Il catabolismo dei lipidi e l’ossidazione
degli acidi grassi
Nella maggior parte degli organismi i lipidi si
trovano
principalmente
nella
forma
di
TRIACILGLICEROLI (trigliceridi)
In molte piante una grossa quantità di trigliceridi
sono conservati nei semi, nei frutti e nei granuli
pollinici (eccezione jojoba che accumula acidi
grassi e cere nei semi)
I lipidi di deposito vegetali sono anche
un’importante fonte di grassi alimentari per l’uomo
e gli altri animali
Industria manifatturiera: produzione detergenti,
rivestimenti, materie plastiche e lubrificanti.
I lipidi rappresentano una enorme riserva
energetica. Questo in virtù del fatto che il carbonio
nei triacilgliceroli presenta un grado di riduzione
maggiore di quello presente nei carboidrati
Non bisogna dimenticare la presenza del gruppo
carbossilico, altamente ossidato, ma la maggior
parte del carbonio è in forma metilenica, quindi
altamente ossidata
Da ciò ne deriva che l’ossidazione dei grassi
consuma più ossigeno, in rapporto al peso,
dell’ossidazione dei carboidrati e rilascia una
quantità superiore di energia metabolica
Negli animali, i grassi provengono da tre vie:
- alimentazione
- biosintesi ex-novo nel fegato
- mobilizzazione
adipociti
del
grasso
accumulato
negli
La mobilizzazione dei lipidi negli animali è operata
dalle proteine [(very low density lipoprotein (VLDL),
low density lipoprotein (LDL)
high density
lipoprotein (HDL)]
Nei semi deidratati, prima della germinazione, vi è
una bassissima attività enzimatica, mentre durante
l’imbibizione si ha un incremento di attività di un
gran numero di enzimi tra i quali le lipasi dei
trigliceridi
I loro substrati sono contenuti in gocce di olio
(oleosomi) dentro i semi e gli enzimi agiscono sulla
superficie delle gocce, probabilmente con l’aiuto di
proteine leganti per facilitare il processo
Gli
enzimi
deputati
alla
scissione dei trigliceridi sono
idrolasi
più
propriamente
chiamate LIPASI
Le lipasi catalizzano il rilascio di
acidi grassi esterificati sulle
posizioni 1 e 3 dei trigliceridi
per
ottenere
un
monoacilglicerolo
Il rilascio di tutti e tre gli acidi
grassi è ottenuto attraverso la
migrazione dell’acido grasso
dalla posizione 2 alla posizione
1 e il suo susseguente rilascio
Il glicerolo rilasciato viene convertito ad un
intermedio
della
Glicolisi,
il
DIIDROSSIACETONFOSFATO, in una reazione
catalizzata dalla GLICEROLO CHINASI e dalla
GLICEROLO FOSFATO DEIDROGENASI
La scissione dei fosfolipidi
Gli enzimi coinvolti nella scissione e
rimodellazione dei fosfolipidi sono chiamati
FOSFOLIPASI
Nelle piante e nei tessuti animali, un gran
numero di fosfolipasi è stato identificato in
funzione del sito di attacco sulla molecola del
fosfolipide
Il composto risultante che contiene solo un acido
grasso, nella posizione 1 o 2, è chiamato lisofosfolipide.
Questi enzimi sono coinvolti nella modificazione della
composizione in acidi grassi delle membrane
fosfolipidiche esistenti
La fosfolipasi B sembra essere presente solo nei
microrganismi, e poiché agisce sia sulla posizione 1
che 2, è capace di agire sia sui fosfolipidi intatti che sui
fosfolipidi liberi
La fosfolipasi C ha un ruolo importante nel controllo
dell’attività enzimatica. Molti enzimi sono regolati da un
meccanismo di fosforilazione/defosforilazione calcio
dipendente
Anche se la fosfolipasi D è stata trovata nei tessuti
animali, la fonte principale di quest’enzima sono i
tessuti vegetali.
