APOPTOSI E NECROSI
Per necrosi si intende la morte cellulare accidentale, che coinvolge contemporaneamente gruppi più
o meno estesi di cellule, facenti parte di un tessuto o di un organo. Le cause sono di vario tipo:
•Traumi
•Anossia
•Ischemia
•Ipertermia
•Esposizione al calore
•Azione di veleni o tossine
A seconda della natura dell’agente lesivo, la necrosi interviene:
•per massicce variazioni osmotiche,
•per arresto dell’apporto di ossigeno o di sostanze nutritive,
•per denaturazione di proteine (strutturali ed enzimatiche)
Tuttavia la necrosi può essere preceduta da un danno subletale quando l’intensità degli stimoli è al
di sotto di una certa soglia, che varia da tessuto a tessuto e che a sua volta può evolvere in tre
direzioni:
adattamento,
necrosi
apoptosi
ADATTAMENTO
Entro certi limiti, eccessive stimolazioni fisiologiche o alcuni stimoli patologici
possono innescare varie forme di adattamento cellulare, morfologico o
fisiologico/funzionale, grazie alle quali le cellule acquisiscono nuove caratteristiche
che, seppure anomale, ne preservano la vitalità e ne modulano la funzione ad un dato stimolo.
SE i limiti di una risposta adattativa vengono superati oppure
se in qualche modo l’adattamento non è possibile
si verificano una serie di eventi genericamente chiamati
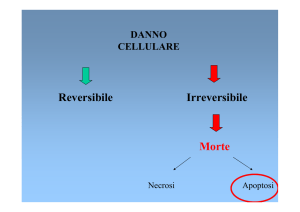
DANNO CELLULARE.
Il danno cellulare è reversibile fino ad un certo punto, ma se lo stimolo persiste oppure è grave fin
dall’inizio la cellula raggiunge un punto di non ritorno e va incontro al danno cellulare irreversibile
e alla morte cellulare.
RISPOSTE CELLULARI AL DANNO
A.Danno subletaleadattamento cellularedi tipo variomorfologico e fisiologico:
•adattamenti metabolici
•adattamenti strutturaliriguardano modificazioni della crescita, delle dimensioni e del
differenziamento cellulare;
•iperplasia:aumento del numero delle cellule in un dato tessuto o in un organo
•ipertrofia: aumento delle dimensioni delle cellule(ipoplasia)
•atrofia(ipotrofia): diminuzione delle dimensioni e della funzione cellulare
•metaplasia: alterazione del differenziamento cellulare;Risposte adattative reversibili: cessano dopo
la rimozione dello stimolo.B. Danno letaleNECROSI O APOPTOSI
Le cellule necrotiche esplodono, riversando il loro contenuto sulle cellule adiacenti e scatenando
una risposta flogistica a carico dell’area occupata dai detriti delle cellule morte. In seguito subentra
il richiamo chemiotattico dei leucociti che ha come scopo un duplice effetto: da un lato la fagocitosi
dei detriti, dall’altro il rilascio da parte di esse di enzimi lisosomiali che agiscono sulle cellule
morte, facilitandone la dissoluzione.
Perdita della capacità omeostatica: assorbimento di acqua e ioni,aumento delle dimensioni cellulari
e mitocondriali, dissoluzione nucleare.Rottura della membrana plasmatica,rilascio di enzimi
proteoliticiche provocano Infiammazione.
Meccanismi di azione del danno
intracellulari particolarmente
vulnerabili:integrità delle membrane cellulari; respirazione aerobia; sintesi proteica; integrità
correlate che, indipendentemente dal preciso punto di attacco, il danno in una sede porta ad un
Meccanismo di azione del dannoPer alcuni agenti dannosi i siti biochimici di attacco sono ben
definiti (tossine batteriche, cianuro inibisce la citocromo ossidasi) ma in generale le cause precise
che portano a morte cellulare non sono conosciute. In generale si può dire che la glicolisi, il ciclo
dell’acido citrico e la fosforilazione ossidativa sono particolarmente vulnerabili. Indipendentemente
dall’agente scatenante le maggiori cause di morte sono:A.deplezionedi ATP(prodotto dalla
fosforilazione ossidativa e dalla glicolisi) e la diminuita sintesi di ATP sono conseguenze comuni
del danno ischemico e tossico;B.ossigeno e radicali liberi derivati dall’ossigeno; questi si formano
durante la riduzione dell’ossigeno molecolare ad acqua;C.calcio intracellularee
dei mitocondri e reticoloendoplasmaticodaATPasiassociate alla membrana. Interruzione di ATP o
aumento non specifico dellapermiabiltàdi membrana fanno aumentareCa intracellulare.Qsattiva
numerosi enzimi (fosfolipasi; proteasi,ATPasi,endonucleasi)D.Difetti della permeabilità di
membranaE.
I meccanismi d'azione con cui le varie cause di necrosi cellulare agiscono sulla cellula possono
essere diversi, ma sono sostanzialmente riconducibili a danni diretti sulle strutture di membrana o
alterazioni degli enzimi coinvolti nel metabolismo energetico, che si riflettono sulla permeabilità
selettiva della membrana cellulare.
•Si verifica entrata di Na+e fuoriuscita di K+. La ridistribuzione cationica è accompagnata da
spostamento di liquidi e rigonfiamento cellulare
•Il mancato funzionamento delle pompe cationiche o i danni diretti sulla membrana provocano un
aumento soprattutto di Ca+ +intracellulare che entra nella cellula mosso da un alto gradiente di
concentrazione.
