■REVIEW ■
IL DOGMA INFRANTO
DALLE STAMINALI NEURALI
La scoperta che il cervello adulto non perde la capacità di generare neuroni
ha aperto un filone di ricerca ambizioso ma ancora disseminato di incognite
L U C A C O L U C C I D ’ A M AT O , C A R L A P E R R O N E C A PA N O , U M B E RT O D I P O R Z I O
L
E CELLULE STAMINALI, LE FONDATRICI di ogni cellula del corpo,
rappresentano uno dei campi più promettenti della ricerca biomedica sia per lo studio dei meccanismi patogenetici delle
malattie sia per lo sviluppo di nuove terapie. In particolare, la scoperta di staminali che danno origine al sistema nervoso e vi permangono
anche nella vita adulta, le staminali neurali, ha rivoluzionato alcuni
concetti fondamentali della neurobiologia. Fino ai primi anni ‘90 infatti dominava l’opinione che nel sistema nervoso centrale dei mammiferi la capacità di generare nuove cellule nervose (neurogenesi) si arrestasse definitivamente poco dopo la nascita.
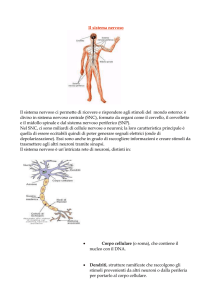
La formazione del sistema nervoso inizia nelle prime fasi embrionali, durante la gastrulazione, quando si formano i tre foglietti dai quali deriveranno tutti i tessuti: l’endoderma, il mesoderma e l’ectoderma. Dall’ectoderma, grazie a una complessa serie di interazioni cellulari, si forma
il tubo neurale, la struttura da cui deriverà il sistema nervoso centrale
composto dal cervello e dal midollo spinale. Al termine della formazione del tubo neurale, l’ectoderma originario è diviso in tre gruppi di cellule: quelle all’interno del tubo daranno vita al cervello e al midollo spinale; quelle all’esterno del tubo genereranno l’epidermide; le cellule situate nella giunzione fra tubo neurale ed epidermide, denominate cresta
neurale, migreranno in periferia a formare neuroni e glia del sistema nervoso periferico e intestinale e altri tipi cellulari in varie sedi dell’organismo. Qui discuteremo della capacità proliferativa della progenie del primo gruppo di cellule, quelle situate all’interno del tubo neurale, che costituiscono il cosiddetto neuroepitelio germinativo. Quest’ultimo permane dopo l’embriogenesi in un’area chiamata zona subventricolare, che
sarà il sito di origine delle staminali neurali nell’adulto.
«Una volta terminato lo sviluppo, le fonti di crescita e rigenerazione degli assoni e dei dendriti si prosciugano irrevocabilmente. Nei
centri nervosi dell’adulto le vie nervose sono in qualche modo fisse,
finite e immutabili. Ogni cosa può morire ma nulla può rigenerare. Sta
alla scienza del futuro cambiare, se possibile, questa dura legge» 1.
Questa era la convinzione del medico e istologo spagnolo Santiago
Ramon y Cajal, lungimirante studioso del sistema nervoso (oggi si
direbbe neurobiologo) che dominò la scena internazionale fra la
seconda metà dell’Ottocento e i primi decenni del Novecento, condividendo con l’italiano Camillo Golgi il Nobel per la Medicina nel
1906. Golgi, professore di patologia generale nell’Università di Pavia
dal 1881, non poteva che condividere questa convinzione, sia pure per
motivi diversi. Egli infatti, per spiegare la propagazione dell’impulso
elettrico tra centri distanti, riteneva erroneamente che il sistema ner-
52 ■ DARWIN ■ SETTEMBRE/OTTOBRE
CORTESIA SU-CHUN ZHANG, UNIVERSITY OF WISCONSIN-MADISON
Cellule della glia (verdi) e neuroni (rossi) maturi ricavati in vitro dalle staminali embrionali umane.
53 ■ DARWIN ■ SETTEMBRE/OTTOBRE
voso centrale fosse formato non da elementi cellulari distinti ma da un’unica rete fibrillare di connessioni, derivante dalla fusione di cellule.
