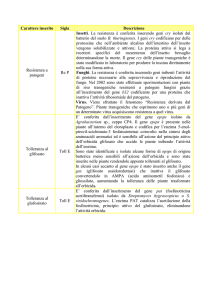
PIANTE GENETICAMENTE MODIFICATE
METODI DI TRASFORMAZIONE GENETICA
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
METODI DI TRASFERIMENTO GENICO
Fasi richieste per ottenere piante transgeniche:
1.clonaggio del gene
2.incorporazione del gene in un vettore
3.incorporazione del gene nel DNA della specie ricevente e
selezione delle piante trasformate
4.verifica dell'incorporazione del gene e della capacità di
esprimere le sue funzioni nella
specie ricevente.
2
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
METODI DI TRASFERIMENTO GENICO
Incorporazione del gene in un vettore e tipi di vettori
Il vettore più utilizzato è il plasmide, molecola circolare di DNA
capace di replicazione autonoma nella cellula batterica.
Si deve distinguere tra vettori utilizzati nella trasformazione con
•metodi fisici
•Agrobatterio
Raramente, si utilizzano anche vettori virali.
3
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
METODI DI TRASFERIMENTO GENICO
Figura 19.26
Confronto tra vettori virali e plasmidici riguardo alle loro modalità di
espressione.
4
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
METODI DI TRASFERIMENTO GENICO
Figura 19.27
Elementi costitutivi essenziali
di un genoma virale e vettore modello
per l’espressione transiente di un gene
esogeno in pianta.
5
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
METODI DI TRASFERIMENTO GENICO
Uno dei plasmidi più utilizzati per clonare in E. coli è pbr322:
dimensioni ridotte (4363 bp);
due marcatori: resistenza all'ampicillina (apr) ed a tetraciclina (tcr);
numerosi siti unici di restrizione;
integra frammenti di DNA da clonare lunghi fino a ca 10 kb.
Esistono varie categorie di vettori:
di clonaggio (pbr 322 di E. coli);
di trasformazione (pbr 322 + gene marcatore selezionabile);
di espressione (pbr 322 + gene marcatore selezionabile + sequenze per la
corretta espressione del gene da trasferire).
6
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
METODI DI TRASFERIMENTO GENICO MEDIATO DA VIRUS E AGROBATTERI
Caratteristiche di un vettore di espressione:
dimensioni ridotte per facilitarne l'isolamento e la sopravvivenza nella
cellula;
presenza di vari siti di restrizione unici per inserire le sequenze da clonare;
Capacità di integrarsi nel genoma dell'ospite;
presenza di marcatori genici in grado di conferire un vantaggio selettivo alle
cellule trasformate
presenza di promotori efficienti che garantiscano una attiva trascrizione del
DNA inserito.
7
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
METODI DI TRASFERIMENTO GENICO MEDIATO AGROBATTERI
Chi è Agrobacterum tumefaciens?
E’ un batterio gram negativo della famiglia delle
Rhizobiacee di forma bastoncellare. È
capace di infettare principalmente piante
dicotiledoni e causare una crescita
paragonabile a quella tumorale, una
patologia nota con il nome di galla del
colletto.
Come agisce Agrobacterum tumefaciens?
Possiede un plasmide, chiamato plasmide Ti
(Tumor Inducing) di circa 180 kb, che contiene un
segmento di DNA, definito T-DNA, che
penetra all'interno delle cellule vegetali
integrandosi nel loro genoma
e
ne
provoca
la
crescita
incontrollata.
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
METODI DI TRASFERIMENTO GENICO
MEDIATO AGROBATTERI
I plasmidi Ti sono costituiti da:
un frammento (T-DNA) che viene
integrato nel genoma della pianta
ospite;
la regione Vir che contiene
otto operoni
responsabili
della virulenza;
i geni per il catabolismo delle
opine;
sequenze
che
coniugazione permettono
tra
cellule
la
batteriche;
un’origine
di
replicazione
che permette di mantenere il
plasmide nel
corso
della
proliferazione
delle
cellule
9
batteriche.
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
METODI DI TRASFERIMENTO GENICO MEDIATO DA AGROBATTERI
Regione del T-DNA – regione di ca. 50 kb che viene inserita nel DNA
della pianta ospite e delimitata da sequenze terminali denominate “RB right border” e “LB - left border“.
Una volta inserita nell’ospite, la regione T provoca:
geni oncogeni con la
crescita
tumorale,
interferendo mediante
i
propri produzione di auxine e citochinine della
pianta.
sintesi delle opine che fungono da substrato metabolico per gli Agrobatteri e
stimolano anche la coniugazione del plasmide Ti .
10
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
METODI DI TRASFERIMENTO GENICO MEDIATO AGROBATTERI
Regione Vir
La regione (ca. 40 kb) contiene 8 operoni vir (A, B, C, D, E, F, G ed H), per lo più
policistronici (vari ORF), che regolano la sintesi delle proteine indispensabili per il
riconoscimento dell’ospite e il trasferimento del T-DNA alle sue cellule.
L’espressione dei geni vir avviene solo in presenza di cellule meccanicamente
danneggiate.
Il trasferimento del filamento -T è in parte analogo alla coniugazione batterica. Le
differenze consistono nel superamento della parete e membrana cellulare e del
citoplasma dell'ospite e nell'integrazione nel DNA nucleare.
11
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
METODI DI TRASFERIMENTO GENICO MEDIATO AGROBATTERI
Un meccanismo complesso…
1.Attivazione mediante segnale della pianta (composti fenolici prodotti in
seguito a ferita).
2.Induzione del complesso di virulenza
(geni Vir)
3.Generazione del pilo per il trasferimento del T-DNA
4.Formazione del T-DNA transfer complex.
5.Trasporto all’interno della cellula vegetale
6.Integrazione nel genoma vegetale
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
METODI DI TRASFERIMENTO GENICO MEDIATO DA AGROBATTERI
Come sfruttare Agrobacterum tumefaciens per la trasformazione genetica vegetale?
L'inattivazione dei geni oncogeni responsabili del tumore porta all'ottenimento di
plasmidi "disarmati" che risultano funzionali per il trasferimento ma non
causano tumore (vengono mantenute le sequenze di bordo e un marcatore di
selezione vegetale o un gene reporter).