•L'aumento del Ca+ +citosolico attiva le fosfolipasi di membrana che degradano i fosfolipidi della
membrana aumentando il danno cellulare.
•Le idrolasi rilasciate dai lisosomi(autolisi) producono una rapida accelerazione della distruzione
cellulare
•Il DNA viene esposto all'azione delle desossiribonucleasi lisosomiali dalla digestione proteolitica
degli istoni
APOPTOSI
Il concetto che le cellule morissero durante il loro sviluppo naturale era già stato ipotizzato più di
200 anni fa. Già nel XVII secolo Harveyaveva osservato il rimodellamento del cuore di un
embrione mentre nel XVIII secolo R.Hookaveva constatato che alcune cellule del tronco della
quercia erano sottoposte a quello che lui interpretò essere una sorta di morte fisiologica. Il primo
scienziato ad osservare e descrivere la morte cellulare nel sistema nervoso di embrioni di rospo fu
Carl Vogtnel XIX secolo (1842). Ancora nello stesso secolo (anno 1889) va collocata una pietra
miliare delle scoperte che hanno indirizzato verso il concetto di morte programmata. In quell’anno
August Weismannpubblicò infatti un trattato sull’eredità si cominciava a prendere atto che
l’organismo contenesse un’ informazione che ne programmava il tempo di vita più lungo rispetto ad
altre. Il concetto di morte cellulare naturale fu riscoperto intorno agli anni ’50 con le scoperte di
Saunders(1948) che mostrò che esisteva un tipo di morte naturale scandito da stimoli intrinseci e
precisi tempo-dipendenti o di Rita Levi Montalcini e Hamburgernel 1949 che avrebbero portato
alla consapevolezza che molti neuroni vengono persi nel corso della normale neurogenesi, o ancora
si ricordi l’importantissima pubblicazione di Glucksmann nel 1951 che sottolineava l’esistenza di
un processo di eliminazione naturale di strutture non più utili da parte degli organismi.
Nel 1965 Lockshin e Williams, lavorando sulla metamorfosi degli insetti, estesero il concetto e
introdussero il termine “morte programmata”. Intorno al 1977 J. Sulstone H.R.Horvitzfornirono
una prima comprensione del fenomeno dal punto di vista genetico dallo studio del nematode
C.Elegansche inizialmente genera 1090 cellule somatiche per poi perderne 131 durante lo sviluppo.
Nel 1988 David Vouxe colleghi identificarono un componente fondamentale del sistema (Bcl-2)
negli invertebrati. La biologia di questo gene venne studiata a fondo da Stanley Korsmeyere
colleghi. Il primo elemento di riconoscimento del fenomeno apoptotico che non avesse a che fare
con la morfologia si presentò di pari passo con l’osservazione che la morte cellulare fosse
accompagnata solitamente dalla rapida attivazione di endonucleasi(H.Wyllie, 1980). Confronti tra
aspetti morfologici e anatomici di vertebrati e invertebrati erano stati fatti fin dal 1969, ma solo nel
1992 venne reso noto che il gene Bcl-2umano poteva inibire la morte cellulare nel nematode. Si
ricordano inoltre le scoperte di Yuan del 1993 e di Newmeyere Reeddel 1994 i quali scoprirono che
l’esecuzione dell’apoptosi è mediata dai mitocondri. Nel 1999 Lodish, Desaghere
Martinouparlarono di come l’apoptosi poteva essere innescata dall’estrusione del citocromo c nel
citoplasma. Recentemente le conoscenze riguardo alla morte cellulare si sono moltiplicate e più il
campo di ricerca si estende, prima verranno proposti nuovi agenti terapeutici capaci di trattare
quelle malattie legate ad un inappropriato controllo del complesso fenomeno della morte cellulare
programmata.
La parola deriva dal greco -da , -cadere giù fu utilizzato per la prima volta dal Prof.
James Cormack(Dipartimento di Greco, Università di Aberdeen).
A differenza della necrosi l’apoptosi è un processo attivo, durante il quale la cellula stessa accende
uno specifico programma che ne determina la morte, una sorta di suicidio.
Il processo apoptotico, sebbene abbia nei vari modelli caratteristiche comuni, sicuramente non é un
fenomeno univoco, può essere spontaneo o indotto da agenti di diverso genere.
Nel corso dell’apoptosi:
il DNA si frammenta
il citoscheletro collassa
la cromatina si condensa
la cellula si contrae formando una
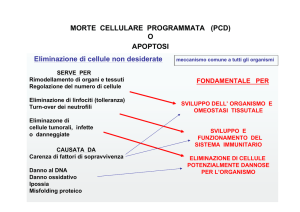
Importante normalmente in processi fisiologici e di sviluppoperché elimina cellule non
funzionali o strutturalmente inutiliinterviene:
•NELLO SVILUPPO: maturazione sistema immune, morfogenesi, embriogenesi
•NELL’ADULTO: DNA danneggiato e riparazione delle ferite
serie di bolle (BLEBS) che, separandosi dalla cellula, formano i corpi apoptoci, i quali
vengono fagocitati da circolanti professionali o da cellule epiteliali contigue (fagociti
occasionali)
Aspetti morfologico funzionali dell'apoptosi
Modificazione delle dimensioni cellulari: la condensazione citoplasmatica provoca una
contrazione cellulare.
