstessoaRvanop53,ilqualeasuavoltaaRvap21/waf(bloccandola
proliferazione),aRvagenipro-apoptoLci(Bad,Bax,PUMA,…)edaRva
GADD45(growtharrestandDNA-damageinducible).
SeilDNAvieneriparatop53aRvailsuoinibitoreMDM2,cheasuavoltalega
p53eloportaadegradazione.
Seildannononvieneriparatop53aRvailprocessodiapoptosi.
Nabissi2016
ATMeATRfosforilanop53arrestandolafaseG1/S
Nabissi2016
nabissi14
• ImeccanismichecontrollanoilpassaggiodallafaseG1allaS,consistono
anche di processi d’inibizione della proliferazione cellulare, i quali
regolanol’aRvitàdeicomplessiC/CDK.
• Ledueprincipaliclassid’inibitorisonofaGoriproteicidellafamigliaCip/
KideINK4/Arf.
• LafamigliaCip/Kidècompostadatremebri:p21Waf,p27ep57.
• p57 funziona come freno all’interno del ciclo cellulare, mentre p21/waf
sonoprincipalmentedeimediatoridisegnalicitostaLci.
• Ink4eArfagisconorispeRvamentebloccandoilciclocellulareedagendo
sulfaGoretrascrizionalep53.
Nabissi2016
Inibizione di P53
MDM2 associata con p53 TAD
MDM2-binding comporta
– 1. Repressione della transattivazione
– 2. Destabilizzazione di p53 in quanto MDM2-binding stimola
degradazione di p53
Nabissi2016
p53andMDM2formanounfeedbackloopautoregolatorio.
p53 s7mola l’espressione di MDM2; MDM2 inibisce p53 bloccando la sua a0vità
trascrizionale,favorendolasuadegradazione.
DNAdamagefavoriscelafosforilazionedip53,prevenendolasuaassociazioneconMDM2.
ARF,previeneladegradazionedip53MDM2-mediata.
Quindi inibitori dell’interazione p53–MDM2 possono a0vare p53 nei tumori esprimen7
p53.
Nabissi2016
Diversestrategieperrompereillegame
MDM2-p53
Primadell’aRvazione
aRvato→
fosforilazione→
RoGuradellegame
aRvazione→
fosforilazione→
InaRvatal’azionediE3
AcLvated→
ARF-binding→
inacLvatedE3-act
Nabissi2016
SelariparazionedelDNAnonvieneeffeGuataallorap53
puo’indurreapoptosiaGraversol’aRvazionedigeni
proapoptoLcidellafamigliaBcl-2,Faserepressionedi
genianL-apoptoLci.
L’azionedip53comefaGoreresponsabiledelbloccodel
ciclocellulareodell’aRvazionedell’apoptosisembrasia
regolatadaisuilivelliall’internodellacellula.
BassilivelliinduconobloccodelciclocellularementrealL
livellil’apoptosi.
Nabissi2016
Celldeath(Apoptosis)byp53
Nabissi2016
Bersaglio:Microambiente
Lanecessitàperitumoridirifornirsidinutrientepuo’esserecontrastatacon
farmacianLangiogeneLci,comenell’usodianLcorpimonoclonalicontroil
VEGF,neltraGamentodelcancroalcolon,maèun’approcciochepotrebbe
essereapplicatoinmolLtumori.
Inoltrelaterapiacheagiscepiu’sull’ospite(celluleendoteliali)chesultumore
èsicuramentemenosoggeGaaresistenzafarmacologica,inquantolecellule
normalehannosicuramentemenoplasLcitàgeneLcadelletumorali.
StudirecenLriguardanLleinterazionitumore-stromarivelanouncomplesso
scambiod’informazionichenonriguardanosololavascolarizzazione.
LostromacomprendeoltreaifibroblasLunnumeroelevatodicellule
infiammatoriechepossonochiaramenteinfluenzarelacrescitadeltumore.
UnodeifaGoristromalimaggiormentestudiatoèTGF-β, ilqualesvolge
diversiruolichepossonoinfluenzareposiLvamenteonegaLvamentela
crescitatumorale.
Nabissi2016
TGF-beta(TGF-β)
Neitessu7,l’omeostasirichiedeunbilanciamentodelleinterazionifralecellule
e la matrice extracellulare. Queste interazioni coopera7ve coinvolgono
numerose citochine che agiscono a<raverso specifici rece<ori di membrana.