D’altronde, fino a non molti anni fa generazioni
di studenti imparavano che i tessuti umani si suddividono in labili, come il sangue, stabili, come il fegato, e perenni, come il sistema nervoso, secondo la
classificazione fatta dal medico e istologo dell’Ottocento Giulio Bizzozero, maestro di Golgi e autore
del primo trattato italiano di istologia. Il sistema nervoso, insegnava Bizzozero, è composto di cellule prive di potenzialità replicativa e che durano tutta la vita 2. Questa convinzione è perdurata a lungo, per ragioni che possono essere ricondotte a quattro motivi principali.
■ Motivi clinici: è noto che i pazienti con lesioni del parenchima nervoso non recuperano le funzioni perse se non in misura marginale. Nelle malattie
cerebrovascolari, nelle lesioni traumatiche, nelle
malattie neurodegenerative croniche si osserva un
inesorabile peggioramento che le terapie possono rallentare ma mai revertire. Queste osservazioni sono
state a lungo interpretate come la prova di quanto
sostenuto dai grandi neurobiologi del passato. Si riteneva inoltre che questo fosse il prezzo pagato dall’evoluzione per raggiungere la raffinatezza e la complessità del sistema nervoso dei mammiferi.
■ Motivi evolutivo-teleologici: il sistema nervoso centrale presiede a funzioni di estrema delicatezza la cui minima modulazione risulta in cambiamenti comportamentali decisivi per la sopravvivenza.
Perciò si riteneva che l’eventuale attività proliferativa, con la migrazione e il differenziamento delle cellule neoformate, avrebbe inevitabilmente alterato la
peculiare e complessa comunicazione fra le cellule
nervose, scompaginando in modo insopportabile i
circuiti preesistenti. Inoltre, l’osservazione che nelle
cellule cancerose l’acquisizione della capacità di
proliferare incessantemente si accompagna alla perdita delle caratteristiche differenziate rafforzava l’idea che l’estremo differenziamento del sistema nervoso fosse incompatibile con una qualsiasi attività
proliferativa.
■ Teoria dell’apprendimento e della memoria: si
ipotizzava che il ricordo di quanto avvenuto fosse
dovuto al richiamo di circuiti neuronali stabili, definiti engrammi, formatisi con l’apprendimento. In
questo contesto la formazione di nuovi neuroni era
vista come incompatibile con la memoria.
■ Motivi tecnico-sperimentali: un potente impedimento alla dimostrazione della neurogenesi nel
sistema nervoso centrale è stata l’assenza di metodi e
strumenti adeguati. Solo in anni relativamente recenti l’evoluzione delle tecniche sperimentali, in particolare la microscopia, e l’identificazione di molecole
capaci di caratterizzare i diversi tipi di cellule nervo-
CORTESIA ERIC A. NEWMAN, UNIVERSITY OF MINNESOTA
■ REVIEW ■
se e le fasi della loro vita, hanno fornito ai ricercatori gli strumenti necessari.
Oggi è definitivamente provato che queste convinzioni erano errate e che vi è produzione di nuovi
neuroni, sia pur limitata, nel sistema nervoso centrale dell’adulto anche nei mammiferi e nell’uomo.
Pertanto possiamo dire con Fernando Nottebohm,
neurobiologo della Rockfeller University di New
York, che «oltre ogni ragionevole dubbio, il dibattito se la neurogenesi abbia luogo nel cervello dei vertebrati adulti a sangue caldo è chiuso» 3.
La scoperta può essere datata agli anni ‘60,
quando Joseph Altman e i suoi collaboratori del Mit
hanno dimostrato l’esistenza di cellule proliferanti in
una zona del cervello, chiamata giro dentato dell’ippocampo, mediante l’uso della timidina radioattiva.
La timidina, una delle quattro basi del DNA, si integra nel DNA delle cellule in divisione e permette di
visualizzarle con tecniche di autoradiografia.
Tuttavia la mancanza di marcatori molecolari non
aveva permesso di dimostrare con certezza che le
cellule che incorporavano la timidina sarebbero
diventate neuroni 4.
Negli anni ‘70 Michel Kaplan ha ripreso le
ricerche di Altman accoppiando all’autoradiografia
la microscopia elettronica e riuscendo così a dimostrare che le cellule marcate con timidina nel giro
dentato del ratto erano effettivamente neuroni 5.