Modificazioni della membrana plasmatica: cambia la distribuzione dei fosfolipidi di membrana.
Durante lo stimolo apoptotico la fosfatidilserina(PS) trasloca dalla parte citoplasmatica alla parte
extracellulare della membrana plasmatica.
Distruzione della fisiologia mitocondriale: depolarizzazione del potenziale di membrana
mitocondriale e alterazione della permeabilità della membrana mitocondriale esterna con
conseguente rilascio di citocromo-cnel citosol.
Modificazioni citoplasmatiche: il rilascio di citocromo-c dai mitocondri provoca l’attivazione
delle caspasi in grado di tagliare diversi substrati citoplasmatici e nucleari.
Modificazioni nucleari: la firma biochimica dell’apoptosi è la regolare frammentazione del DNA
dovuta al taglio fra le unità nucleosomali.
Famiglia di proteasi che hanno una cisteina (c-) nel sito attivo etagliano proteine bersaglio a livello
di residui di acido aspartico (-asp-).
Sono formate da due subunità che formano un eterodimero e due di questi dimeri si uniscono per
formare il tetramero attivo.
Le caspasi attive generano reazione a catena:CASCATA DELLE CASPASICASPASI
INIZIATRICI:pro-caspasi che operano all’inizio, sono attivate da:-stimoli extracellulari (capsasi-8
e caspasi-10)-rilascio citocromo c nel citosol (caspasi-9)CASPASI EFFETTRICI:Caspasi-3,-6,-7:
sono responsabili della rottura proteolitica di importanti substrati citoplasmatici e nucleari
Si può dire che l’apoptosi abbia inizio all’attivazione delle caspasi in conseguenza a segnali di
diverso tipo che indicano tutti la necessità che una determinata cellula venga eliminata per apoptosi.
L’attivazione delle caspasi è resa possibile dall’innesco di una cascata di eventi molecolari che
possono essere indotti in modi differenti.
Le fasi in cui può essere suddivisa l’apoptosi sono quindi due:
e le caspasi agiscono per causare la vera e propria morte cellulare.
L’attivazione delle caspasi è dovuta a due meccanismi che di per sé sono separati, ma che
convergono entrambi al medesimo scopo. La due vie possibili sono dette:
Via estrinseca e via intrinseca
Alla base di esse vi sono fenomeni molecolari profondamente diversi, ma non si deve dimenticare
che le due vie possono interconnettersi a più livelli.
La via estrinseca si basa su un sistema classico di interazione recettore-ligando che, una volta
attivato, determina l’interazione di più pro-caspasi con alcune proteine attivatrici. Questi recettori di
morte appartengono ad una famiglia genica che comprende recettori simili al TNF-R (Recettore per
il Tumor Necrosis Factor) tra i quali il più studiato in termini di segnalazione apoptotica è il Fas.
Il Fas è costituito da:
•un dominio extracellulare interazione con ligando specifico
•un dominio trans-membrana
•dominio di morte (DD) regione citoplasmatica organizzata in a-eliche
Il Fas è presente sulla membrana plasmatica di tutte le cellule, mentre il suo ligando, FasL, è
espresso solo da tipi cellulari specifici.
FasL induce l’aggregazione del recettore, successivamente la proteina FADD si lega al dominio di
morte del recettore mediando il reclutamento del proenzima Caspasi-8 presso il recettore.
L’avvicinamento di più Pro-Caspasi-8 stimola la dimerizzazione di due pro-enzimi con il
conseguente cambio conformazionale richiesto per l’innesco dell’attività proteolitica.
La via intrinseca è sotto il diretto controllo del mitocondrio e viene innescata in seguito al rilascio
di specifici componenti mitocondriali.
La caspasi regolativa è la Caspasi-9, che contiene un pro-dominio regolativo contraddistinto da una
serie di a-eliche denominato CARD (CaspaseActivationand RecruitmentDomine), il quale media le
interazioni proteina-proteina riconoscendo un altro dominio CARD presente su altre proteine.
Nell’uomo, il gene che rappresenta il punto cruciale della cascata che attiva le caspasi è Apaf1(ApoptoticProteaseActivatingFactor-1), che contiene un dominio CARD, che può interagire con il
CARD della caspasi-9.
L’evento che promuove il legame tra Apaf-1e caspasi-9 consiste nell’arrivo nel citoplasma di un
terzo componente, il citocromo c normalmente presente nel mitocondrio, che determina un
cambiamento conformazionale in Apaf-1consentendo il riconoscimento e l’interazione del suo
dominio CARD con quello della caspasi-9; si forma così una ruota proteolitica detta apoptosoma,
formata da sette dimeri di caspasi-9 che interagiscono con sette monomeridi Apaf-1. Il meccanismo
di funzionamento di questa via si basa sull’equilibrio che si viene a stabilire tra la presenza di
molecole pro-eantiapoptoticheche regolano la permeabilità mitocondriale e il rilascio di induttori di
morte sequestrati all’interno dei mitocondri
REGOLAZIONE DELLA VIA INTRINSECA
Le proteine Bcl2 dei mammiferi regolano la via intrinseca dell’apoptosi controllando il rilascio del
citocromo c e di altre proteine mitocondriali. Le Bcl2si dividono in:
BH123(Baxe Bak) e le BH3;
gruppo appartiene Blc2stessa e BclXL (prodotto dello splicingalternativo diBclX);
La fase terminale dell’apoptosi è mediata da una cascata proteolitica verso cui convergono i vari
meccanismi di innesco. Le caspasi di inizio infatti mediano l’attivazione delle caspasi effettrici, che
una volta attivate clivano le proteine citoscheletriche e quelle della matrice nucleare, distruggendo
quindi il citoscheletro e portando alla frammentazione del nucleo.
