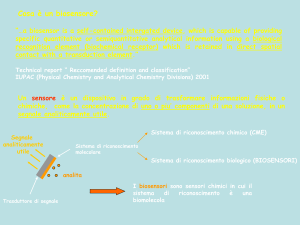
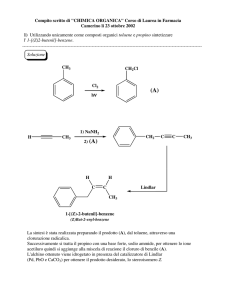
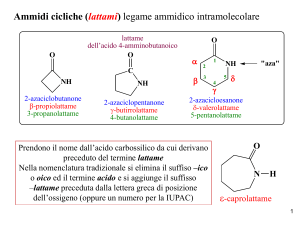
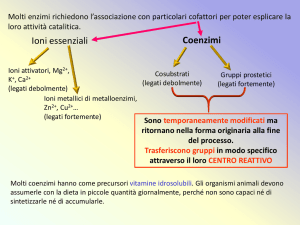
Transferasi che hanno come coenzima
FOSFOPANTOTEINA E COENZIMA A
(derivati dalla VITAMINA B3 )
Altre transferasi hanno come CoE:
Vitamina B3 (sintetizzata in vegetali e molti procarioti
anche ospiti dell’intestino crasso di molti vertebrati).
Queste cellule usano come base di partenza la valina e
l’acido aspartico.
O
HO
CH3 O
OH
O
NH2
Acido aspartico
Prof. G. Gilardi - Biological Chemistry
H3 C
OH
NH2
valina
1
Sintesi della Vit. B3
La valina è sottoposta ad un processo di transdesaminazione si ha
uscita di NH3 e sostituzione con OH diventa α-chetoacido.
CH3 O
CH3 O
H3C
CH3 O
OH
NH2
H3 C
OH
+ NH2-enzima
H3 C
NH
+
OH
O
enzima
Valina
ac. α−cheto β-metil butirrico
base di Schiff
Un secondo enzima riconosce questo substrato, ha nel sito attivo un residuo di
lisina che può dare attacco nucleofilo sul C=O dell’α-chetoacido. A reazione
ultimata si ottiene il substrato legato all’E con liberazione virtuale di H2O. Il
composto è una Base di Schiff. Altri R aa presenti nel sito attivo protonano l’N.
Prof. G. Gilardi - Biological Chemistry
2
Sintesi della Vit. B3
Si scatena così la trasposizione del doppio legame nel substrato con
dissociazione dal Cβ di 1 H + .
Il ritorno alla forma di risonanza precedente (eq.tra le forme di risonanza)
rende il Cβ carbanionico ed in grado di accettare certi gruppi (uno di
questi è l’idrossimetile + CH2OH)
CH3
C
H C CH3
CH3
+
N
H
enzima
CH
COOH
3
COOH
CH3
C
C
C
H+
N
H
enzima
C
CH3
+
N
H
enzima
CH3
CH3
CH3
+ CH2OH
Prof. G. Gilardi - Biological Chemistry
3
Sintesi della vit B3
Poi avremo deprotonazione dell’N della base di Schiff.
Idrolisi del doppio legame alla fine il prodotto sarà il derivato precedente che si è
accresciuto del gruppo –CH2OH. Il Cα è ridotto a gruppo alcolico secondario da una
oxred. NAD dipendente).
PRODOTTO FINALE che esce dal sito attivo dell’enzima: α,γ diossi- βdimetil butirrico o
ACIDO PANTOICO
COOH
C O
H3C
C
H2C
CH3
OH
COOH
C OH
H
C
H3C
H2C
CH3
OH
ACIDO PANTOICO
Prof. G. Gilardi - Biological Chemistry
4
Sintesi della Vit. B3
2) L’acido aspartico, attraverso un E liasico viene solo decarbossilato sulla
posizione 1. Questa decarbossilazione lo trasforma in una struttura con un
C in meno, è un aa ma non α-aa: è un β-aa
β ALANINA.
O
HO
CH2
CH2
O
HO
C
CH2
CH2
OH
NH2
CH2
O
ac.aspartico
CH3
NH2
β-alanina
Prof. G. Gilardi - Biological Chemistry
5
Sintesi della Vit. B3
SINTESI vit B3.