E’ un enzima molto attivo coinvolto nella reazione di
scambio degli acidi fosfatidici; comunque il suo ruolo
nel metabolismo dei fosfolipidi non è ancora del tutto
chiaro
Catabolismo dei glicolipidi
Le piante sono molto ricche di glicolipidi. Il loro
catabolismo è particolarmente attivo durante la
senescenza ed il susseguente danno ai tessuti
La degradazione completa avviene in due stadi:
1) gli acidi grassi sono rimossi dalla acil-idrolasi
2) gli zuccheri residui sono rimossi per azione delle
galattosidasi
Nei ruminanti una grande quantità dei lipidi ingeriti
sono degradati per azione dell’acil-idrolasi e della
glicosidasi batterica
L’ossidazione dei grassi avviene mediante la betaossidazione
Negli animali questa avviene nel mitocondrio o nel
perossioma, mentre nelle piante e in molti funghi
avviene solo nel perossisoma
In realtà l’ossidazione avviene nel perossisoma degli
organismi animali solo quando la catena è troppo
lunga per potere entrare nel mitocondrio anche se
si blocca quando la catena è formata da 8 atomi di
C e quindi prosegue nel mitocondrio
Come vedremo nel caso della ossidazione
perossisomale non si produce ATP ed invece
perossido di idrogeno che è convertito ad acqua ed
ossigeno dalla catalasi
Ossidazione degli acidi grassi
Gli acidi grassi possono andare incontro ad una serie di
modificazioni ossidative
Quantitativamente il percorso biochimico più importante
nell’ossidazione degli acidi grassi è la –OSSIDAZIONE
Questo processo avviene in tutti gli organismi ed è la
principale via attraverso cui gli acidi grassi sono
utilizzati per produrre energia
Nella –ossidazione, gli acidi grassi sono degradati a
composti più piccoli, solitamente acetil-CoA, che può
poi essere ossidato attraverso il ciclo TCA
La conversione delle lunghe catene degli acidi
grassi in acetil-CoA e la sua successiva
ossidazione nel ciclo TCA porta alla produzione di
una grossa quantità di coenzimi ridotti quali NADH
e FADH2, che verranno utilizzati per la produzione
di ATP
Nelle piante e nei funghi, inoltre, la –ossidazione
può essere utilizzata per ottenere acetil-CoA per la
sintesi dei carboidrati tramite la via del gliossilato
Due altri processi ossidativi, conosciuti come ossidazione e -ossidazione, sono coinvolti nella
modificazione degli acidi grassi
-ossidazione: provoca l’ossidazione degli acidi
grassi sul carbonio in posizione 2 ( rispetto al
carbonile)
-ossidazione: determina l’ossidazione degli acidi
grassi sul gruppo metile terminale cioè sul
carbonio
–ossidazione
Quasi tutti i tessuti sono in grado di ossidare gli
acidi grassi tramite la –ossidazione
Nella cellula sono presenti due principali siti dove
avviene la –ossidazione
Negli animali è la matrice mitocondriale il luogo in
cui gli acidi grassi sono completamente ossidati da
acetil-CoA. Nel fegato e nei reni riveste invece
maggior importanza l’ossidazione che avviene nei
perossisomi
La –ossidazione nei perossisomi sembra essere
un meccanismo utilizzato principalmente per
ridurre la lunghezza delle catene degli acidi grassi,
che
possono
essere
successivamente
completamente ossidati nella matrice mitocondriale
Nei tessuti fogliari delle piante, sembrano i
perossisomi più che i mitocondri, gli organelli
deputati all’ossidazione degli acidi grassi, mentre
nei semi i gliossisomi sono i siti principali
dell’ossidazione degli acidi grassi, in quanto il
prodotto dell’ossidazione, l’acetil-CoA, può entrare
direttamente nel ciclo del gliossilato
–ossidazione mitocondriale
Gli acidi grassi metabolizzati dalla –ossidazione
possono derivare sia da fonti esogene (alimenti) che
endogene (esempio tessuto adiposo e fegato)
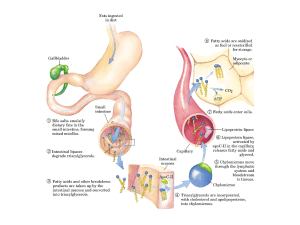
Il trasporto degli acidi grassi nel sangue avviene nella
forma non esterificata
(NEFAs) con la proteina
trasportatrice, che normalmente è l’ALBUMINA,
prodotta dal fegato.