Nel nucleo i bersagli dell’attivazione delle caspasi comprendono proteine coinvolte nella
trascrizione, nella replicazione e nella riparazione del DNA. In particolare, l’attivazione della
caspasi 3attiva una DNasi citoplasmatica (CAD), clivando un inibitore dell’enzima (ICAD).
Nel caso dell’apoptosi mediata dai linfociti T citotossici (CTL), è possibile saltare le fasi di inizio e
passare subito alla fase effettrice. Questi infatti, una volta riconosciuta la cellula bersaglio,
attraverso specifici segnali da questa appositamente esposti sulla membrana, secernono perforina.
Questi contengono una serina proteasi chiamata granzima B che fa le veci di una caspasi effettrice e
taglia le proteine a livello dei residui di acido aspartico attivando così una grande quantità di
caspasi. In definitiva i CTL uccidono la cellula bersaglio passando direttamente alla fase effettrice
dell’apoptosi.
Non appena la cellula intraprende la via dell’apoptosi, essa secerne anche dei fattori solubili in
grado di reclutare i fagociti. Il reclutamento dei fagociti consente una rapida eliminazione delle
cellule apoptotiche, prima che esse vadano incontro al rilascio del loro contenuto cellulare in modo
da evitare una conseguente risposta infiammatoria.
E’ importante comunque ricordare che i fagociti si comportano differentemente nei confronti delle
cellule apoptotiche di quanto non facciano rispetto ai batteri per esempio, cercando di minimizzare
la produzione di citochine pro-infiammatorie. Il riconoscimento della cellula apoptotica da parte del
fagocita è facilitata da molecole marcatrici che le cellule stesse espongono sulla loro membrana
permettendo il riconoscimento. Le cellule apoptotiche possono inoltre essere opsonizzate per la
fagocitosi attraverso il legame di sostanze secrete appositamente dai fagociti stessi. Il processo
fagocitario è così efficiente che le cellule morte scompaiono senza lasciare la minima traccia e
l’infiammazione è praticamente assente.
Gli inibitori dell’apoptosi (IAP) sono stati identificati per la prima volta in certi virus di insetto,
codificano proteine che impediscono alle cellule del virus di suicidarsi per apoptosi.
Tutti gli IAP hanno uno o più domini BIP (baculovirus IAP repeat)che permettono loro di legarsi
alle caspasi attivate e di inibirle, stabiliscono infatti una soglia inibitrice che le caspasi attivate
devono superare per scatenare l’apoptosi.
In Drosophilaquesta barriera inibitrice può essere neutralizzata da proteine anti-IAP, che sono
prodotte in risposta a vari stimoli apoptotici.
L’apoptosi interviene in processi fisiologici
L’apoptosi ha un ruolo complementare, ma opposto a quello della mitosi e della proliferazione
cellulare nella regolazione delle popolazioni cellulari. Si stima che per mantenere l’omeostasi in un
corpo umano adulto sia necessaria la sintesi di circa 10 miliardi di nuove cellule ogni giorno per
bilanciare la perdita di quelle che muoiono per apoptosi. Questo numero può inoltre crescere
significativamente quando si ha un aumento di apoptosi durante il normale sviluppo e
invecchiamento o in caso di malattia. Da ricordare che un ruolo molto importante nel mantenimento
dell’omeostasi numerica è svolto dai fattori di crescita, che hanno sia la funzione di indurre le
cellule bersaglio alla proliferazione, sia quella di svolgere il ruolo di “fattori di sopravvivenza”.
Tra cui l’impianto dell’ovocita, l’organogenesi, l’involuzione di strutture durante lo sviluppo e la
metamorfosi. Ne è un esempio la perdita delle membrane interdigitali e l’apertura della rima
palpebrale durante lo sviluppo del sistema nervoso e immunitario.
Distruzione delle cellule endometriali durante il ciclo mestruale, l’atresia dei follicoli ovarici nella
menopausa, la regressione della mammella dopo lo svezzamento.
•NELL’ELIMINAZIONE DELLE CELLULE CHE PERDONO ACCIDENTALMENTE
CONTATTO CON IL TESSUTO D’ORIGINE(ANOIKIS):Si tratta del programma apoptotico
che viene attuato quando la cellula perde contatto con le cellule circostanti. E’ una prima forma di
protezione degli organismi multicellulari nei confronti della possibilità che delle cellule vadano in
sedi improprie.
•MORTE DELLE CELLULE DELL’OSPITE CHE HANNO ESAURITO LA LORO
UTILITA’:E’ questo il caso dei neutrofili durante la risposta infiammatoria acuta e i linfociti al
temine di una risposta immunitaria. In queste situazioni le cellule vanno incontro ad apoptosi in
quanto private dei necessari segnali di sopravvivenza.