Ultima tappa: unione di acido pantoico e β-alanina.
La reazione è promossa da un Manganese-Enzima.
Il Mn lega con legame dativo l’O del carbossile poi il legame da dativo a
covalente (il Mn aumenta il suo numero di covalenza).
Si ha una sottrazione di OH ed il carbocatione che resta subisce l’attacco
nucleofilo dell’NH2 della β-alanina.
Mn- Enzima
vitamina B3.
+
H
H3C
acido pantoico
COOH
C OH
C
H2C
CH3
OH
H
H3C
COOH
C OH
C
H2C
Prof. G. Gilardi - Biological Chemistry
CH3
OH
H2N CH2
CH2
+
COOH
β-ALANINA
6
Vitamina B3: acido pantotenico
OH
H 3C
HO
C
CH 2
C
NH
CH 2 O
CH 2 C
CH3 O
OH
AC. PANTOTENICO o vit B3
E’ la molecola base con cui tutte le cellule costruiscono i CoE per le
transferasi B3 dipendenti.
Prof. G. Gilardi - Biological Chemistry
7
Vit B3: Formazione del
coenzima
La prima trasformazione è rappresentata dalla marcatura per
fosforilazione, transferasi cinasica, che destabilizza con il metallo
di transizione l’ultimo legame di ATP
Si forma fosfato catione che dà attacco elettrofilo
nell’ossidrile.
ATP
ADP + Pi
ACIDO FOSFOPANTOTENICO
OH
H 3C
PH O
C
CH 2
C
NH
CH3 O
Prof. G. Gilardi - Biological Chemistry
CH 2 O
CH 2 C
OH
8
Vit B3: Formazione del coenzima.
FOSFOPANTOTEINA
La seconda trasformazione riguarda il terminale carbossilico. (CISTEINA)
Intervengono 2 metalloproteine distinte
1) ospita nel sito attivo il terminale carbossilico e l’aa cisteina.
Il metallo di transizionesi lega all’O dell’OH virtuale sottrazione di H2O il carbossilecatione può ricevere attacco nucleofilo dall’NH2 della cisteina (virtuale saturazione di
H2O)
HO
OH
H3C
OH
O
P
HO
C
NH
CH
O
C
CH2
CH2
C
CH3
CH2
O
O
+
C
H2N
O
H
C
CH2 SH
OH
OH
H3C
OH
O
P
HO
O
CH2
NH
CH
C
C
CH3
O
CH2
CH2
NH
COOH
C
CH
O
CH2 SH
FOSFOPANTOTENILCISTEINA
Prof. G. Gilardi - Biological Chemistry
9
Vit B3: Formazione del coenzima.
FOSFOPANTOTEINA
2) il secondo enzima riconosce ed accoglie il terminale cisteinico
il Me riconosce l’O del carbossile (legato col doppio legame al C)
Interazione prima dativa poi covalente, sottrazione del carbossile come catione.
FOSFOPANTOTEINA
La cisteina diventa cisteamina
La fosfopantoteina può già essere molecola Coenzimatica.
OH
H3C
OH
O
P
HO
CH
C
O
CH2
NH
C
CH3
CH2
O
CH2
NH
C
CH
CH2 SH
O
Prof. G. Gilardi - Biological Chemistry
10
Enzimi a fosfopantoteina
L’enzima accettore ha nel sito attivo un residuo di Ser.
Poi per azione dell’E con il metallo di transizione che destabilizza il legame a cui partecipano i fosforili
E legato alla fosfopantoteina.
Enzima-CH2OH +ATPÆEnzima- CH2OP ÆEnzima- CH2OPP + P Pantoteina ÆEnzima- P-Pantoteina-SH
In queste proteine la fosfopantoteina è legata covalentemente all’apoproteina.
La fosfopantoteina si comporta come un braccio mobile (ex: sostituzione dimetilica) piuttosto lungo
con alternanza di tratti idrofobici ed idrofilici (sostituzioni alcoliche e legami carbammidici)
Questa alternanza di centri idrofilici ed idrofobici può creare interazioni con aa di diversi domini che
circoscrivono la zona in cui si inserisce il gruppo prostetico
braccio separatore e mobile
quando interagisce con gruppi diversi in diverse direzioni trascina con questi spostamenti e
indirizza in diverse direzioni il –SH che deve trasportare il substrato.