Ciononostante la comunità scientifica è rimasta
scettica, indifferente o chiaramente ostile 6. Una
vera svolta ha avuto luogo grazie agli studi di
Nottebhom e collaboratori che negli anni ‘80, stu-
54 ■ DARWIN ■ SETTEMBRE/OTTOBRE
CORTESIA ERIC A. NEWMAN, UNIVERSITY OF MINNESOTA
■ REVIEW ■
Cellule della glia di retina di ratto trattate con anticorpi specifici per i diversi tipi cellulari e visualizzate in microscopia confocale con
immunofluorescenza. Nelle immagini a sinistra e al centro si vedono in blu i vasi sanguigni, in giallo gli astrociti e in rosso i prolungamenti delle cellule di Müller, un peculiare tipo di cellule gliali della retina. Nell’immagine a destra gli astrociti appaiono in blu e
le cellule di Müller in rosso. L’effetto tridimensionale è stato ottenuto combinando le immagini di più sezioni successive di tessuto.
diando le basi neurologiche del canto degli uccelli,
hanno dimostrato che vi è neurogenesi nel cervello
di uccelli adulti 3. Solo negli anni ‘90 il lavoro di differenti laboratori in Canada, in Australia e negli
Stati Uniti ha dimostrato che nel cervello dei mammiferi adulti esistevano staminali neurali che potevano essere isolate ed espanse in vitro 7. In seguito si
è osservato che esiste neurogenesi in vivo e che
nuovi neuroni vengono generati di continuo in particolari regioni del cervello adulto: nell’ippocampo
(una delle aree deputate alla formazione della
memoria) e nel sottile strato di cellule che riveste i
ventricoli cerebrali, chiamato ependima periventricolare o zona subventricolare. Oggi inoltre sono
state isolate staminali neurali da quasi tutte le aree
del cervello embrionale di mammiferi.
È opportuno rimarcare come queste osservazioni siano state rese possibili da una serie di progressi:
l’identificazione di proteine diffusibili, chiamate fattori di crescita, che aggiunte nel mezzo di coltura
inducono la proliferazione o il differenziamento
delle cellule in vitro; l’identificazione di marcatori
cellulari specifici, che hanno permesso di caratterizzare il fenotipo delle cellule generate in vitro
mediante tecniche di immunoistochimica (uso di
anticorpi e loro visualizzazione al microscopio
mediante marcatori visibili o fluorescenti); la sostituzione della timidina marcata con la bromodeossiuri-
dina, un analogo della timidina visualizzabile
mediante immunoistochimica; la nuova tecnica della
microscopia confocale, che permette di localizzare
in una stessa cellula più molecole usando fluorocromi dai colori differenti.
Una per tutte
La staminale neurale si definisce come una cellula in
grado sia di riprodurre se stessa sia di dar vita a tutte
le popolazioni cellulari del sistema nervoso, quali i
neuroni, che trasmettono l’impulso nervoso, l’astroglia, una popolazione mista con funzioni molteplici
tra cui il supporto e la nutrizione dei neuroni, e l’oligodendroglia che produce la mielina, la guaina che
riveste gli assoni del cervello e del midollo spinale
garantendo una veloce conduzione dell’impulso elettrico. Questo tipo di divisione, che genera una cellula figlia simile alla madre e una figlia diversa, è chiamata divisione asimmetrica e si distingue dalla forma
più comune, la divisione simmetrica, mediante la
quale una cellula madre genera due figlie uguali.
La divisione asimmetrica è una proprietà peculiare
delle staminali, che tuttavia vanno anche incontro a
divisione simmetrica. Le staminali inoltre sono capaci di dividersi indefinitamente.
In coltura le staminali neurali sopravvivono e
proliferano se ricevono specifici fattori mitogeni e
possono essere riconosciute per la capacità di dare
55 ■ DARWIN ■ SETTEMBRE/OTTOBRE
■ REVIEW ■
origine a strutture chiamate neurosfere, ovvero
aggregati misti di staminali e precursori neurali che
si moltiplicano in vitro in particolari condizioni.
Non è tuttora chiaro se la staminalità sia un’entità a
cui corrispondono precisi connotati molecolari o
piuttosto una funzione che si attiva in particolari
condizioni.