•NEL DANNO CELLULARE O NELL’INFEZIONE:Molte sostanze tossiche, farmaci, radicali
liberi dell’ossigeno e radiazioni ionizzanti causano danni al DNA o gravi stress al reticolo
endoplasmatico o ai mitocondri: tutte queste condizioni, qualora superino una certa soglia, possono
indurre la morte per apoptosi. L’apoptosi può essere determinata anche dall’accumulo di proteine
non correttamente folded, nel qual caso si ha stress del reticolo endoplasmatico, evento scatenante
la morte cellulare. Le infezioni virali fanno anch’esse scattare nella cellula un meccanismo a livello
del sistema immunitario che tende a indurre apoptosi, anche in questo caso per difendere
l’organismo intero dalla propagazione virale. A livello del sistema immunitario l’apoptosi è in
particolare indotta dai linfociti citotossici, proprio come difesa contro virus e tumori per eliminare
le cellule infettate e quelle neoplastiche (lo stesso meccanismo è responsabile del rigetto cellulare
nei trapianti). DIFETTO APOPTOSINon consente un turn-over sufficientemente veloce delle
cellule anormali, che non possono perciò venire adeguatamente sostituite in quanto la loro
sopravvivenza è prolungata.
CANCRO
tumori con mutazioni di p53 oneoplasie ormonodipendenti(es. carcinomi di mammella, prostata e
ovaia)
MALATTIE AUTOIMMUNIche insorgono in seguito alla mancata eliminazione dei linfociti
autoreattividopo l’incontro con gli antigeni self.
DIABETE DITIPO I
AUMENTO APOPTOSI
In queste malattie si ha una perdita notevole di cellule normali o di cellule con funzione difensiva.
MALATTIE NEURODEGENERATIVE
•Atrofia Muscolo Spinale (SMA)
•Sclerosi Laterale Amiotrofica(SLA)
•Malattia di Alzheimer
•Malattia di Parkinson
DANNO ISCHEMICO
MORTE DELLE CELLULEINFETTATE DA VIRUS
DIFETTO APOPTOSICANCROCELLULE CANCRO DISATTIVAZIONE PUNTI CELLULE
DANNEGGIATECONTROLLO CICLO CELLULARE EVITANO APOPTOSIINTERFERONI –a
e –b INDUCONO TRASCRIZIONE GENE p53AUMENTO PROTEINE p53ONCOSOPPRESSORI
•L. Yang ha riportato in un articolo del 2003, il risultato del lavoro svolto riguardo il segnale di
morte difettoso in un tipo di cancro delle cellule polmonari (NCI-H460): La proteina
XIAP(inibitrice dell'apoptosi X-linked) è sovra-espressa nelle cellule H460.Le XIAP legano la
forma attivata della caspasi-9, e sopprimono l'attività dell'attivatore apoptotico citocromo c. La via
apoptotica è stata trovata altamente ripristinata nelle cellule H460 che presentavano un peptide
Smacche lega le IAP (proteine inibitrici l'apoptosi).
•La sovraespressionedell'inibitore di apoptosi Bcl-2è frequente nel linfoma follicolare.
•Un interessante caso di riutilizzo dei prodotti dell'apoptosi è stato presentato da M. L. Albert in
un articolo: egli descrive come le cellule dendritiche fagocitino le cellule tumorali apoptotiche;
dopo la maturazione, queste cellule dendritiche presentano l'antigene ai linfociti T killer, che poi
diventano specifici per distruggere le cellule che stanno subendo una trasformazione maligna.
Questa via apoptosi-dipendenteper l'attivazione dei linfociti T non è presente durante la necrosi ed
ha aperto interessanti possibilità nella ricerca sull'immunità tumorale.
DIFETTO APOPTOSIMALATTIE AUTOIMMUNIL'apoptosi può giocare un ruolo importante
nelle malattie autoimmuni sotto due diversi aspetti:
•DIFETTI REALIZZAZIONE APOPTOSI:persistenza e proliferazione di linfociti
autoreattiviche possono scatenare una risposta di natura autoimmune e contemporaneamente dar
vita a linfoproliferazionenon maligna.
•ECCESSIVA MORTALITA’ DIUN TESSUTO:manifestazioni di ipersensibilità secondarie e
determinate da uno stato di infiammazione cronica.Diabete di tipo I:difetti nel funzionamento di
Fas a carico delle isole pancreatiche.Mutazioni genetiche come causa di difetti apoptotici nel
sistema immunitario ha finora ottenuto risultati positivi solo per quanto riguarda il sistema Fas/FasL.Sia nei modelli murini che nell'uomo tali mutazioni sembrano essere associate prevalentemente a
sindromi linfoproliferativedi tipo autoimmune.Sebbene ciò indichi una stretta correlazione tra Fas e
autoimmunità la ricerca di mutazioni nei pazienti non ha finora fornito i risultati attesi.La stessa
linfoproliferazionenon è necessariamente legata all'insorgenza delle malattie autoimmuni.Sebbene
la perdita della tolleranza sia considerata la principale causa di tali patologie i dati in nostro
possesso ci permettono di formulare ipotesi solo sulla base di fattori di rischio potenziali
ECCESSO DIAPOPTOSIMALATTIE NEURODEGENERATIVELa graduale perdita di
neuroni in diverse parti del cervello caratterizza gran parte delle patologie neurodegenerative. Dato
che il sistema nervoso centrale (SNC) durante lo sviluppo è sede di un’intensa attività apoptotica e
che nel periodo adulto la sopravvivenza cellulare dipende dall’espressione di geni
antiapoptoticiquali Bcl-xLsi è ipotizzato che esso sia un tessuto particolarmente suscettibile ai
difetti dei pathwayapoptotici. Tale ipotesi è avvalorata da numerose evidenze sperimentali che
mostrano un coinvolgimento dell’apoptosi nello sviluppo di diverse malattie dell’SNC.