La virulenza di un particolare ceppo di Agrobatterio dipende da eventuali
mutazioni nella regione vir, alcune delle quali hanno consentito di ottenere
ceppi ipervirulenti capaci di infettare anche cellule di Monocotiledoni.
Clonare all’interno del T-DNA disarmato la sequenza del gene di interesse che
verrà trasferito alla cellula vegetale. Si include un polylinker sintetico con vari siti
unici di restrizione in cui inserire uno o più geni da trasferire.
13
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
METODI DI TRASFERIMENTO GENICO MEDIATO AGROBATTERI
PROBLEMA
I plasmidi per clonare in E. coli non si replicano
in Agrobatterio e i plasmidi Ti non hanno
regione di replicazione per E.coli.
SOLUZIONE
VETTORI COINTEGRATI
Un unico plasmide contiene tutte
le funzioni necessarie per:
• Replicazione autonoma in E.coli e
Agrobacterium.
• Clonaggio della sequenza del
gene di interesse.
• Geni
per
il
nella
trasferimento
cellula vegetale.
VETTORI BINARI
•
•
•
Due plasmidi
separati:
Replicazione VETTORE
autonoma in E.coli e
Agrobacteium.
BINARIO
Clonaggio della sequenza del
gene di interesse.
PLASMIDE
Geni per il trasferimento nella
HELPER
cellula vegetale.
14
19.6MODIFICATE
PIANTE GENETICAMENTE
(PGM):
METODI
DI TRASFORMAZIONE GENETICA
METODI DI TRASFERIMENTO GENICO MEDIATO AGROBATTERI
Vettori binari (2 plasmidi separati):
Un PLASMIDE BINARIO di dimensioni ridotte (ca. 10 kb) con un "mini" T-DNA che
porta il gene da trasferire e capace di replicazione autonoma in diverse
specie
batteriche (es. E. coli e Agrobacterium).
Richiedono la presenza di un PLASMIDE HELPER,
presente
solo
nell’Agrobatterio,
contenente
la regione vir; il plasmide helper deriva da un
plasmide Ti "disarmato" e fornisce in trans le
funzioni di virulenza.
Nell’ Agrobatterio si
introducono quindi due
plasmidi,
entrambi
capaci di replicazione autonoma.
16
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
METODI DI TRASFERIMENTO GENICO MEDIATO E AGROBATTERI
Si esaminano prima le problematiche relative all’utilizzazione dell’Agrobatterio. Numerosi
fattori influiscono sull’efficienza globale di trasformazione:
La parte iniziale del processo (interazione Agrobatterio-pianta) è maggiormente
controllata dal patrimonio genetico dell’Agrobatterio.
La fase di integrazione della copia del T-DNA dipende dalle condizioni presenti nelle
cellule della pianta e dai fattori che regolano i processi di sintesi e di riparazione del DNA.
Nella fase successiva è importante la proliferazione e la differenziazione delle cellule
stabilmente trasformate. Necessità di sistemi in vitro caratterizzati da elevato tasso di
proliferazione e capacità morfogenica elevata e prolungata nel tempo.
In passato, la maggiore limitazione all'uso di A. tumefaciens è stata
la specificità per le Dicotiledoni. A partire dal 1995, l’uso di ceppi ipervirulenti ha
permesso di trasformare riso e mais utilizzando A. tumefaciens. Oggi quasi tutti i
cereali sono trasformabili con Agrobatterio.
18
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
METODI DI TRASFERIMENTO GENICO MEDIATO DA AGROBATTERI
L’infezione può essere realizzata immergendo gli espianti in una
sospensione di Agrobatterio oppure procurando lesioni all’espianto con
una lama previamente immersa nella sospensione batterica.
Nella fase di co-coltura, gli espianti vengono posti in substrato privo di
antibiotico su cui vengono lasciati per pochi giorni. Terminata la
fase di co-coltura si procede ad una decontaminazione degli espianti
dall’Agrobatterio con antibiotici opportuni.
Nella fase selettiva vera e propria, volta all’isolamento delle cellule che
esprimono il gene marcatore, al fine di limitare gli “escape” è
necessario mantenere l'agente selettivo nelle successive subcolture.
19
19.6MODIFICATE
PIANTE GENETICAMENTE
(PGM):
METODI
DI TRASFORMAZIONE GENETICA
METODI DI TRASFERIMENTO GENICO MEDIATO DA
AGROBATTERI
20
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
METODI DI TRASFERIMENTO GENICO CON METODI CHIMICO-FISICI
Come metodi alternativi all’ A. tumefaciens si sono sviluppati sistemi che non
utilizzano un vettore biologico e che quindi prescindono dalle interazioni tipiche
tra i componenti di un sistema biologico:
•Metodo biolistico
•Elettroporazione
•PEG/Lisosomi
•Microiniezione
•Trasformazione mediata dal cloroplasto
Ideati principalmente per specie recalcitranti alla trasformazione con
Agrobacterium
21
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
METODI DI TRASFERIMENTO GENICO CON METODI CHIMICO-FISICI
Metodo biolistico:
Particelle ricoperte con copie del gene
vengono accelerate ad
alta
velocità
per nelle cellule
consentire la loro penetrazione
bersaglio.
ASPETTI TECNICI: microproiettili di tungsteno, oro,
platino (1-4 µm di diametro) vengono accelerati a 400-550
m/s mediante l’uso di polvere da sparo, aria
compressa, scariche elettriche o gas compressi. Per
accelerare maggiormente le microparticelle le si fa
aderire ad un “macrocarrier” poi intercettato prima di
raggiungere il tessuto bersaglio.
Il successo del metodo biolistico
dipende dalla capacità di
delle microparticelle e dal danno
penetrazione
procurato alle cellule.
TESSUTI
BERSAGLIO:
callo
embriogenico
ed
organogenico, embrioni immaturi (cereali), polline,
meristemi, foglie, endosperma, sospensioni cellulari.
22
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
METODI CHIMICO-FISICI DI TRASFERIMENTO GENICO
Il metodo biolistico è particolarmente utile per determinare in tempi brevi (1-2
giorni) l'espressione transiente di geni anche se non integrati nel DNA genomico.
Il metodo biolistico
permette
quindi
saggi
di espressione transiente in
molti tipi di tessuto di specie diverse in tempi brevi.