Quando il bilanciamento tra cellule e matrice extracellulare è perturbato si
possono sviluppare diverse patologie. Questo fenomeno è par7colarmente
evidenteperilTGF-β cheèunmembrodellafamigliadifa<oridicrescitache
includelaproteinamorfogeneLcaosseael’acLvina.Quasitu0i7pidicellule
producono TGF-β ed hanno i rece<ori per il TGF-β il quale regola la
proliferazione e la differenziazione delle cellule, lo sviluppo embrionale, la
riparazionedelleferiteel'angiogenesi.
Nabissi2016
IlruoloessenzialedelTGF-β,dimostratoindiversiesperimen7,dimostrachel'incremento
o decremento di TGF-β è legato a diversi 7pi di patologie, fra cui il cancro. Ci sono tre
isoformediTFG-β(1,2,3)ognunacodificatadagenidis7n7mastru<uralmentealtamente
conservatemadifferen7nell’affinitàdilegamealrece<ore.IlTGF-βèsinte7zzatocome
unlargoprecursorecontenenteunaregionechevienetagliataprimadellasecrezionedel
precursore,macherimanea<accatamediantelegaminoncovalen7.Unavoltasecretoil
TGF-β viene immagazzinato nella matrice extracellulare, in un complesso comprendente
unaproteinadilegame(TGF-βbindingprotein,TGFBP),cheneevitaillegamealrece<ore.
IlTGF-βvienepoirilasciatomedianteunaglicoproteinadimatrice(trombospondina-1)la
qualecambialaconformazionedellaTGF-βBP.IlTGF-βa0voagiscesutre7pidirece<ori
(I,II,III),mai7piIeIIsonoquellispecificisoloperTGF-β.
Nabissi2016
TGF-βèimmaganizzatonellaECMinformaina0va
complessatoconbeta-glicaniedecorina
precursore
maturo
Dopoiltaglio
TGF-β
formaomodimerivialegami
S-S
TagliatomaancoralegatoallaECM
TGFBP
TGF-βa0vopuo’essererilasciato
tramiteacidificazione
ContaGocellula-cellula,proteasi,
trombospondina
Nabissi2016
I rece<ori di 7po II e II contengono delle chinasi serina-treonina nel loro dominio
intracellulare che a0vano la fosforilazione di diversi fa<ori di trascrizione conosciu7
come SMAD (10 membri). SMAD2 e SMAD3 sono fosforila7 mediante a0vazione del
TGF-RI, SMAD4 è un partner comune di tu0 gli SMAD a0va7 da rece<ori, mentre
SMAD6eSMAD7bloccanolafosforilazionediSMAD2eSMAD3,inibendocosìilsegnale
a0vatodaTGF-β. Unmeccanismogeneraledell’azionedelTGF-β,consistenellegame
del TGF-β al TGF-RII che recluta, lega e fosforila il TGF-RI il quale s7mola l’a0vità
chinasica. che fosforila SMAD2 o SMAD3 i quali formano un complesso che migra nel
nucleoelegaaltrifa<oritrascrizionaliregolandolatrascrizionedispecificigeni
Nabissi2016
TGF-β/SMAD
Nabissi2016
RuolodelTGF-βnelcancro
Regolazionedelciclocellulareedeffe<osullaproliferazione.
Nellecelluleematopoie=che,endotelialiedepitelialiilTGF-βèunpotenteinibitoredella
proliferazionecellulare,s7molandolasintesidelleCDKI(p15)edinibendolefunzioniela
produzionedelleCDK2,CDK4,ciclinaAedE.Ques7effe0risultanoinunariduzionedella
fosforilazionediRb,bloccandocosi’l’a0vazionedeifa<oriE2F.Neitumorilemutazioni
acaricodelpathwayTGF-β,comportanounaproliferazioneincontrollata.Questaperdita
nel controllo della proliferazione ha come conseguenza anche un’induzione della
secrezione di TGF-β da parte dei fibroblas7, che agisce (essendo le cellule tumorali
resisten7)suglistessifibroblas7,sulsistemaimmunitario,sullecelluleendotelialiesulla
muscolatura liscia causando immunosoppressione, angiogenesi e s7molando
l’invasivitàdeltumore.