•ATROFIA MUSCOLO SPINALE (SMA) :degenerazione dei motoneuroni spinali causata da
delezionedel gene SMA che normalmente promuove l’attività della proteina antiapoptoticaBcl-2;
•MALATTIA DIALZHEIMERè associata a diverse mutazioni in geni che codificano per proteine
coinvolte più o meno direttamente nel processo apoptotico. Come la presenilina2, che sembra agire
a valle di Fas. Se la presenilina2 è mutata questa inibizione non avviene e FasL viene
iperespressoinducendo l’apoptosi nei neuroni dopaminergici. Il coinvolgimento del processo
apoptotico nella malattia di Alzheimer è stata osservata anche a livello delle placche beta amiloidi,
aggregati proteici insolubili caratteristici della patologia che, accumulandosi nel tessuto cerebrale,
causano la necrosi di intere aree del cervello. In linee cellulari neuronali si è infatti osservato che i
peptidi delle placche beta amiloidi sono in grado di inibire la trascrizione dei geni
antiapoptotici(Bcl-2) e di stimolare invece quella di geni proapoptotici(Bax), rendendo i neuroni più
suscettibili alla morte, soprattutto in risposta allo stress ossidativo.
•SCLEROSI LATERALE AMIOTROFICA (SLA):progressiva perdita di motoneuroni spinali. Il
fatto che proteine antiapoptotichepossano rallentare la malattia in modelli animali suggerisce un
importante ruolo dell’apoptosi;
•MALATTIA DIPARKINSON: ruolo dell’apoptosi nel determinare la progressiva perdita dei
neuroni dopaminergicinigrostriatali. In particolare si è scoperto che la selegilina, farmaco storico
utilizzato nel trattamento della malattia esercita il suo effetto neuroprotettivoalterando l’espressione,
tra gli altri, dei geni antiapoptoticiBcl-2 e Bcl-xL.•ISCHEMIA CEREBRALE: Sebbene
nell’ischemia grave il cuore della lesione sia prevalentemente formato da tessuto necrotico, nelle
zone circostanti prevale il processo apoptotico, accompagnata da iperespressionedelle caspasi,
soprattutto della caspasi 3. Facendo iperesprimereagli animali proteine antiapoptoticheo inibitori
specifici di caspasi si è osservato un danno cerebrale più ridotto dopo induzione di ischemia.
• 11 Caspasi nell’uomo - 6 apoptotiche3 Caspasi
iniziatrici (Casp-8, -9, -10) hanno grossi domini
di attivazione (129-219 aa.), contengono DED
e CARD per il legame agli adattatoriattivatori
e per il reclutamento
• 3 Caspasi effettrici (Casp-3, -6, -7) svolgono
compiti esecutivi nell’apoptosi vera e propria,
degradano proteine ...
• 5 Caspasi coinvolte nell’attivazione di
citochine (interleukin converting enzymes ...)
I substrati delle caspasi
• Proteine del citoscheletro
• Proteine del nucelar envelope
• Fattori di trascrizione
• Proteine della regolazioen del ciclo
cellulare
• Proteine della trasduzione del segnale
• Replicazione del DNA
• Circa 60 targets diversi
Approfondimento
L’apoptosi è la morte programmata cellulare ed è un meccanismo intrinseco di
autodistruzione della cellula. Si tratta di un fenomeno molto importante durante
l’embriogenesi (es. nella regressione delle pliche interdigitali); nella metamorfosi e
durante la vita dell’adulto (es. per il mantenimento dell’omeostasi numerica tissutale;
nella selezione clonale negativa dei linfociti T autoreattivi; in presenza di atrofia
dovuta ad assenza di stimolazione; nella distruzione di quelle cellule il cui DNA
risulta danneggiato o mutato). L’apoptosi, la cui prima descrizione morfologica risale
al 1842 da parte di Vogt, ha inizio con la perdita dei microvilli e delle giunzioni
intercellulari, la riduzione del volume cellulare e alterazioni morfo-funzionali del
nucleo (addossamento della cromatina) e termina con la frammentazione della cellula
in corpi apoptotici. Si tratta di un processo attivo che richiede energia, trascrizione
genica e sintesi proteica e risulta dal bilancio tra i segnali di morte e di
sopravvivenza. Per quanto riguarda le caratteristiche biochimiche, la morte
programmata è caratterizzata dall’esternalizzazione della fosfatidilserina ad opera
della flippasi e dall’attivazione di enzimi come le caspasi, cistein-proteasi iniziatrici
ed effettrici, attivate in seguito ad auto-clivaggio e responsabili, a loro volta, del
clivaggio di substrati a valle; le transglutaminasi, che creano legami molto forti tra
le proteine per formare uno scaffold protettivo nella membrana cellulare per la
resistenza ad agenti litici; le endonucleasi, che degradano la cromatina in frammenti
caratteristici tutti multipli di 200 pb (ladder pattern).