Altrimenti vengono trasportati come fosfolipidi e
trigliceridi
incorporati
nelle
lipoproteine,
principalmente lipoproteine a bassissima densità
(VLDL)
Gli acidi grassi legati alle lipoproteine sono rilasciati
nel sangue attraverso l’azione della lipoproteina
lipasi, enzima localizzato sulla superficie del lumen
delle cellule capillari dell’endotelio dei capillari
I NEFAs passano dal sangue alle cellule adiacenti sia
per diffusione sia per un processo mediato della
membrana
ATTIVAZIONE DELL’ACIDO GRASSO
Nel citoplasma, gli acidi grassi devono essere
attivati come esteri del CoA prima di essere
successivamente metabolizzati
La reazione richiede ATP ed è catalizzata da un
enzima chiamato ACETIL-CoA SINTASI
L’acido grasso così attivato sul lato citosolico della
membrana mitocondriale esterna può essere trasportato
nel mitocondrio oppure essere utilizzati nel citosol per la
sintesi dei lipidi di membrana
Una serie di enzimi catalizzano questa reazione, ma
hanno differente specificità per la lunghezza della
catena. I due più importanti nell’ossidazione degli
acidi grassi sono:
• acetil-CoA sintasi a media catena (C4-C12)
• acetil-CoA sintasi a lunga catena (C10 e più)
L’acetil-CoA sintasi a lunga catena è un enzima
legato alla membrana dei perossisomi, nel reticolo
endoplasmatico e nella membrana mitocondriale
esterna
Questa distribuzione permette agli acidi grassi di
essere metabolizzati attraverso diversi percorsi
metabolici in differenti siti sub-cellulari
Ad esempio la riduzione della lunghezza della catena
che avviene nei perossisomi, la sintesi dei fosfolipidi
e trigliceridi nel reticolo endoplasmatico e la –
ossidazione nei mitocondri
La membrana mitocondriale interna costituisce una
barriera agli acetil-CoA destinati all’ossidazione. Né il
CoA né gli esteri del CoA possono attraversarla.
Questo è dovuto principalmente alle grandi dimensioni
ed alla natura delle cariche presenti nella molecola del
CoA
Per facilitare il movimento degli acidi grassi attraverso
la membrana interna mitocondriale verso la matrice, gli
acetil-CoA sono convertiti in acil-carnitina ad opera
dell’enzima carnitina-aciltransferasi I, localizzata nello
spazio intermembrane mitocondriale
Il complesso Acido grasso-carnitina si forma attraverso
la membrana interna tramite una proteina di trasporto,
la Acil-carnitina Translocasi, che attua un meccanismo
di antiporto (entrata della Carnitina e uscita della Acido
grasso-carnitina).
Nella matrice mitocondriale l’acido grasso viene
trasferito dalla Carnitina di nuovo al CoA grazie alla
Carnitina aciltransferasi II. Una volta entrati nella
matrice mitocondriale come estere del CoA gli acidi
grassi subiscono la –ossidazione.