Il punto è rilevante perché, se diversi tipi cellulari in condizioni adeguate possono assumere le
caratteristiche di cellula staminale (come recentemente dimostrato per un particolare tipo di cellule
gliali, chiamato glia radiale), allora la conoscenza
approfondita di queste condizioni aprirebbe la strada a una manipolazione farmacologica della neurogenesi.
Resta inoltre da chiarire se il differenziamento
della staminale neurale segua un meccanismo di tipo
sequenziale-lineare, per cui da ogni tipo cellulare a in
presenza di un dato fattore deriva necessariamente il
tipo cellulare b, oppure stocastico-probabilistico, per
cui un progenitore genera più tipi di cellule che, a
seconda degli stimoli che incontrano in una determinata finestra spaziotemporale, possono sopravvivere,
differenziarsi o morire. I due modelli potrebbero non
essere mutuamente esclusivi.
Come durante l’embriogenesi, anche nel cervello
adulto e in vitro le staminali e i precursori neurali possono differenziarsi e produrre vari tipi di neuroni e di
cellule gliali, sia spontaneamente sia in seguito a manipolazioni sperimentali. La diversificazione di una
popolazione relativamente omogenea di staminali
neurali in una miriade di tipi cellulari diversi è un processo complesso e richiede una molteplicità di fattori,
spesso in una precisa sequenza.
Semplificando molto possiamo dire che segnali
diffusibili presenti nell’ambiente in cui crescono le cellule, rilasciati per esempio da altre cellule, si distribuiscono secondo gradienti di concentrazione e vengono recepiti da quelle cellule che possiedono gli appropriati recettori. Questi segnali possono anche essere situati sulla superficie di cellule circostanti o nel
supporto su cui poggiano le cellule, la matrice extracellulare. Nella cellula che recepisce i segnali, i recettori che si attivano determinano una cascata di segnalazioni intracellulari che possono raggiungere il nucleo e attivare o spegnere specifici gruppi di geni.
Questi a loro volta dirigono la sintesi di proteine che
determinano morfologia e funzioni specifiche. Il susseguirsi di questi eventi porta infine al differenziamento delle cellule.
Allo stato attuale le conoscenze su questi meccanismi molecolari sono ancora insufficienti e derivano in prevalenza da studi in vitro, e va da sé che il
loro approfondimento è il prerequisito essenziale per
un sicuro impiego terapeutico. Inoltre a tutt’oggi non
c’è alcuna certezza che i neuroni generati in vitro
dalle staminali siano davvero equivalenti per struttura e funzioni a quelli dell’organismo. Questi limiti
andrebbero tenuti nel giusto conto quando si prospettano applicazioni terapeutiche a breve.
Dagli uccelli agli umani
Studi sempre più numerosi indicano che si può
indurre la neurogenesi nel cervello di uccelli e mammiferi adulti a partire da staminali neurali. I pionieristici studi del gruppo di Nottebohm avevano dimostrato che in uccelli canterini, come i canarini (Serinus
canaria) e il diamante mandarino (Taeniopygia guttata),
nuovi neuroni si integrano nei circuiti deputati all’apprendimento e all’esecuzione del canto. La produzione di questi neuroni si attiva in primavera, è sotto
controllo ormonale e risente del ciclo circadiano.
All’inizio tali fenomeni si ritenevano peculiari del
sistema del canto negli uccelli, ma è noto da secoli
che gli uccelli apprendono la loro canzone e oggi è
chiaro anche che questo apprendimento avviene con
meccanismi sovrapponibili all’apprendimento del
linguaggio umano.
Dopo gli uccelli, la neurogenesi è stata riconosciuta anche in due aree cerebrali dei mammiferi
adulti: la zona subventricolare dei ventricoli laterali
(cavità situate al di sotto della corteccia cerebrale) e
la zona subgranulare dell’ippocampo, una struttura
del lobo temporale implicata nell’apprendimento e
nella memoria. Nell’ippocampo le cellule proliferanti danno origine a neuroni del giro dentato. Nella
zona subventricolare, i neuroni in via di differenziamento (neuroblasti) che si generano si organizzano
in catene e migrano tangenzialmente, formando la
cosiddetta corrente migratoria rostrale, fino a raggiungere i bulbi olfattivi. Qui si differenziano e svolgono un ruolo di collegamento tra i principali neuroni dei bulbi olfattivi, modulando la codificazione
spaziale e temporale dell’informazione olfattiva.