L’apoptosi può essere innescata tramite due vie: la via estrinseca o recettoriale e la
via intrinseca o mitocondriale. La prima via passa attraverso l’attivazione di
specifici recettori (Fas, CD95, Killer/DR5), i cui domini di morte (death domain o
DD) nella porzione intracellulare sono in grado di reclutare specifiche molecole
adattatrici, come FADD, i cui domini effettori di morte (death effector domain o
DED) sono, a loro volta, in grado di reclutare i DED domain di specifiche caspasi
iniziatrici come la caspasi 8: a questo punto, due caspasi 8 vengono a trovarsi vicine e
si auto-clivano secondo la logica trans e danno origine a un tetramero attivo, con
attività cistein-proteasica nei confronti di caspasi effettrici e di altri specifici substrati
a valle (Fig 1B). La via intrinseca, invece, è innescata, da una parte, tramite
l’attivazione di specifiche proteine pro-apoptotiche come Bid, Bad, Bak e Bax e
dall’altra, tramite l’inattivazione di proteine anti-apoptotiche come Bcl-2 e Bcl-XL. Il
bilancio tra le prime e le seconde proteine determina il prevalere dell’apoptosi o della
sopravvivenza cellulare. Quando nella cellula prevalgono le proteine pro-apoptotiche,
si ha un aumento della permeabilità mitocondriale e il conseguente rilascio del
citocromo c, che attiva e fa assumere una conformazione distesa ad Apaf-1, i cui
CARD domain sono in grado di interagire con i CARD domain di caspasi iniziatrici,
come la caspasi 9 (Fig 1A).
Fig.1
Le due vie apoptotiche sono strettamente correlate e dipendenti l’una dall’altra:
l’attivazione della via estrinseca, e quindi della caspasi 8, porta al clivaggio della
proteina pro-apoptotica Bid, dando origine alla forma tronca tBid, che agisce
direttamente a livello mitocondriale sulle proteine Bax e Bak, inducendo la loro
oligomerizzazione. Gli oligomeri di Bax e Bak, che si sono così formati, danno
origine alla formazione di veri e propri pori sulla membrana esterna del mitocondrio,
favorendo l’aumento della permeabilità mitocondriale e il conseguente rilascio del
citocromo c, dell’endonucleasi G, di Smac/Diablo (proteina che lega e inibisce le
XIAP, cioè le proteine che inbiscono l’apoptosi), di AIF (il fattore che induce
apoptosi) e di altri fattori apoptotici (Fig.2).
Fig.2
P53 E L’APOPTOSI:
p53 è un oncosoppressore essenziale per prevenire una proliferazione inappropriata
delle cellule e per mantenere l’integrità del genoma in seguito all’attivazione di
specifici oncogeni o in seguito a vari stimoli di stress genotossici. I livelli di p53 sono
tenuti bassi dal suo normale turnover di proteolisi da parte dell’ubiquitina ligasi
MDM2. Una volta attivata, p53 può indurre, a seconda delle circostanze, l’arresto del
ciclo cellulare o l’apoptosi. Ma da cosa dipenda l’attivazione dell’uno o dell’altro
fenomeno non è ancora chiaro. Ci sono diversi modelli che danno possibili
spiegazioni riguardo l’azione di p53 nell’induzione di questi fenomeni; due di questi
modelli sono: “p53 dumb” e “p53 smart” (K.H.Vousden, Cell 2000). Secondo il
primo modello, l’induzione da parte di p53 dei geni per l’arresto del ciclo cellulare e
dei geni apoptotici è la stessa in tutte le condizioni. In base al modello “p53 smart”,
invece, l’espressione differenziale delle due categorie di geni sarebbe causata da p53
stessa: in particolare, è stato visto che alti livelli di p53 attivano preferenzialmente i
geni apoptotici, i cui promotori, infatti, sono legati da p53 a bassa affinità. Inoltre,
modificazioni post-traduzionali di p53, ad esempio, la fosforilazione sul residuo di
serina 46, favorirebbero la trascrizione dei geni apoptotici: questo sarebbe dovuto al
fatto che tali modificazioni alterano la specificità di legame di p53 ai promotori
apoptotici e a co-attivatori specifici per l’apoptosi (Fig.3).
Fig.3
Tra i co-attivatori specifici per l’apoptosi vi sono: ASPP1, p63, p73 e STAT1; perché
le cellule vadano in apoptosi, p21, ossia l’inibitore delle chinasi, cicline-dipendenti,
deve essere inibito, altrimenti si avrebbe l’arresto del ciclo cellulare e non apoptosi.
E’ importante che p21 e la proteina 14-3-3σ siano inattive. A questo proposito, e’
stato visto che c-Myc porta all’apoptosi inibendo l’arresto del ciclo cellulare. Questa
proteina è , infatti, reclutata da Miz-1 per sopprimere in modo specifico il promotore
di p21. Anche hDaxx è in grado di reprimere p21. Il bilancio tra p21 e PUMA, che interagisce con
Bcl-2 e promuove la traslocazione al mitocondrio e la multimerizzazione di Bax, sembra essere
decisivo nel determinare la risposta dell’attivazione di p53. Inoltre, l’arresto del ciclo cellulare
potrebbe, in qualche modo, inibire la risposta apoptotica. La prevalenza di arresto del ciclo
Fig.4
cellulare o di apoptosi dipende, inoltre, dall’interazione del pathway di p53 con altri pathway
cellulari: infatti, l’attivazione dei target apoptotici da sola, non è sufficiente a indurre apoptosi in
alcune cellule, e a prevenire l’induzione di p21: altri regolatori del ciclo cellulare sono necessari
per guidare le cellule verso l’apoptosi. (Fig. 4) (J.Yu e L.Zhang, Molecular Cell Biochemical and
Biophysical Research Communications, 2001). L’induzione dell’apoptosi da parte di p53 può
avvenire in modo trascrizione dipendente e trascrizione indipendente, tramite un diretto
reclutamento di p53 sulla membrana mitocondriale (Fig.5) (U.M.Moll, Letters 2001).