Le reazioni della –ossidazione
La –ossidazione è costituita da una sequenza di
quattro reazioni necessarie per rimuovere un’unità
bicarboniosa della terminazione carbossilica
dell’acido grasso
Questa sequenza di reazioni è ripetuta fino a
quando l’acido grasso è completamente ossidato
1) Reazione iniziale di deidrogenazione
Il primo passo della –ossidazione di un acil-CoA è la
rimozione di un atomo di idrogeno dai carboni 2 () e 3 ()
con formazione del 2 TRANS-ENOIL-CoA
Si tratta di un’ossidazione operata da tre diverse ACETLCoA DEIDROGENASI che agiscono su acidi grassi a
catena di diversa lunghezza (corta, media e lunga)
Ciascuna di esse ha tuttavia come gruppo prostetico il
FAD. Gli atomi di idrogeno sono accettati dal FAD che
viene ridotto a FADH2
Dal FADH2 gli elettroni sono
traslocati ad una flavoproteina che
trasferisce elettroni (ETF) che, a
Sua volta li trasferisce alla catena
di trasporto mitocondriale
2) Idratazione e deidrogenazione
Il passo successivo della reazione è l’idratazione
della molecola. L’enzima responsabile di questa
reazione è l’ENOIL-CoA IDRATASI
Questo enzima catalizza l’addizione stereospecifica
dell’acqua sul doppio legame con formazione
dell’L-3-IDROSSIALCIL-CoA
Successivamente si ha una deidrogenazione NAD+dipendente. L’enzima specifico è la 3-IDROSSIACIL-CoA
DEIDROGENASI. Questo enzima è relativamente non
specifico per la lunghezza della catena, ma è estremamente
specifico riguardo allo stereoisomero L del 3-idrossiacilCoA
L’enzima catalizza la rimozione di due atomi di idrogeno dal
carbonio 1 ed il loro trasferimento al NAD+. Il prodotto della
reazione è il 3-CHETACIL-CoA
3) Rottura del legame tioestere
Il passo finale nel ciclo di reazioni della –
ossidazione è la rottura tiolitica del legame del 3acetoacil-CoA tra i carboni 2 e 3 ad opera del CoA
L’enzima che opera la rottura è chiamato –
CHETOTIOLASI. Un gruppo tiolico essenziale
dell’enzima (E-SH) attacca il substrato portando
alla formazione dell’enzima acilato e di acetil-CoA
come intermedi
Resa energetica della
–ossidazione
Le due riserve più usate dalle cellule per la produzione di
Acetil-CoA sono il glucosio e gli acidi grassi. Gli acidi grassi a
lunga catena sono composti altamente ridotti e dunque la loro
completa ossidazione può condurre alla produzione di un
grande quantitativo di ATP
La completa –ossidazione di una mole di acido palmitico
produce 8 moli di acetil-CoA che può essere successivamente
ossidato attraverso il ciclo TCA
Una molecola di ATP è idrolizzata ad AMP e PPi per
l’attivazione degli acidi grassi prima del trasporto della matrice
mitocondriale
TABELLA 14.1 Resa in ATP dell’ossidazione dell’acido palmitico (C16)
Percorso
Conversione netta
Cofattori
prodotti
Resa in ATP
–ossidazione
Da acido palmitico a 8
acetil-CoA
7 NADH
21 ATP
7 FADH2
14 ATP
24 NADH
72 ATP
8 FADH2
16 ATP
8 GTP
8 ATP
Totale
131 ATP
Ciclo TCA
8 Acetil-CoA a CO2 e
acqua
Una molecola di ATP è idrolizzata ad AMP e PPi per
l’attivazione degli acidi grassi prima del trasporto
nella matrice mitocondriale
Questo equivale all’utilizzazione di 2 ATP, quindi
si ha una sintesi netta di 129 moli di ATP dalla beta
ossidazione di una mole di acido palmitico
–ossidazione degli acidi grassi
a catena dispari
La –ossidazione degli acidi grassi a catena dispari
è in parte simile a quella degli acidi grassi a catena
pari
Il processo procede attraverso la rimozione
sequenziale di acetil-CoA fino a quando non
rimane l’intermedio a cinque atomi di cabonio.