Successivamente si verifica la stabilità dell’espressione a distanza di ca. 12 mesi dopo una fase selettiva: in questo caso, il livello di espressione
riscontrato, è maggiormente
predittivo dell’integrazione stabile del DNA nel
genoma vegetale.
23
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
INTEGRAZIONE ED ESPRESSIONE DEI TRANSGENI
Selezione e rigenerazione delle colture transgeniche
Data la bassa frequenza di trasformazione, è necessario utilizzare geni marcatori
selezionabili eucariotici per recuperare le cellule trasformate. A tal fine le
caratteristiche ideali del gene marcatore sono:
la facile identificazione delle cellule che lo esprimono
poche possibilità di escape delle cellule non trasformate all'agente selettivo a cui
il gene marcatore conferisce resistenza
nessun effetto negativo sulla capacità rigenerativa dei tessuti e sulla fertilità delle
piante rigenerate
i geni marcatori più utilizzati sono: epsp sintetatsi, fosfinotricina acetiltransferasi
(PAT, BAR), acetolattato sintetasi, neomicina fosfotranferasi II (NPTII).
25
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
INTEGRAZIONE ED ESPRESSIONE DEI TRANSGENI
Verifica dell'incorporazione del gene e dell’espressione nella pianta e
caratterizzazione delle progenie
Per essere utilizzabili in programmi di miglioramento genetico le piante
transgeniche devono esprimere regolarmente il gene inserito e, qualora si tratti di
specie a propagazione sessuata, mantenere inalterato il livello di espressione
nelle generazioni successive.
L’avvenuta trasformazione e la corretta espressione del transgene si dimostrano
a livello genetico, citogenetico, biochimico e fenotipico.
Segnali Southern in DNA non digerito ad alto peso molecolare, bande PCR, o
segnali da una singola linea putativamente trasformata non sono criteri accettabili
poichè è più difficile escludere la possibilità di artefatti in tali dati.
26
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
INTEGRAZIONE ED ESPRESSIONE DEI TRANSGENI
Figura 19.35
Sezione di tessuto fogliare
con cellule competenti, non
competenti
e potenzialmente competenti
alla rigenerazione e/o
alla trasformazione.
27
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
INTEGRAZIONE ED ESPRESSIONE DEI TRANSGENI
La sopravvivenza di linee cellulari su substrato selettivo non è sufficiente,
poichè vi è la possibilità di escape, di protezione da parte di prodotti secreti da
cellule microbiche contaminanti e/o la selezione di mutanti resistenti all'agente
selettivo. Saggi enzimatici su estratti cellulari sono inadeguati per la possibile
presenza di cellule microbiche contaminanti.
L’analisi del fenotipo fornisce indicazioni spesso imprecise a seguito della
tenue o mancata espressione e/o a seguito di silenziamento del gene inserito.
Dati fenotipici certi richiedono:
risultati negativi da controlli non trasformati;
un saggio che riveli il prodotto del transgene all'interno delle cellule
vegetali distintamente da possibili cellule microbiche contaminanti;
preferibilmente, il transgene non sarà espresso in cellule batteriche (GUS
con un introne);
Non si nota specificità di sequenza per i siti di inserimento nel genoma della
pianta.
Rispetto al metodo con Agrobatterio, il metodo biolistico integra un maggior
numero di copie nel genoma della pianta;
28
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
INTEGRAZIONE ED ESPRESSIONE DEI TRANSGENI
Importante verificare il destino del gene marcatore utilizzato per selezionare gli
eventi transgenici e del gene per cui non si applica pressione selettiva;
l’orientamento del gene marcatore e del gene da trasferire possono influire sul
grado di riarrangiamento e di espressione.
La struttura dei transgeni integrati risulta più complessa in assenza di pressione
selettiva.
La caratterizzazione dei materiali transgenici prevede la verifica di:
organizzazione molecolare delle sequenze inserite nel genoma della pianta;
livello di espressione dei geni inseriti;
trasmissione dei geni inseriti ed analisi dei rapporti di segregazione.
29
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
METODI DI TRASFERIMENTO GENICO
MEDIATO DA VIRUS E AGROBATTERI
Tabella 19.4
Elenco di sistemi di
geni marcatori
selezionabili
e di geni reporter
usati per la
trasformazione
genetica
delle piante.
30
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
INTEGRAZIONE ED ESPRESSIONE DEI TRANSGENI
Principali geni reporter utilizzati in Ingegneria Vegetale
Pianta di N. tabacum trasformata con il gene della Luciferasi
Pianta
di Arabidopsis
trasformata con il gene
GUS
Pianta di Arabidopsis
trasformata con il gene GFP
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
INTEGRAZIONE ED ESPRESSIONE DEI TRANSGENI
Figura 19.36
Segregazione 3:1 di un ipotetico marcatore PCR-derivato osservata
nella discendenza T1 ottenuta autofecondando una pianta T0
avente una singola copia del transgene.
32
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
INTEGRAZIONE ED ESPRESSIONE DEI TRANSGENI
Trasmissione del transgene e rapporti di segregazione
Verificare la trasmissibilità del transgene per via materna e per via paterna, poichè
particolari riarrangiamenti cromosomici sono trasmessi in misura diversa per via
femminile e per via maschile.
Verificare la trasmissibilità del transgene per autofecondazioni successive o
attraverso incrocio.
Trasmissione del transgene alle progenie
In un caso si è rilevata instabilità alla terza generazione dopo autofecondazione.
In alcuni casi si è riscontrata assenza di espressione per silenziamento del gene
sia a seguito di autofecondazione che di incrocio.
34
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
APPLICAZIONI DELLE VARIETÀ TRANSGENICHE
Applicazioni dell'ingegneria genetica
Obiettivi principali dell’ingegneria genetica applicata al miglioramento genetico delle
piante erbacee:
generare piante utili non ottenibili con il miglioramento convenzionale;
correggere difetti in cultivars in modo più efficiente rispetto al miglioramento
convenzionale;
L'entità del successo dipende dall'efficienza della trasformazione e dall’effetto
fenotipico e dalla stabilità di tale effetto in condizioni ambientali diverse e nelle
successive generazioni
Distinguere tra interventi per aggiungere una nuova funzione (es: resistenza a
insetti) da interventi per eliminare o ridurre una funzione indesiderata (es: inibizione
della PG o della biosintesi dell’etilene per ritardare la maturazione del frutto).