Nabissi2016
Nabissi2016
SLmolal’espressionedeiCDKI
comep15ep21ereprime
l’espressionedic-myc
TGF-β
Nabissi2016
L’arrestoèmediatodalladownregolazionediMyc,ilquale
rilasciaMiz1.Miz1silegaal
promotoredelp15eloaKvain
co-operazioneconilcomplesso
Smad.
TopiSMAD4+/-sviluppanopolipiduodenaliegastricicon
abbondantestroma
edinfiltrazionidieosinofilia6-12mesi
CarcinomiSmad4+/-Apc+/-
mostrano
proliferazionedellecellulestromali
eforteinvasioneLssutale.
Nabissi2016
EffeGosullemetastasi
TGF-β è uno dei piu’ poten7 s7molatori della produzione e deposizione della matrice
extracellulare. S7mola la produzione e influenza le proprietà adesive della matrice
extracellulare,a<raversoprincipalmenteduemeccanismi.TGF-βsLmolaifibroblasLed
altrecelluleaprodurreleproteinedellamatriceextracellulareeleproteinediadesione
cellulare,inclusoilcollagene,fibronecLnaedintegrine.Secondo,TGF-βdecrementala
produzione di enzimi che degradano la matrice extracellulare, incluso collagenasi,
eparinasi e stromelisine ed incrementa la produzione di proteine che inibiscono gli
enzimi che degradano la matrice extracellulare come gli inibitori dell’aRvatore del
plasminogenoegliinibitoridellemetalloproteasi(TIMPs).
Nei tumori TGF-β è aumentato e la sua azione induce sLmolazione delle molecole di
Nabissi2016
adesionecellulare,sLmolandol’invasivitàel’angiogenesi.
Cosasuccedeallecelluletumorali
quandoTGF-βpathwayèspento(viaperditadiSMAD
oTGFB-R2)
Lecellulenonsonopiu’responsiveaTGF-βmacon7nuano
aprodurloerilasciarlo
Celluledicarcinoma
sonoimmerse
nellecellulestromalinormali
interagendo
1. ProduzionelocalediTGF-β
sopprimelarispostaimmunitariaanLtumorale
2.TGF-βsLmolal’angiogenesi
TGF-beta1èspessoelevatonelplasmadipazienLcontumorealseno,polmone,fegatoe
prostata
Nabissi2016
EffeRimmunosoppressivi
TGF-β che è prodo<o da tu0 i leucoci7, promuove la loro differenziazione ed
inibiscelaproliferazioneeda0vazione.
Sopprimendolaproliferazioneleucocitaria,induceunadiminuzionedicelluledel
sistema immunitario, suggerendo un ruolo del TGF-β nell’aiutare il tumore a
sfuggiredallaimmunosorveglianza.
Nabissi2016
Nabissi2016
VIRUS ONCOGENI
• Le par7celle virali o virioni, hanno piccole dimensioni (da 0,02 a 0,4 µm) e una
sempliceorganizzazionestru<urale.
• Sono cos7tui7 da piccole par7celle formate essenzialmente da solo materiale
gene7co (RNA o DNA), de<o core, avvolto da membrane prote0ve, quali capside e
pericapsidedinaturalipoe/oglicoproteico,chehaunaduplicefunzionediproteggere
ilgenomaemediarelapenetrazionenellacellulaospite.
• La mancanza di ribosomi e di sistemi enzima7ci deputa7 alla produzione di energia
(ATP)fasicheilprocessodireplicazionepossaavveniresoloquandoilgenomavirale,
penetrato nella cellula ospite, si spoglia della capsula ed inizia il suo processo
replica7voconilcontributodellacellulaospite.
Nabissi2016
Molti virus hanno invece un secondo rivestimento, proveniente dalla
membrana cellulare della cellula ospite e formato da fosfolipidi. Questo
secondo rivestimento è chiamato envelope, su cui sono evidenti le
molecole necessarie per l’infezione.
Nabissi2016
Virus oncogeni
Le caratteristiche generali che sono rappresentative della trasformazione
virale delle cellule eucariotiche sono:
1) la singola interazione di una particella virale con la cellula suscettibile è
sufficiente per indurre la trasformazione
2) La trasformazione induce una modificazione genetica spesso
irreversibile nella cellula infettata, in quanto il genoma virale s’integra
stabilmente nel DNA cellulare.