Questo è quindi convertito in una molecola di
acetil-CoA e una molecola di propionil-CoA
Poiché il propionil-CoA è un composto a tre atomi di
carbonio questo può essere utilizzato per la sintesi del
glucosio
Comunque, la quantità di acidi grassi a catena dispari
trovata nei tessuti animali è solitamente piccola (1-2%) e
dunque il contributo alla produzione di glucosio è
trascurabile
in
confronto
con
altri
composti
gluconeogenetici
Questa è negli animali una delle poche possibilità di
conversione di un prodotto dell’ossidazione degli acidi
grassi in glucosio
In questo caso il succinil-CoA
entra nel ciclo TCA e viene
demolito per la produzione di
energia
La –ossidazione degli acidi grassi
insaturi
Una –ossidazione leggermente modificata rispetto a quella
tipica è necessaria per l’ossidazione degli acidi grassi insaturi
L’intermedio insaturo prodotto dall’Acil-CoA deidrogenasi ha la
configurazione 2–Trans, mentre negli acidi grassi insaturi si
riscontra un doppio legame in configurazione cis
Poiché gli acidi grassi insaturi sono ossidati tramite la
di una unità bicarboniosa dalla coda carbossilica, la
dei doppi legami si sposta mano a mano che la
accorcia sul carbonio del gruppo carbossilico che è
carbonio 1
rimozione
posizione
catena si
sempre il
Tale situazione ha due importanti conseguenze nella –
ossidazione degli acidi grassi insaturi
Per un acido grasso come l’acido oleico che ha un
doppio legame cis tra il carbonio 9 e 10, è
necessaria una reazione addizionale per fare
avvenire la –ossidazione
Il primo passo è la rimozione dei sei atomi di
carbonio (tre molecole di Acetil-CoA) attraverso il
solito percorso della –ossidazione
Il risultato è la produzione di un intermedio enoilCoA di 12 atomi di carbonio (3 cis-dodecanoilCoA) in cui il doppio legame è nella posizione
sbagliata 3 invece che 2, e nella configurazione
errata cis anziché trans
La posizione e la configurazione del doppio
legame vengono modificate grazie all’enzima
enoil-CoA isomerasi
Questo enzima converte il doppio legame 3 cis in
un doppio legame 2 trans che permette alla –
ossidazione di continuare regolarmente
Attraverso l’utilizzo di questi due enzimi
addizionali, l’enoil-CoA isomerasi e la 2,4-dienoilCoA riduttasi, quasi tutti gli acidi grassi presenti in
natura possono essere ossidati tramite la –
ossidazione
CONTROLLO DELLA Β- OSSIDAZIONE
- Trasporto degli acil-coA nei mitocondri (CAT è
inibita da malonil-CoA)
- Rapporto NAD/NADH
- AcetilCoA inibisce la tiolasi
La –ossidazione degli acidi grassi nei
perossisomi e negli gliossisomi differisce da quella
dei mitocondri
La prima reazione è catalizzata dall’ACIL-CoA
OSSIDASI e si ha produzione di perossido di
idrogeno, che è poi scisso dall’azione della catalasi
Le rimanenti reazioni del ciclo di –ossidazione
sembrano essere le stesse di quelle dei mitocondri,
anche se potrebbero esserci differenze strutturali e
funzionali degli enzimi nei diversi organelli
subcellulari
La membrana dei perossisomi e gliossisomi non
costituisce, al contrario della membrana interna
mitocondriale, una barriera di permeabilità ai
derivati degli acidi grassi collegati al CoA
Quindi non è necessario che gli acidi grassi
vengano trasportati dalla carnitina prima di essere
ossidati
Differentemente dai mitocondri, i perossisomi e
gliossisomi non contengono una catena di
trasporto elettronico capace di utilizzare il NADH
prodotto
Per assicurare l’apporto di NAD+ (ossidato), per
l’ossidazione continua degli acidi grassi, il NADH
(ridotto) è trasportato nel citosol in cambio di
NAD+
La situazione è diversa tra cellule animali e
vegetali
a)Negli ANIMALI l’ossidazione perossisomica dà
un significativo contributo al processo totale di
ossidazione degli acidi grassi: nel fegato si può
raggiungere il 50% del totale degli acidi grassi
ossidati.