Specie erbacee di elevato interesse agronomico fino ad ora trasformate con
successo: mais, riso, frumento, orzo, cotone, soia, fagiolo, barbabietola,
girasole, fagiolo, tabacco, petunia, patata, pomodoro, melanzana, cassava.
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
Diffusione delle piante
transgeniche
Principali Produttori:
•Stati Uniti d’America (69 millioni di ha)
Mais, Soia, Cotone, Canola, Barbabietola, Erba medica
•Brasile (30.3 milioni di ha)
Soia, Mais, Cotone
•Argenitina (23.7 milioni di ha)
Soia, Mais, Cotone
•India (10.6 milioni di ha)
Cotone
•Canada (10.4 milioni di ha)
Canola, Mais
Crescita esponenziale della superficie
coltivata a colture transgeniche:
Dai 1.7 milioni di ha del 1996 ai 160
milioni di ha del 2011.
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
Diffusione delle piante
transgeniche
Negli ultimi anni
l’incremento delle
superfici adibite alla
coltivazioni transgeniche
è dovuto in particolare a
Paesi in via di Sviluppo
Un record di 170.3
milioni di ettari è stato
raggiunto nel 2012 con
un incremento rispetto
all’anno precedente del
6 %.
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
APPLICAZIONI DELLE VARIETÀ TRANSGENICHE
Considerazioni sull’utilizzazione dei prodotti vegetali transgenici
I rischi associati all’uso in campo ed al consumo di prodotti vegetali
transgenici vanno rigorosamente valutati e saggiati con attenzione di caso in caso.
Esistono Istituzioni e Commissioni di esperti in materia preposti alla definizione
delle regole e dei protocolli da seguire e delle prove tossicologiche necessarie
per determinare l’evetuale patogenicità di certi prodotti transgenici (es:
allergenicità).
I timori presenti al momento attuale nel pubblico sono del tutto ingiustificati
sulla base dei risultati sino ad ora ottenuti. Questo non comporta l’abbandono di
un atteggiamento di cautela.
E’ preoccupante il ruolo preponderante che il settore privato ha assunto in
questo settore, soprattutto per i grossi investimenti richiesti per la
commercializzazione delle colture GM.
Un problema normativo rilevante è costituito dalla brevettazione dei protocolli
sperimentali, dei geni clonati e delle piante transgeniche così ottenute.
Consensus Document (5/5/2004). Sicurezza alimentare e OGM.
http://www.siga.unina.it/circolari/Consensus_ITA.pdf
Consensus Document (15/3/2006). Cesistenza tra colture tradizionali,
biologiche e geneticamente modificate
http://www.salmone.org/wp-content/uploads/2007/09/consensus-coes.pdf
Documenti siglati da tutte le maggiori Società scientifiche italiane
Altri link di interesse per colture transgeniche
http://www.salmone.org/
www.AgBioWorld.org
Vedi anche: http://www.youtube.com/watch?v=QoXU6f-Arbo
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
APPLICAZIONI DELLE VARIETÀ TRANSGENICHE
Le principali applicazioni delle piante transgeniche sono:
modificare il processo di maturazione
resistenza ad erbicidi
resistenza a stress biotici (virus, batteri, funghi, nematodi, insetti)
resistenza a stress abiotici (freddo, salinità, eccesso idrico, pH, siccità)
modificare la composizione delle sostanze di riserva (carboidrati, lipidi, proteine)
modificare la biosintesi di metaboliti secondari (es: antociani, carotenoidi, ecc.)
modificare la fenologia (es: fitocromo, data di antesi)
modificare la ripartizione di fotoassimilati (es: rapporti sink-source)
produzione di anticorpi a fini terapeutici (es: immunoglobuline)
produzione di vaccini
produzione di enzimi ad uso industriale
(es: alfa-amilasi)
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
APPLICAZIONI DELLE VARIETÀ TRANSGENICHE
Resistenza ad erbicidi
Geni per resistenza ad erbicidi ad ampio spettro di azione e caratterizzati da
elevata attività specifica, bassa tossicità, rapida biodegradazione. Si sono
perseguite due strategie:
Principali specie trasformate: mais, cotone, soia, canola
Trasferimento di geni in grado di inattivare l'erbicida:
Glufosinate (fosfinotricina e bialaphos). E’ il principio attivo dell’erbicida Basta;
inibisce l’azione della GS (glutamina sintasi) e quindi altera tutto il metabolismo
azotato della cellula. Il gene bar isolato da Streptomyces hygroscopicussintetizza
la fosfinotricina acetiltransferasi (PAT) che detossifica il glufosinate.
Bromoxynil e’ il principio attivo dell’erbicida Brominalmais che viene utilizzato
anche in miscela; inibisce il trasporto degli elettroni nella fotosintesi e la
respirazione. Si è utilizzato il gene bxn da Klebsiella azaenae che produce un
enzima in grado di catabolizzare il bromoxinil.
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
APPLICAZIONI DELLE VARIETÀ TRANSGENICHE
Trasferimento di geni insensibili all'erbicida:
Glifosate. E’il principio attivo dell’erbicida Roundup; inibendo l’azione della EPSP sintasi, blocca, nel
cloroplasto, la biosintesi degli aminoacidi aromatici. Il gene EPSP sintasi insensibile al Glifosate è stato isolato
in Salmonella typhimurium.
E’ anche possibile ottenere resistenza attraverso la sovraespressione
dell’enzima della pianta mediante l’uso di promotori costitutivi forti.
Principali specie trasformate per resistenza ad erbicidi: mais, cotone, soia, canola
Più recentemente anche:
•Amaranthus palmeri trasformato con il gene della EPSP sintasi (Gaines et al., 2010)
•Patata dolce (Ipomoea batats) trasformata con gene bar per la resistenza a
Glufosinate (Zang et al., 2009)
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
APPLICAZIONI DELLE VARIETÀ TRANSGENICHE
Tabella 19.7
Alcuni esempi di erbicidi con diversa modalità di azione
e di enzimi codificati da geni clonati per lo più nei procarioti
ed impiegati per produrre piante transgeniche resistenti agli erbicidi.
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
APPLICAZIONI DELLE VARIETÀ TRANSGENICHE
Resistenza a stress biotici
Coat protein: Geni del capside virale che espressi nella pianta conferiscono resistenza a vari virus. Si
ritiene che la resistenza sia riconducibile ad un meccanismo di co-soppressione: la
pianta, riconosciuta la CP come proteina estranea, attiverebbe la sintesi di una molecola specifica in
grado di bloccare od attenuare l’espressione del gene virale deputato alla sintesi della CP.