I nuovi neuroni prodotti dalle staminali migrano
anche nella corteccia cerebrale, dove possono differenziarsi e sviluppare assoni funzionali che si integrano in nuovi circuiti. Lo hanno dimostrato Gould e
Gross, dell’Università di Princeton 8, che hanno messo in luce cellule marcate con bromodeossiuridina, e
quindi neoformate, in varie aree della corteccia cerebrale di scimmia. Gli assoni di questi neuroni si erano
accresciuti raggiungendo zone molto distanti dal corpo cellulare.
I neuroni neoformati avevano colonizzato la regione prefrontale, che controlla le decisioni esecutive e la memoria a breve termine, nonché la regione
temporale inferiore, importante per il riconoscimento visivo degli oggetti e delle facce, e la regione parietale posteriore, necessaria per la rappresentazione
spaziale degli oggetti. Pertanto le staminali neurali
avevano prodotto nuovi neuroni che sembravano es-
56 ■ DARWIN ■ SETTEMBRE/OTTOBRE
CORTESIA SU-CHUN ZHANG, UNIVERSITY OF WISCONSIN-MADISON
■ REVIEW ■
Cellule neurali ricavate dalle staminali embrionali umane. I motoneuroni sono visibili in rosa, le fibre nervose in verde.
sersi integrati nelle aree corticali per contribuire alle
funzioni cognitive superiori.
Anche alcuni stati patologici sembrano indurre la
neurogenesi nel cervello adulto. L’ictus ischemico
indotto sperimentalmente nei ratti stimola una sorprendente proliferazione di cellule della zona subventricolare che migrano e colonizzato la zona lesa,
adiacente ai ventricoli laterali, dove si differenziano
nei neuroni specifici di quest’area. Questa neurogenesi continua per molto tempo dopo la lesione e i
neuroni neoformati sopravvivono a lungo. È ipotizzabile che un tale tipo di neurogenesi abbia luogo
anche nell’uomo e spieghi alcuni aspetti del recupero funzionale dopo l’ictus.
Per quanto le ricerche sulla manipolazione della
neurogenesi siano ancora in fase iniziale, varie osservazioni degli ultimi anni convergono sulla possibilità che la neurogenesi sia modificabile da varie sollecitazioni ambientali, ormonali e farmacologiche.
Resta da approfondire il contributo funzionale dei
neuroni neoformati nel mammifero adulto, ma studi
recenti hanno dato indicazioni positive, dimostrando che i nuovi neuroni si integrano nei circuiti preesistenti e sono funzionali sia nell’ippocampo e nel
bulbo olfattivo sia nella corteccia cerebrale. Inoltre è
dimostrato che manipolazioni della proliferazione o
del differenziamento delle staminali neurali alterano
il comportamento e manipolazioni del comportamento alterano la neurogenesi. Per esempio nei roditori gli antidepressivi stimolano la neurogenesi ippocampale, la cui inibizione riduce gli effetti di questi
farmaci, il che suggerisce un inaspettato ruolo della
neurogenesi ippocampale nella regolazione degli
stati dell’umore e la possibilità di una sua manipolazione farmacologica 8.
È noto che la neurogenesi nel giro dentato dell’ippocampo è influenzata da stimoli quali l’attività
motoria, lo stress, l’età, gli ormoni, il trattamento con
antidepressivi, l’apprendimento e un ambiente ricco
di sollecitazioni. D’altro canto l’inibizione della neurogenesi blocca alcune funzioni di apprendimento
dipendenti dall’ippocampo, confermando che i neuroni neoformati nell’adulto partecipano alla formazione della memoria e ai processi cognitivi.
Analogamente si è visto che la neurogenesi nel bulbo
olfattivo è modulata dagli odori e che la sua inibizione determina deficit di discriminazione olfattiva,
suggerendo che essa abbia un profondo effetto sulla
memoria e sulle funzioni olfattive, essenziali per la
sopravvivenza in molti animali 10.