Comunque,
nei
perossisomi,
l’ossidazione degli acidi grassi è parziale
Questi
organelli
difatti
sembrano
essere
particolarmente importanti nella conversione degli
acidi grassi a lunga catena in acidi grassi a media
catena che poi vengono trasportati, tramite lo
shuttle della carnitina, nei mitocondri, dove il
processo di –ossidazione si completa
L’acetil-CoA prodotto nei perossisomi viene anche
trasportato nella matrice mitocondriale dove può
essere ossidato dal ciclo TCA o convertito in corpi
chetonici. Il movimento di acetato tra gli organelli
subcellulari si basa sempre su meccanismi
mediati dalla carnitina
• Nelle piante, i gliossisomi nei semi ed i perossisomi nelle
foglie sono i principali siti di –ossidazione. Infatti, si pensa
che
la
–ossidazione
mitocondriale
contribuisca
marginalmente all’ossidazione degli acidi grassi nelle piante,
mentre è sicuro che sia i gliossisomi che i perossisomi sono
capaci di ossidare completamente gli acidi grassi
In alcuni semi germinanti, la quantità di acidi grassi mobilitata
dai grassi di riserva e ossidata attraverso i gliossisomi è
enorme, specialmente nelle oleaginose. Questi speciali
organelli contengono sia enzimi necessari per ossidare gli
acidi in acetil-CoA, sia gli enzimi del ciclo del gliossilato,
l’isocitrato liasi e la malato sintasi che permettono alle piante
di convertire l’acetil-CoA in glucosio
-ossidazione
Come suggerisce lo stesso nome, questo
processo determina l’ossidazione degli acidi
grassi a livello del carbonio (carbonio 2). Si può
avere la rimozione di un singolo atomo di carbonio
dal gruppo carbossilico o la produzione di acidi
grassi -idrossilici
(a) Rimozione
COOH
del
gruppo
(b) Produzione di un -idrossi acido
grasso
Il significato dell’-ossidazione nei tessuti animali è ancora
sconosciuto. Ci sono tre campi in cui potrebbe giocare un ruolo:
Primo, potrebbe essere un meccanismo attraverso il quale sono
sintetizzati gli acidi grassi a catena dispari, ad esempio la
rimozione di un atomo di carbonio dal gruppo carbossilico di un
acido grasso a catena pari
Secondo, potrebbe essere coinvolta nella sintesi degli acidi grassi
-idrossilici trovati in certi tessuti, in particolare nei tessuti
cerebrali dove gli sfingolipidi contengono una grande quantità di
questo tipo di acidi grassi
Terzo, potrebbe agire di concerto con la -ossidazione per facilitare
l’ossidazione di acidi grassi che hanno caratteristiche strutturali
che ne impedirebbero l’ossidazione tramite la sola -ossidazione
Anche nelle piante l’importanza della ossidazione non è chiara. Si è tuttavia dimostrato
che, con l’eccezione dei semi germinanti dove la
-ossidazione è estremamente attiva, l’ossidazione potrebbe essere il percorso più
importante per l’ossidazione degli acidi grassi
Oltre al suo ruolo nella degradazione degli acidi
grassi, l’-ossidazione è certamente coinvolta
nella produzione degli alcooli a lunga catena e
degli idrocarburi trovati nella cutina e suberina
che compongono la cuticola
-ossidazione
In questo processo, gli acidi grassi vanno incontro
all’ossidazione del carbonio o metilico
trasformando acidi dicarbossilici e acidi grassi idrossilici
Negli animali, l’enzima responsabile sembra
essere un’ossidasi con funzioni miste, associata
al reticolo endoplasmatico e probabilmente
coinvolgente un citocromo specializzato (P450).
Nelle piante il coinvolgimento del citocromo è
dubbio
La -ossidazione può essere un passo essenziale
nell’ossidazione degli acidi grassi là dove il
gruppo carbossilico sia indisponibile per la ossidazione; infatti, la produzione di un gruppo
carbossile dal carbonio metilico di un acido
grasso permette al processo della -ossidazione
di partire dalla parte opposta della molecola
La produzione di un acido grasso -idrossilico
può avere un ruolo anche nella formazione della
cutina e della suberina