Piante transgeniche che esprimono I geni CP di Tobacco Mosaic Virus (TMV) sono risultate resistenti a
TMV (Powell-Abel et al., 1986). In Cina è stata commercializzata la prima varietà di Tabacco
resistente a virus e da allora anche zucchino e melone.
Un altro metodo si basa sulla manipolazione delle proteine che permettono il passaggio del virus tra i
plasmodesmi che uniscono le cellule. Rispetto al precedente metodo, risulta a spettro d’azione più
ampio ed è più difficilmente superabile da parte del virus.
Espressione in tabacco del gene di lievito L3Δche conferisce in frumento resistenza alla tossicità da
Deossinvalenolo (DON) tossina del fungo Fusarium gramienarum (Di et al., 2010). La tossina DON
inibisce la traduzione mediante legame con la proteina L3 ribosomiale. La proteina L3Δ di lievito
risulta invece insensibile alla tossina.
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
APPLICAZIONI DELLE VARIETÀ TRANSGENICHE
Resistenza ad insetti
Uno dei più grandi successi della trasformazione genetica dei vegetali:
Il gene Bt di Bacillus thuringensis
La tossina Bt risulta efficace soprattutto contro Lepidotteri e in misura minore contro Ditteri e Coleotteri.
Innocua per Mammiferi
La proteina BT viene sintetizzata all’interno della pianta sotto forma di cristallo inattivo (cry-Bt). Se ingerita
dall’insetto, la proteina viene resa attiva in seguito a digestione intestinale. La tossina attiva si lega a
recettori delle cellule intestinali e determina la perdita incontrollata di ioni e morte cellulare per
lisi osmotica.
Il Bacillus thuringiensis è anche utilizzato direttamente irrorando le colture; il trattamento risulta
però alquanto oneroso e di scarsa efficacia se non ripetuto periodicamente. A seconda della composizione
amminoacidica, le proteine BT risultano più efficaci nei confronti di particolari ordini di insetti.
Cotone e mais Bt sono le colture che hanno riscosso il maggior successo.
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
APPLICAZIONI DELLE VARIETÀ TRANSGENICHE
Figura 19.44 Batteri
di Bacillus
thuringiensis (A) ed
esempi di piante
transgeniche
resistenti agli insetti:
danni da piralide in
spighe e culmi di
mais (B, C): varietà
di mais Bt (D); foglie
di una varietà di soia
Bt e di una normale
(E,F); capsule di una
varietà di cotone Bt
e di una normale
(G,H).
Mais BT resistente alla piralide e alla
diabrotica
(geni CryA da Bacillus thuringiensis)
wild-type
Mais GM per il
gene cry3Bb1
Danni alla radice
causati dalla
Diabrotica
Benefici derivanti dalla Gene Revolution
• Le biotecnologie contribuiscono ad una produzione
più sostenibile di prodotti vegetali tramite:
• Minor consumo di pesticidi
• Minor numero di lavorazioni del terreno
• Minore erosione del suolo
• Maggiore produttività
• Maggiore reddito per l’agricoltore
• Produzione più economica di biomasse
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
APPLICAZIONI DELLE VARIETÀ TRANSGENICHE
Resistenza ad insetti
Altri esempi
Alcune piante sono naturalmente equipaggiate con sistemi di difesa naturale.
Inibitori delle Proteasi: L’apparato digerente di molti insetti contiene tripsine e chimotripsine.
L’inibizione di queste proteine causa gravi danni alla loro crescita e sviluppo. Esempio del Riso:
trasformato con inibitori della proteasi della patata, di fagiolino, soia e orzo.
Inibitori della Alfa-amilasi: Inibiscono negli insetti il normale metabilismo dei carboidrati.
L’espressione di alfa-amilasi di segale (Secale cereale) in tabacco ha consentito di sviluppare piante
resistenti a Anthonomus grandis (curculionide patogeno di molte specie vegetali).
Lectine: Sono proteine di origine vegetale in grado di legare specifici carboidrati. Lectine da Bucaneve
(Galanthus nivalis) chiamate GNA sono state ampiamente utilizzate per introdurre resistenza a insetti in
specie coltivate (Riso e Patata). Determina 80% di mortalità senza alcun effetto sui mammiferi ed ha
effetto principale su omotteri e afidi.
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
APPLICAZIONI DELLE VARIETÀ TRANSGENICHE
Figura 19.45
Mortalità delle larve
del coleottero
Callosobruchus
maculatus
sulle piante di pisello
transgenico in
relazione alla
quantità
di inibitore dell’αamilasi.
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
APPLICAZIONI DELLE VARIETÀ TRANSGENICHE
Resistenza a stress abiotici
L’identificazione di geni responsabili della resistenza agli stress
abiotici è molto complesso:
• Controllo genetico quantitativo.
• Caratteri differenti sotto controllo degli stessi geni (effetti
pleiotropici).
• Differenti forme di stress abiotico interagiscono tra loro
attenuando o magnificando l’effetto dello stress.
• Condizioni ambientali difficili da controllare e spesso più
tipologie di stress agiscono contemporaneamente
Difficile identificare geni veramente utili per
trasformazione genetica
Possibili geni utilizzabili per introdurre resistenza a stress
abiotici:
• Percezione/Trasmissione del segnale cellulare
• Controllo della trascrizione
• Meccanismo di risposta allo stress
Vinocur and Altman, Current opinion in plant biology 16: 123-132
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
APPLICAZIONI DELLE VARIETÀ TRANSGENICHE
Resistenza a stress abiotici Alcuni esempi
Percezione/Trasmissione del segnale cellulare
Espressione costitutiva in Zea mays del gene di tabacco MAPKKK/NPK1 coinvolto nella trasmissione del segnale
(via della MAP chinasi) attiva la cascata del segnale che determina maggiore tolleranza a stress da freddo, caldo e salinità
(Shou et al., 2004).
Meccanismi di risposta allo stress
Heat shock Protein
Sovra-espressione del gene HSP101 di Arabidopsis in Oryza sativa determina una migliore performance durante la fase di
recupero dallo stress da caldo (Katyar-Agarwal et al., 2003). Sovra-espressione del gene HVA1 (proteina LEA isolata da
Hordeum vulgare) conferisce in piante di riso transgenico resistenza alla disidratazione cellulare (Chandra Babu et al., 2004).