Per alcune forma virali il DNA virale rimane nella cellula ospite in forma
episomiale (DNA non integrato).
Nabissi2016
3) La trasformazione comporta l’espressione dei
geni virali necessari al mantenimento del DNA
viralenellacellulainfe<ata.
4) Le cellule trasformate possono rilasciare il virus,
esprimerean7genivirali(nucleari,citoplasma7cio
d i m e m b r a n a ) , a c q u i s i r e u n f e n o 7 p o
immortalizzato (perdere i meccanismi di controllo
dellaproliferazioneecrescereinmodoindefinito).
Nabissi2016
Cancroindo<oda
virusaDNA
ONCOGENE
•Ungenechepotenzialmentetrasformalecellulenormaliin
cellulecancerose
•Quandol’oncogeneètrasmessodavirus
ONCOGENEVIRALE
v-onc
Nabissi2016
• Virus oncogeni a DNA
I virus oncogeni a DNA appartengono a cinque famiglie:
PAPOVAVIRUS: PA (papilloma), PO (polioma), VA (virus vacuolizzante)
ADENOVIRUS: isolati per la prima volta dalle adenoidi
HERPESVIRUS: induce delle striscianti progressioni di lesioni
erpetiche (herpes dal greco strisciare)
POXVIRUS: deriva dalla parola inglese pock (pustola)
HEPADNAVIRUS: deriva da HEPA(tic), DNA, virus
Nabissi2016
• Il comportamento dei virus dipende dalla permissività della
cellula ospite, infatti nelle cellule permissive in cui viene
permessa la replicazione delle particelle virali, dopo
integrazione del genoma virale in quello dell’ospite si ha
una infezione di tipo produttivo con effetto citopatico litico e
quindi morte della cellula. Di conseguenza non si ha
trasformazione cellulare con possibile sviluppo di cellule
tumorali.
Nabissi2016
• Nelle cellule non permissive non si ha produzione delle particelle virali
necessarie a completare il ciclo replicativo del virus e quindi i pochi
geni virali espressi possono spingere la cellula verso la
trasformazione. La trasformazione puo’ contribuire allo sviluppo di
tumori in quanto puo’ indurre la modifica di meccanismi cellulari
(proliferazione, apoptosi, sopravvivenza, immunosoppressione) grazie
all’integrazione di porzioni di DNA virale nel DNA della cellula ospite
che avrà introdotto un nuovo gene o alterata l’espressione di geni
preesistenti, inoltre la cellula trasformata non rilascia mai il virus
infettivo.
Nabissi2016
Nabissi2016
• Nell’ambito del processo di trasformazione
numerose proteine virali interagiscono con le
proteine cellulari dell’ospite formando dei complessi
che modificano i normali processi regolatori cellulari,
spesso regolando attivando proto-oncogeni o
disattivando le funzioni dei proto-oncosoppressori.
Nabissi2016
•
•
PAPOVAVIRUS
Famiglia di virus di piccole dimensioni con DNA bicatenario racchiuso in un
capside privo di envelope, costituita da due generi: i poliomavirus ed i
papillomavirus.
•
Il genoma dei poliomavirus è costituito da 7 geni codificanti proteine precoci e
tardive, le tardive sono proteine virali strutturali e quindi non vengono mai
espressi nelle cellule trasformate. Le proteine precoci (antigene T piccolo,
medio e grande),sono necessarie per la replicazione del DNA virale nelle cellule
permissive e per la trasformazione. Nelle cellule trasformate l’antigene T medio
si associa alla membrana plasmatica ed attiva la protein-chinasi c-src, antigene
T piccolo si localizza nel nucleo mentre il T grande è in parte associato alla
membrana plasmatica diventando bersaglio dei linfociti T citotossici.
•
Comunque l’infezione da poliomavirus non è attualmente associato a tumori
umani, pur essendo presenti in diverse patologie umane regioni geniche del
virus.