Fig.5
Per quanto riguarda l’induzione dell’apoptosi in modo trascrizione dipendente, p53 attiva la
trascrizione dei recettori di morte (DR5, CD95,Fas); di proteine proapoptotiche della famiglia BH3
–only (Bax, PUMA, NOXA, Bid); di adattatori apoptotici come Apaf-1; della proteina PIG-3 che
induce stress ossidativo e causa un aumento dei radicali reattivi dell’ossigeno; di IGF-BD (insulin
growth factor-binding protein), che compete per il legame con l’IGF1R, bloccando la via di
trasduzione dei segnali di sopravvivenza trasmessi da questo recettore in presenza dell’IGF1.
Inoltre, p53 reprime la trascrizione di proteine anti-apoptotiche come Bcl-2 e Bcl-XL (Fig.6) (R.V.
Sionov, Oncogene 1999).
Fig.6
Quindi, p53 promuovendo la trascrizione dei propri target, attiva indirettamente sia la via estrinseca
che la via intrinseca dell’apoptosi. Nell’articolo “PUMA, a novel proapoptotic gene, is induced by
p53”, pubblicato sulla rivista Molecular Cell da Nakano e Vousden nel 2001, viene descritto il
modo in cui è stato identificato PUMA (p53 upregulated modulator of apoptosis). Sono stati
effettuati dei microarray per comparare i pattern di espressione in cellule con o senza p53. Con
questo approccio, sono stati identificati diversi geni la cui espressione è attivata in risposta
all’induzione di p53. Tra questi, era stato trovato un gene, la cui sequenza era omologa ad un
parziale cDNA riportato nel database GenBank, che potenzialmente codificava una proteina
contente il dominio BH3. Con maggior frequenza, sono state poi identificate due sequenza
correlate che sembrano due varianti di splicing alternativo dello stesso trascritto (PUMA α e
PUMAβ). Con diversi Northen Blot, usando una sonda specifica dell’esone 4 di questi trascritti, è
stato confermato che PUMA è indotto da p53. Tramite esperimenti di immunofluorescenza, è stato
visto che PUMA localizza al mitocondrio. Questa proteina inibisce la formazione di colonie in
vitro e induce apoptosi, legando Bcl-2 e inducendo così il rilascio del citocromo c e l’attività delle
procaspasi 3 e 9. L’inibizione dell’espressione di PUMA, con specifici oligonucleotidi anti-senso,
riduce l’apoptosi mediata da p53 . Per quanto riguarda l’induzione dell’apoptosi trascrizioneindipendente, ci sono diversi modelli riguardo l’azione di p53 a livello mitocondriale. In base al
primo modello, p53 si lega alle proteine anti-apoptotiche Bcl-2 e Bcl-XL ed è successivamente
spiazzata dalla proteina PUMA, la cui trascrizione è attivata da p53 stessa. In questo modo p53
sarebbe attivata, e quindi, potrebbe andare a legare le proteine Bax e Bak, inducendo la loro
oligomerizzazione, con il conseguente aumento della permeabilità mitocondriale e il rilascio del
citocromo c (Fig.9) (D.R.Green, Trends in genetics 2003).
Fig.9
In base al secondo modello, p53 è in grado di legare solo Bcl-2 e Bcl-XL; questo legame
permetterebbe a Bak e Bax di sganciarsi da Bcl-2 e Bcl-XL, dalle quali sono tenute inattive in
cellule in condizioni normali. L’azione di p53 è proprio quella di inattivare Bcl-2 e Bcl-XL, in
modo tale che Bak e Bax possano a questo punto essere attivate dalla forma tronca di Bid (tBid),
uno degli attivatori dell’apoptosi che induce la loro oligomerizzazione, con conseguente
Fig.10
formazione dei pori sulla membrana mitocondriale esterna (Fig.10) (B.M Polster, Journal of
neurochemistry 2003). Concludendo, quindi, secondo il primo modello p53 si comporterebbe come
un attivatore dell’apoptosi, al pari di Bid e Bim, in quanto in grado di legare direttamente Bax e
Bak; in base al secondo modello, invece, p53 si comporterebbe come un sensibilizzatore
dell’apoptosi, al pari di Bad e Bik, in quanto in grado di legare direttamente solo Bcl-2 e Bcl-XL e,
quindi, in grado di attivare solo indirettamente Bak e Bax (Fig.11).
Fig.11
In uno degli articoli pubblicati da Mihara et al. sulla rivista Molecular Cell nel 2003, “P53 has an
apoptogenic Role at the Mithocondria”, viene dimostrato il ruolo di p53 come sensibilizzatore
dell’apoptosi e, quindi la validità del secondo modello. In timociti in coltura irradiati con radiazioni
γ, p53 localizza, in tempi molto brevi, a livello mitocondriale (Fig. 12B), dove interagisce con Bcl2 e Bcl-XL (Fig.12C) ma non con Bax e Bak (Fig.12D). Le interazioni di p53 sono state studiate
effettuando nel primo caso un’immunoprecipitazione con Bcl-XL e un blot anti-p53; nel secondo
un’immunoprecipitazione con p53 e un blot anti-Bax.