Detossificazione da specie reattive dell’ossigeno
Sovra-espressione del gene dell’Aldeide deidrogenasi (AtALDH3) in Arabidopsis conferisce resistenza agli stress da siccità
e salinità (Sunkar et al., 2003). Le piante mostrano maggiore tolleranza alla disidratazione e ad altri fattori correlati
(sale, metalli pesanti, perossido di idrogeno).
Osmoprotettori
Buoni risultati ottenuti mediante trasformazione di Oryza sativa con gene della Colina deidrogenasi (codA) di
Arthrobacter globiformis; Il gene codifica per una proteina che catalizza l’ossidazione della colina in glicina-betaina
(noto osmoprotettore). Le piante transgeniche recuperano più facilmente in seguito a stress da eccesso di salinità e
producono maggiori quantità di seme rispetto al controllo (Mohanty et al., 2002).
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
APPLICAZIONI DELLE VARIETÀ TRANSGENICHE
Modificazione della biosintesi dei lipidi e del loro catabolismo
Si dovrà verificare quale sia il fattore limitante per la biosintesi. Gli acidi grassi
insaturi piùimportanti sono l'oleico (18:1), il linoleico (18:2) e il linolenico (18:3)
ottenuti per successive desaturazioni dallo stearico (18:0). Le desaturasi
responsabili delle prime due desaturazioni sono state clonate.
Le lipossigenasi rivestono un ruolo importante nel settore alimentare
(conservazione e proprietà organolettiche degli alimenti) e anche per la difesa
contro patogeni. Le lipossigenasi catalizzano la idroperossidazione degli acidi
grassi polinsaturi. I geni di varie lipossigenasi sono stati clonati.
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
APPLICAZIONI DELLE VARIETÀ TRANSGENICHE
Modificazione della biosintesi dei carboidrati
una alterata ADPUn gene
mutato di E. coli che codifica per
glucosopirofosforilasi trasferito in patata ha aumentato il livello
dell'amido, risultato importante da un punto di vista commerciale (il tubero ha
un contenuto basso di amido) e della qualità di certi prodotti (es: patate che
assorbono meno olio durante la frittura).
In riso e mais si puo’ manipolare la composizione in amido dell’endosperma. In
riso, si ha ca. il 15-30% di amilosio e il 70-85% di amilopectina. Da un mutante
incapace di produrre amilosio si è isolato il gene waxy.
Manipolando opportunamente questo gene ed inserendolo in altri cereali
si puo’ quindi alterare il rapporto tra amilosio ed amilopectina.
Sono stati isolati geni batterici per la produzione di carboidrati che
permettono l’ottenimento di polimeri particolari che, pur presentando
caratteristiche simili alla plastica, risultano più facilmente biodegradabili.
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
APPLICAZIONI DELLE VARIETÀ TRANSGENICHE
Modifica dei nutrienti
Il golden Rice
La carenza di Vitamina A può causare gravi danni danno all’apparato visivo e causa elevato grado di
mortalità infantile. A livello globale, ca. il 21 % dei bambini soffre di carenza da Vitamina A (Sommer, 2001).
Ca. 800.000 decessi di bambini e donne in gravidanza sono causate da deficienza nella dieta di Vitamina
A. Una possibilità è sovra esprimere I precursori della Vitamina in una coltura povera IL RISO.
L’endosperma del seme di riso manca di provitamina
A. E’ stato quindi prodotto riso transgenico contenete
4 geni isolati da Narcissus e Erwinia. Sono state ottenute linee stabili di riso che contengono grandi quantità di
Vitamina A e conferiscono al seme una colorazione dorata.
Una varietà di Golden Mustard
è
stata
recentemente
sviluppata e la tecnologia
sembra promettente per molte
altre colture..
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
APPLICAZIONI DELLE VARIETÀ TRANSGENICHE
Figura 19.48
Melanzana partenocarpica (foto: A. Spena).
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
APPLICAZIONI DELLE VARIETÀ TRANSGENICHE
Figura 19.50
Fiore wild-type (A) e del mutante
partenocarpico di pomodoro
pat-2 (B, notare l’accrescimento
precoce dell’ovario),
Bacche wild-type (C)
e del mutante pat-2 (D, notare
l’assenza di seme)
(foto: P. Mosconi).
OGM di seconda e terza generazione
• OGM con maggiore contenuto in:
• antiossidanti
• aminoacidi essenziali
• vitamine
• ferro
• omega-3
• micronutrienti
• OGM che producono:
• un diverso profilo di acidi grassi
• anticorpi per uso diagnostico o terapeutico
(es. per antigeni tumorali)
• antigeni (es. per la vaccinazione)
• proteine ricombinanti di interesse terapeutico
(es. emoglobina, lattoferrina, insulina)
High provitamin A corn; 100-200 g of grain
provide full RDI of β -carotene (as a sole
source of vitamin A)
Courtesy of
Paul Christou
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
APPLICAZIONI DELLE VARIETÀ TRANSGENICHE
Molecular pharming
Produzione di Vaccini/Anticorpi:
Problema
Per la produzione di Vaccini su larga scala spesso le cellule di mammifero sono difficili da gestire e si
ottengono basse quantità mentre le cellule batteriche spesso non sintetizzano la proteina nella
sua corretta conformazione nativa.
Quindi ?