Risultati recenti, infine, fanno pensare che l’attività neurale possa influenzare il differenziamento
57 ■ DARWIN ■ SETTEMBRE/OTTOBRE
CORTESIA SU-CHUN ZHANG, UNIVERSITY OF WISCONSIN-MADISON
■ REVIEW ■
Dalle staminali embrionali sono stati ricavati precursori neurali che, trapiantati nel cervello di topi giovani, hanno generato neuroni (a sinistra) e astrociti (a destra) funzionanti. Nella pagina seguente, coltura in laboratorio di cellule staminali umane.
delle staminali cerebrali e la sopravvivenza delle cellule neoformate. In altre parole, i nuovi neuroni che
vengono usati sembrano avere maggiori possibilità di
sopravvivenza rispetto a quelli che non vengono
usati. In questa luce, la neurogenesi nell’adulto
potrebbe essere una forma di adattamento a un
nuovo ambiente, necessaria nel momento in cui c’è
bisogno di una maggiore plasticità.
Va sottolineato che la dimostrazione della neurogenesi ippocampale e corticale nell’adulto richiede
una rielaborazione delle teorie cognitive e della
memoria, finora basate sull’assunto che il cervello
adulto fosse costituito da una rete stabile di neuroni
e che la sua plasticità fosse limitata a modificazioni
delle sinapsi, le strutture deputate alla trasmissione
dell’informazione tra neuroni.
Conversioni e abbagli
Si può riprogrammare una cellula staminale neurale?
Nel 1999 Angelo Vescovi e collaboratori hanno riferito questo risultato nel topo. Nel sistema vascolare
degli animali sono state iniettate staminali neurali
marcate, quindi identificabili anche dopo la proliferazione e il differenziamento. Dopo un certo tempo
in questi topi sono state osservate cellule ematopoietiche che esprimevano il tracciante delle staminali
neurali: le staminali neurali, di origine ectodermica,
si erano quindi convertite in cellule del sistema ematopoietico che è di origine mesodermica 11. Tale
fenomeno – la conversione di una staminale appartenente a un dato foglietto embrionale in una cellula di
origine diversa – è stato definito transdifferenziazione e implica una riprogrammazione nucleare, con l’espressione di alcuni gruppi di geni e lo spegnimento
di altri. In seguito sono stati descritti altri casi analoghi: dalle staminali neurali si sono ottenute cellule di
fegato, polmone, intestino e rene ed è stato realizzato anche il percorso inverso, ricavando neuroni da
cellule ematopoietiche. È verosimile che il fenomeno
avvenga anche nell’uomo, come suggerisce l’osservazione, nei cervelli autoptici di pazienti che avevano
ricevuto un trapianto di cellule ematopoietiche, di
neuroni originati dalle cellule trapiantate.
La transdifferenziazione potrebbe permettere di
ottenere neuroni da tessuti adulti diversi dal cervello
embrionale, magari prelevati dallo stesso paziente
che dovrà ricevere il trapianto, così da generare cellule immunologicamente compatibili ed evitare terapie immunosoppressive. Tuttavia la reale efficacia del
processo resta molto dubbia. Innanzitutto, non è
affatto certo che le cellule nervose generate per
transdifferenziazione – o anche da staminali neurali
in vitro – siano del tutto equivalenti ai neuroni endogeni. Per di più si è osservato che le staminali tra-
58 ■ DARWIN ■ SETTEMBRE/OTTOBRE
■ REVIEW ■
Note
1. Santiago Ramon y Cajal. Estudios sobre la degeneración y regeneración del sistema nervioso. Vol. 2, 1913-14.
2. Bizzozero. Accrescimento e rigenerazione nell’organismo. Archivio
per le Scienze Mediche, 1893.
3. Nottebohm F. Why are some neurons replaced in adult
brain? J Neurosci 2002; 22: 624-8.
4. Altman J. Are new neurons formed in the brains of adult
mammals? Science 1962; 135: 1127-8.
5. Kaplan MS, Hinds JW. Neurogenesis in the adult rat: electron microscopic analysis of light radioautographs. Science
1977; 197: 1092-4.
6. Hope for a New Neurology, Vol. 457, New York Academy of
Sciences, USA.