Nabissi2016
• PAPILLOMAVIRUS
La particella del Human Papillomavirus (HPV) consiste di un DNA circolare
di 8000 bp racchiuso in un capside composto da due molecole (L1 e L2). Il
genoma ha la capacità di codificare per queste due proteine e per almeno
sei proteine precoci (E1, E2, E4-7) che sono necessarie per la replicazione
del DNA virale e per l’assemblamento di nuove particelle virali all’interno
della cellula infettata. I due gruppi di geni sono separati da delle regioni
regolatrici (URR) di circa 1000 bp che non codificano per proteine ma
contengono cis-elementi necessari per la regolazione genica, la
replicazione del genoma e per il suo impacchettamento nelle particella
virali
Nabissi2016
HPVRESPONSABILEDELLAFORMAZIONEDELLEVERRUCHE
Nabissi2016
Nabissi2016
• Il ciclo infettivo del HPV inizia quando le particelle infettive raggiungono lo
strato basale dell’epitelio, dove esse si legano ed entrano nelle cellule,
attraverso piccole lesioni. Il ciclo replicativo all’interno dell’epitelio puo’
essere suddiviso in due parti:
• il genoma virale viene replicato in circa 100 copie e mantenuto per un
periodo di tempo variabile a questo basso numero di copie all’interno delle
cellule permissive. Le proteine E1 e E2 sono essenziali per questa
replicazione del DNA.
• Nella seconda fase le cellule basali sono spinte verso il compartimento subbasale, perdono la loro capacità di dividersi ed iniziano il loro programma di
differenziamento. Il HPV replica in questo compartimento e per il suo rilascio
nell’ambiente extracellulare approfitta della disintegrazione delle cellule
epiteliali che avviene in conseguenza del loro naturale turn-over nello strato
superficiale
Nabissi2016
• Le proteine critiche nel processo di replicazione virale sono le proteine
E6 ed E7 che interagiscono con diverse proteine cellulari.
• Pur essendoci differenze fra le proteine E6/E7 nei sottotipi HPV ad alto
e basso rischio, le principali interazioni caratterizzate delle proteine E6/
E7 sono con le proteine cellulari p53 ed Rb, le quali sono molecole
centrali nel controllo del ciclo cellulare. Il legame di E7 a Rb attiva il
fattore trascrizionale E2F il quale attiva la trascrizione di geni coinvolti
nella replicazione del DNA. La proteina virale E6 interagisce ed inattiva
(portandolo a proteolisi) il fattore trascrizionale p53.
Nabissi2016
• La conseguenza di questa infezione è la perdita del controllo del ciclo
cellulare, della riparazione del DNA ed il rallentamento del processo
differenziativo delle cellule epiteliali.
• L’abilità del HPV di persistere ed indurre progressione verso la
malignità puo’ essere spiegata da una particolarità di questo stadio del
suo ciclo replicativo. La costante attivazione delle proteine E6/E7
portano ad un incremento dell’instabilità genomica, perdita del controllo
del ciclo cellulare ed in ultimo al cancro. Durante la progressione del
tumore il genoma virale s’integra spesso nel genoma della cellula
ospite con il risultato di una costante espressione delle proteine E6/E7
mediante stabilizzazione del loro trascritto (mRNA), grazie all’influenza
sulle modifiche della cromatina o mediante la perdita della regolazione
negativa della trascrizione mediata dalla proteina virale E2.
Nabissi2016
Nabissi2016
• Gli HPV attualmente caratterizzati sono piu’ di 100, di cui 40 capaci di
infettare le mucose del tratto genitale e sono caratterizzati come a basso
rischio ed alto rischio in conseguenza della loro prognosi clinica. Quelli a
basso rischio sono principalmente associati con le lesioni anogenitali
mentre quelli ad alto rischio con i tumori ano genitali. Due dei tipi a basso
rischio (HPV-6 e HPV-11) causano la maggior parte delle lesioni ano
genitali e dei papillomi respiratori ricorrenti. Le infezioni con HPV ad alto
rischio causano virtualmente il 100 % dei tumori alla cervice, 90 % dei
tumori all’ano, 50 % di quelli alla vulva, vagina e pene. HPV-16 e HPV-18
od entrambi sono responsabili del 70 % dei tumori della cervice.