PRODUZIONE DI ANTICORPI
In pianta è possibile produrre vaccini o anticorpi
nella forma nativa in grandi quantità. E’ anche
possibile alimentare direttamente animali e/o
uomo con la pianta modificata
PRODUZIONE DI ANTIGENI
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
APPLICAZIONI DELLE VARIETÀ TRANSGENICHE
Produzione di Vaccini: Alcuni esempi di utilizzo del transgenico per la produzione di vaccini in pianta
Antigeni Batterici
Enterotoxigenic E. coli Heat-labile toxin B subunit (LTB)
Carrots
Immunogenic and protective against CT challenge
Cholera toxin B subunit (CTB)
Soybean
Tomato
Rice
Immunogenic and partial protection against LT challenge in mice
Immunogenic by oral delivery to mice
Immunogenic and protective against CT challenge to mice following oral
administration
Hepatistis B virus surface antigen
Potato
Immunogenic response in humans following oral administration
Hepatitis B virus surface antigen fused with preS1 epitop
Rice
Immunogenic by intraperitoneal delivery to mice
Human group A rotavirus (VP6) protein
Alflfa
Immunogenic in mice and offspring developed less severe diarrhea after
challenge with simian rotavirus SA-11, indicating that antibodies generated in
the dams provided passive heterotypic protection to the pups
Rotavirus (VP7)
Potato tubers
Antigeni Virali
Norwalk virus capsid protein
SARS-CoV S protein (S1)
Smallpox recombinant vaccine virus B5 antigenic domain
(pB5)
Japanese encephalitis virus (JEV) envelope protein (E)
Immunogenic in mice following oral delivery. Neutralization activity against
rotavirus
Tomato Fruit and
Elicit systemic and mucosal antibody responses in mice following oral
Potato
administration
Tomato and tobacco Immunogenic to mice following oral administration
leaf
Tobacco and Collard Antibody response in mice immunized parenterally and protects against lethal
leaf
dose of vaccinia virus
Rice
JEV specific neutralizing antibody detected in mice following intraperitoneal
or oral administration
Daniell et al., Trends Plant Sci. 2009 December; 14(12): 669–679.
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
APPLICAZIONI DELLE VARIETÀ TRANSGENICHE
Produzione di Vaccini: Alcuni esempi di utilizzo del transgenico per la produzione di vaccini in pianta
Antigeni Batterici
Plasmodium yoelii merozoite surface protein (PyMSP4/5) Tobacco
Induces antigen-specific antibodies in mice following parenteral
delivery
Human vaccines (Viral/Transient expression)
Bacterial antigens
Yersinia pestis F1 and LcrV antigens
Tobacco leaf tissue Immunogenic and protective in monkeys against Y. pestis
following subcutaneous injection
Yersinia pestis F1-V antigens
Tobacco leaf
Immunogenic and protection in vaccinated guinea pigs against
Y. pestis aerosol challenge
Smallpox recombinant vaccine virus B5 antigenic domain
(pB5)
Tobacco leaf
Antibody response in mice immunized perenterally and
protective against lethal dose of vaccinia virus
Encoding domain III of the dengue 2 envelope protein
(D2EIII)
Tobacco
Retains antigenicity and immunogenicity as well as inducing
neutralizing antibodies in vaccinated animals.
HIV entry inhibitors red algal protein griffithsin (GRFT)
Tobacco (TMV)
Active against HIV at picomolar concentrations, directly virucidal
via binding to HIV envelope glycoproteins and capable of
blocking cell-to-cell HIV transmission
Pathogenic avian influenza virus (H5N1 subtype)
Tobacco
Immunogenic in mice and ferret and also protects ferrets
against challenge infection with virus
Antigeni Virali
Daniell et al., Trends Plant Sci. 2009 December; 14(12): 669–679.
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
APPLICAZIONI DELLE VARIETÀ TRANSGENICHE
Produzione di Anticorpi
La sintesi di anticorpi in organismi ricombinanti richiede la sintesi di
due proteine e quindi l’espressione di due geni. Il metodo risulta efficace
solo per proteine di peso molecolare inferiore a 20.000 e che possono
uscire dalla parete cellulare. Per le sue rilevanti dimensioni, l’anticorpo non
puo’ uscire dalla cellula.
Differenti tipologie di proteine anticorpali
prodotte in pianta
Produzione su larga scala di anticorpi per uso terapeutico e diagnostico.
L’iniezione di anticorpi diretti ad antigeni di superficie è una delle
possibili strategie per la lotta alle cellule cancerogene; in questo caso sono
richieste quantità elevate di anticorpo.
Pianta
Tipologia di Ab (target)
Obiettivo
Referenza
Tobacco
IgG (nematode)
Plant pathogen resistance
Baum et al. (1996)
Tobacco
sIgA/G (Streptococcus mutans)
Therapeutic (topical)
Ma et al. (1998)
Soybean, rice
IgG (herpes simplex virus)
Therapeutic (topical)
Zeitlin et al. (1998)
Tobacco
IgG (colon cancer)
Therapeutic (systemic injection)
Verch et al. (1998)
Alfalfa
IgG (human IgG)
Diagnostic
Khoudi et al. (1999)
Tobacco
IgG (rabies virus)
Therapeutic: first IgG expressed in plant
showing therapeutic activity (systemic
injection)
Ko et al. (2003)
Tobacco
IgG (hepatitis B virus)
Immunopurification of hepatitis B
surface antigen
Valdes et al. (2003)
Tobacco
IgG (hepatitis B virus)
Therapeutic
Yano et al. (2004)
Ko et al.,Virus research. 2005 111: 93-100.
PRODUZIONE DI BIOFARMACI, ANTICORPI E VACCINI
MEDIANTE PIANTE GENETICAMENTE MODIFICATE
PRODUZIONE DI ANTICORPI (IMMUNOGLOBULINE) NELLE PIANTE
Figura 19.53
Costi per grammo
di immunoglobulina A
prodotta con diversi
sistemi di espressione.
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
INTEGRAZIONE ED ESPRESSIONE DEI TRANSGENI
RNA Interference
Mediante questa tecnica si cerca di ridurre notevolmente l’espressione di un
gene la cui normale espressione risulta negativa ai fini produttivi e/o di
conservazione del prodotto.
Effetto scoperto per la prima volta in petunia
64
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
Ha avuto nella storia differenti nomi (Co-soppressione; RNA interference; RNA
antisenso; Post-trascriptional-gene-silencing) ma il meccanismo di base è simile:
Formazione di una molecola a doppio filamento tra sequenze complementari di
RNA "antisenso" e di mRNA "senso" del gene di cui si vuole sopprimere l'azione.
Si può inserire una sola porzione del gene da silenziare o il gene per intero.
L'RNA antisenso può interferire con la trascrizione, lo splicing, la traduzione del
mRNA senso o rendendo quest'ultimo, a seguito della loro unione, più sensibile alla
degradazione; quest’ultima sembra essere l'ipotesi più probabile.
65
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
La riduzione del
trascritto originale
sembra avvenire a
diversi livelli
Modificazione della cromatina
a livello istonico e inibizione
della trascrizione
Legame con mRNA e creazione
di un complesso non accessibile
per la traduzione.