7. Reynolds BA, Weiss S. Generation of neurons and astrocytes from isolated cells of the adult mammalian central
potenziale di riparare il sistema nervoso danneggiato. In modelli animali di malattia, le staminali si
sono dimostrate capaci di migliorare o curare patologie causate da differenti meccanismi quali gliomi,
sclerosi a placche, malattie neurodegenerative e
ictus cerebrale.
Non sarebbe sorprendente se si scoprisse che il
loro potenziale terapeutico non risiede solo nella
possibilità di rimpiazzare le cellule danneggiate o
perdute, ma anche in una
più vasta e per ora elusiva
capacità di modificare
l’ambiente cerebrale malato, per esempio mediante
il rilascio di fattori di crescita. Tuttavia le speranze
terapeutiche potranno essere soddisfatte solo se si
comprenderà come controllare la proliferazione
delle cellule, come ricavarne i giusti tipi di neuroni, come inserire i nuovi
neuroni nei circuiti nervosi esistenti, come evitarne il
rigetto dopo il trapianto.
È dunque prematuro parlare di staminali, embrionali o adulte, come di una panacea. Al momento
esistono solo approcci sperimentali condotti in prevalenza in vitro e in modelli animali. Gli obiettivi terapeutici non saranno raggiungibili in modo riproducibile e scevro da rischi fin quando non si avrà una conoscenza approfondita dei meccanismi fondamentali
che regolano la vita della cellula, dei neuroni e del
cervello e dei processi dello sviluppo e del differenziamento neuronale.
CORTESIA DI JEFF MILLER, UNIVERSITY OF WISCONSIN-MADISON
piantate in vivo possono fondersi con cellule dell’organismo ospite, e che in vitro le staminali ematopoietiche possono fondersi con le neurosfere; si può quindi ipotizzare che un fenomeno simile sia alla base di
alcune conversioni interpretate in precedenza come
transdifferenziazione. La distinzione tra eventi di
transdifferenziazione e di fusione cellulare resta un
problema irrisolto per la mancanza di marcatori che
discriminino tra i due fenomeni. Anche per questo
allo stato attuale la pretesa equivalenza tra staminali adulte ed embrionali,
talora accampata per
motivi estranei al dibattito scientifico, è tutt’altro
che dimostrata ed è un
errore voler contrapporre
una fonte all’altra.
Inoltre, riguardo alla
riprogrammazione nucleare che avrebbe luogo
nella transdifferenziazione, occorre tenere presenti recenti studi di trasferimento nucleare, che
mostrano come il nucleo della cellula donatrice
conservi una certa memoria (detta epigenetica)
della sua origine: nel rospo Xenopus, per esempio, il
nucleo di una cellula ectodermica trasferito in un
oocita denucleato riesprime marcatori ectodermici
anche nell’endoderma. Quindi non si può escludere
che geni accesi nel nucleo somatico continuino a
esprimersi anche ectopicamente se questo nucleo
viene inserito in un oocita che poi si differenzia in
vari tessuti 12. Di qui nasce la necessità di ulteriori
studi e di grandi cautele nel prevedere l’uso di staminali transdifferenziate o generate per trasferimento di nucleo.
È comprensibile che le staminali neurali suscitino un enorme interesse per il loro straordinario
Luca Colucci D’Amato, Carla Perrone Capano, Umberto di Porzio,
Laboratorio di Neurobiologia, Istituto di Genetica e Biofisica Adriano
Buzzati Traverso, Cnr, Napoli
nervous system. Science 1992; 255: 1707-10.
8. Gould E et al. Neurogenesis in the neocortex of adult primates. Science 1999; 286: 548-552
9. Santarelli L et al. Requirement of hippocampal neurogenesis for the behavioral effects of antidepressants. Science 2003;
301: 805-9.
10. Doetsch F, Hen R. Young and excitable: the function of
new neurons in the adult mammalian brain. Curr Opin Neurobiol
2005; 15: 121-8.
11. Bjornson CR et al. Turning brain into blood: a hematopoietic fate adopted by adult neural stem cells in vivo. Science
1999; 283: 534-7.
12. Ng RK, Gurdon JB. Epigenetic memory of active gene
transcription is inherited through somatic cell nuclear transfer.
PNAS 2005; 102: 1957-62.
59 ■ DARWIN ■ SETTEMBRE/OTTOBRE