Nabissi2016
•
La progressione dell’infezione da HPV a cancro alla cervice è accompagnata da
una sequenza di cambiamenti istologici. La neoplasia intraepiteliale cervicale
(CIN) è un’anormalità istologica dell’epitelio squamoso della cervice che è
associata con l’infezione da HPV ed è riconosciuta come un potenziale
precursore del tumore alla cervice. La CIN è classificata in tre gradi:
•
CIN 1: presenza di una leggera displasia, con presenza di cellule anormali
•
CIN 2: moderata displasia con maggiore presenza di cellule anormali
•
CIN 3: displasia severa con cellule anormali che occupano la maggior parte
dell’epitelio della cervice.
Nabissi2016
Nabissi2016
• Nel caso di CIN 2 e 3 la persistenza della displasia è associata con lo
sviluppo del cancro, con percentuali rispettivamente del 57 % e 70 %.
• I due vaccini che sono stati sviluppati prevengono l’infezione primaria da
HPV e sono costituiti dalle proteine L1 assemblate fra loro in particelle
virali-simili che sono morfologicamente identiche al virione HPV ma non
contengono il genoma virale. Cosi’ il vaccino induce una risposta
anticorpale virus-neutralizzante. I due vaccini contengono uno particelle
virus simile dei ceppi HPV 6-11-16-18 mentre il secondo per HPV 16-18.
Nabissi2016
• Patogenesidell’epaLteB.
• HBV non è dire<amente citotossico per le cellule, infa0 mol7 portatori del
virus HBV sono asintoma7ci o hanno lievi danni epa7ci per quanto vadano
incontroareplicazioneintraepa7cadelvirus.Quindisipensachelarisposta
immunitaria dell’ospite sia il principale determinante del danno epa7co,
infa0 pazien7 con dife0 immunitari che sono infe<a7 da HBV mostrano
spessolievidanniepa7ciacu7maaltapossibilitàdiandareincontroadanni
cronici.LarispostaimmunitariaaHBVcoinvolgerispostamediatadalinfoci7
TMHC-classeIICD4+helpereMCH-classeICD8+dire0controdiversiepitopi
del core ed envelope dell’HBV, mentre nei portatori cronici la risposta
linfocitariaèa<enuata,conunamaggiorepresenzadian7corpian7-HBs
Nabissi2016
Questo tipo di risposta suggerisce che la maggior parte del danno epatico
sia dovuto piu’ ad una risposta antigene-non specifica infiammatoria
secondaria data dal rilascio di prodotti citotossici come TNF, ROS,
proteasi e cellule natural killer (NK).
Nabissi2016
• Carcinoma epatocellulare
• Un altro aspetto dell’infezione da HBV è dato da un alto rischio di
sviluppare carcinoma epatocellulare nei pazienti infettati cronici, con un
incidenza 100 volte superiore a quella dei non-portatori.
• Terapia
• Il successo della terapia nei pazienti con infezione da HBV è nella
riduzione dei livelli di viremia (livelli di antigene HBe) e delle disfunzioni
epatiche (valutabili dai livelli di aminotransferasi).
• Attualmente comunque la scomparsa completa del virus si ha solo nel
5% dei pazienti, sebbene lo sviluppo di nuovi antivirali potrebbe
incrementare questo dato.
Nabissi2016
• Interferone
• Per molti la somministrazione d’interferone alfa è stata la terapia principale,
con una risposta positive nel 30 % dei casi (perdita di HBeAg), sviluppo di
anticorpi anti-HBe e riduzione dei livelli di aminotrasferasi. Tuttavia gli effetti
collaterali, dati in parte dalla stimolazione interferone alfa indotta,di antigeni
MHC classe I sugli epatociti, con conseguente attività citotossica da parte
dei linfociti T o effetti come febbre, mialgia, trombocitemia e depressione
hanno reso difficile l’uso di tale trattamento per molti pazienti.
• Farmaci antivirali
• Analoghi nucleosidici o nucleotidici (ex. Lamivudine), che bloccano la
replicazione virale mediante inibizione dell’attività delle trascrittasi inversa
(RT) senza dare hanno effetto immunomodulatore ma la terapia induce
resistenza farmacologica, mediata da mutazioni puntiformi nel sito catalitico
della RT.
