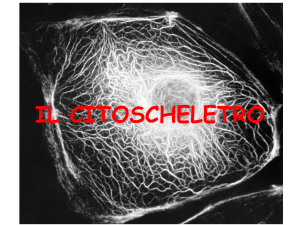
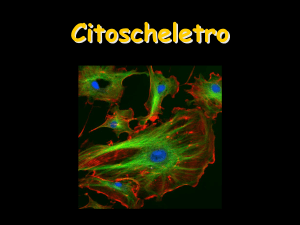
Motilità e contrattilità
cellulare
Il movimento può avvenire a livello:
-Subcellulare→ es cromosomi in mitosi; correnti citoplasma,trasporto vescicole
-Cellulare →protozoi ciliati, spermatozoi, migrazioni morfogenetiche, metastasi
- di tessuto e/o organismo pluricellulare →muscolo scheletrico, cuore
L’energia per il movimento proviene dall’ idrolisi di ATP che determina cambi
conformazionali in speciali proteine motrici
Negli eucarioti il movimento dipende dall’interazione di questi motori molecolari
con un’impalcatura di base formata da:
-MICROTUBULI o
-MICROFILAMENTI
•Movimento basato su microtubuli: interazione tra microtubuli e le MAP motrici:
dineine citoplasmatiche e assonemali, chinesine (movimenti intracellulari e di
appendici esterne).
•Movimento basato su microfilamenti: interazione tra microfilamenti di actina e le
miosine (contrazione muscolare e movimenti non muscolari)
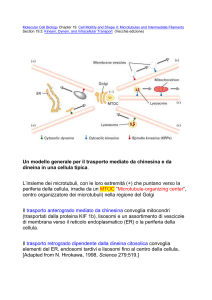
Motilità basata su microtubuli: movimenti intracellulari
I microtubuli sono le vie preferenziali su cui viaggiano vescicole
chinesina e dineina sono MAP motrici responsabili del moto grazie a idrolisi di
ATP;
•Chinesina→trasporto anterogrado (vs polo +, lontano dal MTCO)
•Dineina →trasporto retrogrado (vs polo – e MTCO)
Chinesine
Le chinesine hanno tre domini:
1)testa globulare che lega microtubulo e
idrolizza ATP
2) regione ad elica
3) regione di interazione con altre proteine e
organelli
Le teste globulari funzionano come “piedi”
per camminare lungo microtubulo (vs
estremità +)
Dineine citoplasmatiche
Formate da associazione di diverse
catene; le catene pesanti formano le
teste globulari che interagiscono con il
microtubulo
Si muovono vs estremità (-)
Traffico vescicolare
Le vescicole sono trasportate dalle MAP motrici su vie costituite da
microtubuliàdefinisce la polarità cellulare, struttura e distribuzione del RE,
Golgi, lisosomi, vescicole secrezione
Trasporto assonale
Le proteine motrici trasportano organelli e vescicole dal corpo cellulare verso
estremità e viceversa sfruttando i microtubuli come “piste”
Motilità basata su microtubuli
1. Movimento intracellulare
2. Movimento di appendici mobili (ciglia e flagelli)
In organismi unicell. moto e
raccolta cibo→ es protozoi ciliati
tipo Paramecium
In org. multicell. movimento
ambiente circostante o cell.
sessuali specializzate → es
epitelio vie respiratorie per
raccolta muco-polvere
Ciglia, in genere numerose,
“battito a remo”
Flagelli, più lunghi e radi,
movimento ondulatorio (da forza
propulsiva in ambiente fluido)
la struttura portante di ciglia e flagelli è un assonema (sistema cilindrico di
microtubuli di 0.25μm diametro) rivestito di membrana plasmatica e connesso al
corpo basale, che funziona come MTCO
assonema
Corpo basale= 9 triplette di mt disposte
a cerchio (mt completi)
ha la stessa struttura di centriolo (=9
triplette in cerchio);
funziona da centro di nucleazione per l’
assonema sovrastante
z. transizione
Assonema= 9+2
9 coppie di mt periferiche e 1 centrale
Nelle coppie periferiche un mt (A)
completo ed uno addossato incompleto
(B)
corpo basale
Zona transizione ha organizzazione
intermedia tra assonema e corpo basale
Da ciascun tubulo A delle doppiette esterne partono 2 bracci laterali (interno ed est) di
dineina assonemale, che contatta il tubulo B della doppietta accanto; i bracci sono disposti
a intervalli regolari nel senso longitudinale.
Nexina nei legami interdoppietta; bracci radiali collegano le doppiette alla coppia centrale
La forza meccanica per scorrimento dei mt data da idrolisi di ATP e cambi
conformazionali della dineina che fa scivolare 2 doppiette periferiche una
sull’altra: lo scorrimento diventa flessione locale dell’assonema
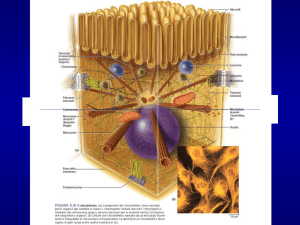
Movimento basato sull’interazione tra actine e miosine
Le miosine coinvolte in processi diversi come la
contrazione muscolare, moto cell., fagocitosi,
strutture udititive, trasporto melanina, etc.
Super-famiglia delle miosine
Tutte hanno almeno una catena pesante
organizzata in zona globulare (testa) che lega
actina e idrolizza ATP e regione fibrosa variabile
(coda) per interazione con altre molecole
Miosine tipo II, più note, nel muscolo liscio e
scheletrico, miocardio e cell non muscolari
Nei sarcomeri c’è una disposizione ordinata di actina, miosina e
proteine accessorie
a)
a) Il sarcomero è l’unità contrattile
b)
La contrazione è spiegata dal modello dello scorrimento dei filamenti :
I filamenti spessi “camminano” su quelli sottili trainadoli vs centro del
sarcomero→ si contrae sarcomero e banda I;
i filamenti non si accorciano, aumenta invece la loro sovrapposizione!
La contrazione è regolata dalla [Ca2+]
La [Ca2+] regola la disponibiltà dei siti di legame per la miosina
sull’actina
L’interazione actina-miosina è coinvolta non solo nella contrazione muscolare ma anche in
molti tipi di movimento non muscolare: es citocinesi, movimento cellulare strisciante
(ameboide e filopodi), chemiotassi, correnti citoplasmatiche
Movimento strisciante mediante emissione di speciali prolungamenti
citoplasmatici : lamellipodi e/o filipodi
Il movimento strisciante mediato da filopodi è tipico dei fibroblasti e delle
migrazioni cellulari embrionali
I filopodi emergono evidenti dalla superficie cellulare, dentro I filopodi si
trovano fasci paralleli di actina
Le miosine “tirano dietro” i
microfilamenti di actina che man
mano si allungano all’estremità (+)
per polimerizzazione.
In seguito ad adesione salda al
substrato, mediata da proteine
integrine, la polimerizzazione dei mf
comporta un allungamento netto
con spostamento in avanti della
protusione
Il movimento ameboide
Il movimento strisciante di tipo ameboide è tipico di alcuni protozoi, funghi e
leucociti (funzione di motilità, nutrimento-fagocitosi, difesa)
Si basa su:
-emissione di protusioni citoplasmatiche dette pseudopodi
-alternanza di transizioni gel-fluido nello stato del citoplasma
Correnti fluide si muovono vs pseudopodio e solidificano sulla punta mentre, al
margine posteriore, l’ectoplasma fluidifica e scorre vs protusione