Legame con mRNA e
degradazione a carico
della Dicer protein.
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
siRNA
Small interfering RNA (21-24 nt). Originano da acidi nucleici aberranti
o patogeni e agiscono in cis inattivando la stessa molecola da cui sono
stati prodotti (meccanismo di difesa della pianta).
Virus-Induced-Gene-Silencing
VIGS
La tecnologia sfrutta il sistema di difesa RNA-mediato che
la pianta attiva contro virus patogeni.
Vettori virali trasformati con geni dell’ospite quando inseriti
in quest’ultimo determinano una riduzione transiente
dell’espressione dei geni stessi.
Esempio in H. vulgare (BSMV mediated silencing):
•Genoma tripartitico (α,β,γ). Il genoma β è disarmato per
impedire la formazione del capside e nel genoma γ è
inserito il gene da silenziare (GOI)
•I tre genomi vengono mescolati in rapporto (1:1:1)
•Inoculati direttamente in foglia o all’apice della pianta fino
alla formazione dei sintomi del mosaico
•Verifica del silenziamento genico dopo poche settimane.
Int J Plant Genomics. 2009. Published online 2009 June 15
VANTAGGI:
Semplice, economico, rapido e in grado di superare ridondanza funzionale.
SVANTAGGI:
Il gene non è mai completamente silenziato, alterazioni fenotipiche dovute al genoma ospite, offtargets, espressione transiente
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
APPLICAZIONI DELLE VARIETÀ TRANSGENICHE
B
Figura 19.46
(A)Strategia antisenso: la trasformazione
di piante con geni aventi un orientamento
inverso rispetto alle regioni di regolazione
determina il blocco della espressione del
corrispondente gene endogeno.
(B)Pomodori transgenici e prodotto
commerciale americano a base di
pomodori transgenici.
TRASFORMAZIONE GENETICA COME STRUMENTO
PER LO STUDIO DELLA FUNZIONE GENICA
miRNA
Micro RNA (21-22 nt). Originano da geni regolatori endogeni e agiscono in
trans silenziando geni specifici con sequenze omologhe (sistema
di regolazione della pianta).
artificial micro RNA: amiRNA
Trasformazione con miRNA artificiale progettati a
partire da un pre-miRNA endogeno in cui la
regione
che determina la specificità viene
sostituita con un porzione del gene target.
Il gene viene trascritto da RNA pol II, tagliato
dall’enzima Dicer-like1 e indirizzato mediante il
complesso AGO/RISC al gene target per il
silenziamento in maniera altamente specifica
VANTAGGI:
Alta specificità e assenza di off-targets, espressione più stabile rispetto a VIGS,
possibilità di progettare costrutti con diversi target, non genera effetti secondari dovuti al
genoma virale.
SVANTAGGI:
Il gene non è mai completamente silenziato, più complesso e laborioso rispetto a VIGS.
PIANTE GENETICAMENTE MODIFICATE (PGM):
METODI DI TRASFORMAZIONE GENETICA
INTEGRAZIONE ED ESPRESSIONE DEI TRANSGENI
Un settore di elevato interesse è la regolazione della maturazione dei frutti in
cui si abbia degradazione enzimatica delle pareti cellulari a seguito dell'azione
della poligalatturonidasi (gene pg). La pg è sintetizzata durante la maturazione.
In pomodoro, un frammento 5' di 730 bp del clone pg cDNA è stato clonato in
senso inverso sotto controllo di CaMV35S. In una pianta così trasformata, il livello
di pg risultava ridotto al 6%. L'espressione del gene antisenso non ha
influenzato l'espressione di altri geni coinvolti nel processo di maturazione.
La tecnica dell'antisenso ha permesso di:
saggiare la funzione di specifici geni
identificare geni a funzione sconosciuta
ottenere piante con nuove caratteristiche
•
•
•
•
ridotta espressione della subunità piccola della rubisco
ridotta espressione di una proteina a 10 kda coinvolta nel fotosistema ii
modifica del colore in fiori di petunia e gerbera
69
modifica del processo di maturazione in pomodoro e melone.
PIANTE GENETICAMENTE MODIFICATE (PGM):
GENE TARGETING
Tecnologie che consentono di introdurre alterazioni mirate a specifiche sequenze
all’interno del genoma ospite senza alterarne il background genetico
Sfruttano i normali meccanismi di riparazione delle
cellule eucariote dei tagli a doppio filamento nel DNA:
•Homologous Recombination (HR): Il gene compromesso viene sostituito per ricombinazione
con una sequenza omologa non-danneggiata.
NHEJ
In seguito al taglio il gene danneggiato viene sostituito
con un gene di nostra progettazione: possiamo
introdurre qualunque tipo di modifica.
HR
HR + NHEJ
N.B.
dobbiamo
fornire
mediante
trasformazione genetica il gene da noi progettato
(Repair Target) per la sostituzione mediante HR
•
Non-Homologous-End-Joining
(NHEJ):
Le
due
estremità
del
gene
danneggiato vengono semplicemente unite
con perdita o inserimento di alcune coppie di
basi.
In seguito al taglio le due estremità vengono ricongiunte
e si creano mutazioni di frameshift quindi solo mutazioni
knock-out.
Problemi irrisolti
• Scarsi gli investimenti a livello nazionale
• Predominio del settore privato
• Scarsa accettazione dei prodotti GM
• Disinformazione su effettivi rischi e
benefici dei prodotti GM
• Eccesso di regolamentazione (OGM)
• Carenza di una comunicazione appropriata
Conclusioni
• La genomica e l’ingegneria genetica offrono molteplici opportunità
per migliorare la resa e sostenibilità della produzione agricola e
migliorare il profilo nutrizionale e la salubrità degli alimenti.
• Per gli OGM, solo una corretta informazione ed una politica meno
restrittiva potranno incentivare adeguatamente la ricerca e le sue
applicazioni; l’accettazione pubblica svolgerà un ruolo fondamentale.
• Il persistere di una ingiustificata penalizzazione verso le colture GM
e le limitazioni alla ricerca contribuiscono ad aumentare
ulteriormente il divario tecnologico dell’Italia nei confronti di molte
altre nazioni.
• Le biotecnologie delle piante agrarie avranno un ruolo di crescente
rilevanza per la competitività e la sostenibilità della filiera agroalimentare, per la produzione di biomasse e la salubrità delle derrate
alimentari.