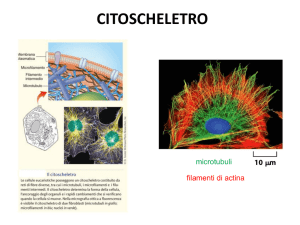
Localizzazione nella cellula
LE TRE CLASSI DI
ELEMENTI
CITOSCHELETRICI:
MICROTUBULI
MICROFILAMENTI
FILAMENTI INTERMEDI
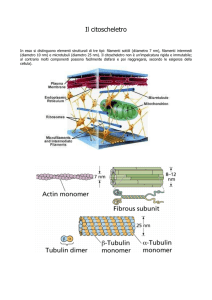
Struttura dei microtubuli
13 protofilamenti ciascuno formato da molecole
globulari di tubulina alfa e beta
Microscopia elettronica di un microtubulo
In sezione longitudinale
Anche in vitro le molecole di tubulina formano
spontaneamente microtubuli polarizzati
I microtubuli si allungano o si
accorciano per aggiunta o
sottrazione di dimeri di tubulina
rispettivamente all’ estremità
positiva rivolta verso la membrana
plasmatica e all’estremità
negativa rivolta verso il centrosoma
I microtubuli sono strutture labili
che possono facilmente polimerizzare
(formazione del fuso mitotico) e
depolimerizzare (per es. quando
la cellula entra in mitosi)
TUBULINA:
L’unità di
base è un
dimero
Le subunità alfa e beta si trovano sempre associate a formare un
eterodimero di tubulina. Entrambe legano una molecola di GTP ma solo
quella legata a beta è Idrolizzabile a GDP. All’interno del microtubulo il
dimero è alfa-tubulina/GTP-GDP/beta-tubulina.
In vitro la polimerizzazione avviene in presenza di GTP, Mg, EGTA e MAPs.
Nella cellula i microtubuli in genere
originano da centri organizzatori
IL CENTROSOMA, CENTRO DI
ORGANIZZAZIONE DEI MICROTUBULI, PUÒ
CONTENERE UNA COPPIA DI CENTRIOLI.
I microtubuli citoplasmatici (a differenza di quelli assonemici) non hanno un
rapporto diretto con i centrioli, ma nascono dal materiale pericentriolare che
contiene anelli di tubulina γ e che appare essere il
centro di organizzazione dei microtubuli, MTOC.
Nella cellula in interfase è di solito presente una coppia di centrioli posti
ad angolo retto tra di loro che occupano il centro geometrico dalla cellula.
In preparazione della mitosi la coppia di centrioli duplica
I centrioli possono prendere origine dai corpi basali e viceversa
Microtubuli e proteine
motrici determinano il
traffico di vescicole
nella cellula
Ai microtubuli sono associate in modo specifico altre proteine chiamate MAP
Alcune MAP sono le proteine tau in grado di stabilizzare la struttura microtubulare
Altre MAP sono le chinesine e dineine citoplasmatiche. Si tratta di motori microtubulari
(enzimi meccanochimici) che convertono l’enegia dell’ATP in movimento di
scorrimento lungo i microtubuli.
PROTEINE MOTRICI
CAMMINANO LUNGO
I TUBULI,
IDROLIZZANDO ATP
ACTINA:
L’unità di
base è un
monomero
Due filamenti a doppia elica
La presenza di un capo del filamento caratterizzato da actina legata ad ATP
identifica l'estremità "più", mentre l'altro capo, dove l'actina è legata a ADP
costituisce l'estremità "meno" del filamento. L'idrolisi dell'ATP in ADP riduce
la forza del legame tra le subunità, favorendo dunque la depolimerizzazione
all'estremità "meno".
I microfilamenti di actina hanno un ruolo
essenziale nel movimento cellulare.
Esistono molti tipi diversi o isoforme di actina
codificate da geni diversi:
-actina delle cellule muscolari scheletriche
-actina delle fibre muscolari cardiache
-actina delle cellule muscolari liscie dei vasi
-actina delle altre cellule muscolari liscie
-due isoforme di actina delle cellule non muscolari
Altre strutture importanti collegate a filamenti di actina:
Si possono identificare due tipi di strutture adesive cellula-matrice,
podosomi ed adesioni focali. Entrambi sono collegati ai filamenti di
actina e alle integrine della membrana plasmatica che a loro volta si
legano alle proteine della matrice extracellulare
Adesioni focali
Filamenti di actina
chiamati anche
Fibre da stress
I microfilamenti hanno sia ruoli strutturali che di guida
per i movimenti degli organelli
Nelle cellule secernenti consentono lo spostamento delle vescicole secretorie
nei pressi della membrana plasmatica, nell’atto finale della secrezione.
Filamenti di actina e molecole di miosina spostano le vescicole verso la membrana
Funzioni dei filamenti di actina
forniscono sostegno e forma alla cellula, collegandosi con
gli organelli interni e con la membrana plasmatica;
permettono una parte del trasporto interno di vescicole;
i sarcomeri, le unità contrattili delle cellule muscolari sono
formati da actina, oltre che da miosina ed altre proteine;
costituiscono la base su cui si formano le principali
estroflessioni cellulari, quali microvilli, stereociglia, pseudopodi;
sono coinvolti nell'adesione tra cellule e tra cellula e un
substrato.
ACTINA
MIOSINA
MIOSINA
La miosina possiede un notevole grado di eterogeneità e nelle cellule
non muscolari si assembla “su domanda” in piccoli filamenti.
Nello spostamento delle vescicole secretorie interviene una miosina
molecolare (miosina 1) che si lega contemporaneamente alla superficie
della vescicola ed ai filamenti di actina
FIBRA MUSCOLARE LISCIA
Fibra muscolare scheletrica
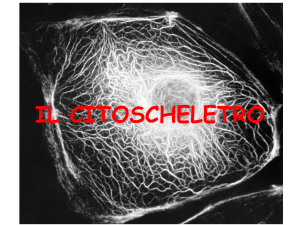
Cellula nervosa colorata in modo da evidenziare il citoscheletro.
E’evidente la abbondanza di strutture citoscheletriche formate da
microtubuli e filamenti intermedi
Tutti i filamenti intermedi sono molto stabili!
Sono formati da subunità proteiche lineari ad alfa elica che in vitro si
autoassemblano spontaneamente in filamenti senza richiedere fattori
catalitici, proteine accessorie o apporti energetici esogeni.