Nabissi2016
Herpesvirus
8Herpesvirusumani
VirusaDNAlineareadoppiofilamento
Possonocausareinfezionili7che,persisten7,
laten7/ricorren7,eimmortalizzan7(EBV)
Alpha-herpesvirus:HSV1,HSV2
VirusdellaVaricellaZoster(VZV/HHV3)
Beta-herpesvirus:Citomegalovirus(CMV/HHV5)
ViruserpeLciumani6e7(HHV-6e7)
Gamma-herpesvirus:Epstein-Barr(EBV/HHV4)
Viruserpe7coumano8(HHV8)
Nabissi2016
La cara<eris7ca peculiare di questa famiglia è la possibilità di
laten7zzare (VIRUS SILENTE, CHE POI PUO’ RIPRENDERE LA FASE
INFETTIVA),inseguitoall’infezioneprimaria.
A distanza di tempo l’infezione si può ria0vare (infezione ricorrente)
con patologie che possono essere anche differen7 dall’infezione
primaria.
Le infezioni sono spesso asintoma7che ma possono assumere decorso
moltosfavorevoleinindividuiimmuno-compromessi
Nabissi2016
INFEZIONELATENTE
HERPESVIRUSTIPO1:dopoinfezioneprimariasilocalizza
neiganglidelnervotrigemino,persistendoinpiu’copiein
formaepisomialeestabilendounainfezionelatenteper
tu<alavita.
S7molipatogenio“stress”possonoria0vareilvirusche
giungeallegiunzionineuro-epitelialieinfeGalecellule
epiteliali,producendolalesioneerpeLca.
Nabissi2016
V I R U S R E S P O N S A B I L E D E L L A
HEPSTEINBARRVIRUS(HBV)
MONONUCLEOSI. SI MOLTIPLICA
NELLE CELLULE EPITELIALI NASOFARINGEEEDINFETTAILINFOCITIB,
I N F O R M A L A T E N T E E D
EPISOMIALE.
ESPRIMEPROTEINEVIRALI(EBNA-1)
CHEINDUCELADNAPOLIMERASIA
PRODURREPIU’COPIEVIRALI.
A L T R E E B N A I N D U C O N O
L’IMMORTALIZZAZIONE DEI LB E
PROTEINE DI SUPERFICIE CHE
I N D U C O N O U N A R I S P O S T A
CELLULO-MEDIATA DEI LINFOCITI T
CITOTOSSICI CHE DISTRUGGONO I
LBINFETTI.
Nabissi2016
CORRELAZIONECONILLINFOMADIHODGKIN
EBVE’TIPICODEIGIOVANIADULTIMENTRENELL’INFANZIAE’SPESSOASINTOMATICA.
LA MALATTIA SI MANIFESTA CON FEBBRE, FARINGITE, INGROSSAMENTO DEI
LINFONODI,SPLENOMEGALIA,ALTERAZIONIDELLEFUNZIONIEPATICHE.
TRASMESSO ATTRAVERSO SECREZIONI FARINGEE, SPESSO TRAMITE SCAMBIO DI
SALIVA.
SEDE PRIMARIA LE CELLULE EPITELIALI DELLA FARINGE, POI LINFOCITI B DOVE
S’INSTAURALALATENZA.
NEILBPUO’INDURREIMMORTALIZZAZIONE
ASSOCIATOANCHEALCARCINOMANASOFARINGEO
Nabissi2016
VirusdellaVaricellaZoster(VZV)
DETERMINALAVARICELLA,MOLTOCONTAGIOSAEDIFFUSA,TIPICADELL’INFANZIA
SITRASMETTECONLESECREZIONIDELLEVIERESPIRATORIEOMEDIANTECONTATTO
CONIFLUIDIVESCICOLARI
INCUBAZIONECIRCA14GIORNI
VZVSIMOLTIPLICANELLEVIEAEREESUPERIORIEPOIDIFFONDENEILINFONODI,POI
SEGUEUNAVIAINFETTIVA(FASEVIREMICA)CONINFEZIONEDELFEGATO,MILZA,RENI,
EPIDERMIDE)
NELL’EPIDERMIDEPROVOCALEMACULE,CHEEVOLVONOINPAPULE,VESCIOLE,
PUSTOLE.
PUO’RIMANEREINUNAFASEDILATENZAAVITA(NEIGANGLISENSORIDORSALI).
LARIATTIVAZIONEPROVOCAUN’ERUZIONEVESCICOLAREDOLOROSA(FUOCODI
SANT’ANTONIO),GENERALMENTENELLAREGIONETORACICA.
Nabissi2016
Nabissi2016