SUBSTRATO acili a catena più o meno lunga (dall’acido acetico-2C- a quello serotico a-26 C-)
Dagli acili più semplici a quelli a lunga catena.
La fosfopantoteina diventa CoE legato covalentemente ad enzimi di trasporto degli acili ed i
movimenti del “braccio spaziatore” sono trasmessi al –SH che è legato, con legame tioestere agli
acili e li può trasportare, orientarli verso altre proteine, enzimi.
Prof. G. Gilardi - Biological Chemistry
11
Vit B3: Formazione del
Coenzima A
La fosfopantoteina può essere intermedio nella sintesi di altri CoE.
Possibilità di interazione della fosfopantoteina con una struttura che mantiene un
derivato pirofosforico (PPi), cioè l’ATP.
Mn-Enzima Un Mn-Enzima riconosce e destabilizza il primo legame pirofosfato.
ATP + P PANTOTEINA Æ PPi + Defosfo CoASH
Interazione attacco nucleofilo che l’O del fosforile del cosubstrato può esercitare.
(sulla base dell’AMP si inserisce la Ppantoteina)
AMP + PPANTOTEINA
ATP → ADP + Pi (sull’OH 3’ del riboso dell’adenina)
DefosfoCoASH + ATP
ADP + CoASH
(ultimo passaggio→ CoA attivo)
E’ una reazione di trasferimento.
CoEnzimaA
Prof. G. Gilardi - Biological Chemistry
12
Coenzima A
NH2
N
N
OH
O
N
N
O
OH O
O P O
OH
H3C
OH
P
O
CH
C
O
CH2
NH
C
CH3
O
CH2
CH2
NH
C
CH
CH2 SH
O
OH
P O
HO
Prof. G. Gilardi - Biological Chemistry
13
CoA: interazione con
l’apoproteina e funzioni
Il CoA acquista così una carica più evidente che lo fa riconoscere dalle sue apoproteine.
La molecola non è lineare ma c’è un ripiegamento attorno alla struttura pirofosforica; questo fa sì che
l’adenina che può transire da lattimica a semilattamica, si può trovare affacciata al SH terminale.
Anche in questo caso il CoE agisce solo con metalli cofattori.
L’effetto è una variazione del pK del SH che può essere in grado di accettare il substrato (o,
cambiando il pK, di dissociarlo).
Il substrato è sempre un acile che si unisce sempre con legame tioestereo.
(dall’acetato→ acetilCoA all’acile più lungo, es.cerobilCoA).
L’interazione con la proteina è di 2 tipi
1) Interazioni idrofilico-idrofobiche sui domini esterni dell’apoproteina (come per la Ppantoteina)
(Qui il braccio non è molto oscillante ma può interagire.)
2) legami salini (più forti) col P in 3’ (nella forma inattiva desforilata non si ha il legame con
l’apoproteina)
L’enzima non si lega all’apoproteina.
Le interazioni saline sono dovute alla conformazione delle apoproteine (del sito attivo).
I cambiamenti di conformazione fanno variare di un fattore compreso tra 103 e 106 il coefficiente di
dissociazione CoE-apoproteina.
Il CoA + substrato (ACILE) possono trasferirsi a diverse proteine e ad ogni trasferimento può
avvenire una ulteriore trasformazione del substrato.
Prof. G. Gilardi - Biological Chemistry
14
Funzioni metaboliche vit B3
Gli enzimi trasportatori di acili interessano alcuni catabolismi ossidativi degli acili.
(β-ossidazione, ciclo di Krebs-indirettamente:respirazione cellulare- demolizione o
sintesi)
Lesione da carenza di B3 riguarda:
→ respirazione cellulare
→ metabolismo lipidico
Macroscopicamente: lesioni sui tessuti a flusso metabolico maggiore: muscolatura,
tessuto nervoso ed epiteli: nevriti, miositi (aberrazioni muscolari), infiammazioni degli
epiteli.
Prof. G. Gilardi - Biological Chemistry
15