Molecole di Adesione
Dott.ssa
Erminia La Camera
L’adesione e la migrazione dei leucociti sono regolate dal legame di
molecole di adesione complementari presenti sui leucociti e sulle superfici
endoteliali.
I mediatori chimici chemioattrattori ed alcune citochine, influiscono su
questi processi modulando l’espressione di superficie e l’avidità di queste
molecole di adesione.
I recettori di adesione coinvolti appartengono a quattro famiglie molecolari:
Le Selectine
La Super Famiglia delle Immunoglobuline
Le Integrine
Le Glicoproteine mucina-simile
SELECTINE
Le selectine sono una famiglia composta da tre proteine correlate tra loro,
che differiscono nella distribuzione cellulare, ma agiscono tutte
nell’adesione dei leucociti alle cellule endoteliali.
Tutte sono glicoproteine transmembranarie a catena singola con la porzione
N-terminale legata a proteine che legano i carboidrati, note come lectine di
tipo C.
Il legame delle selectine ai loro ligandi è calcio-dipendente, ha un’alta
velocità di attacco, ma anche una elevata velocità di scissione ed è a bassa
affinità; questa proprietà consente alle selectine di mediare l’iniziale
adesione e il conseguente rotolamento dei leucociti sull’endotelio in
presenza del flusso ematico.
La L-selectina o CD62L è localizzata all’apice dei microvilli dei leucociti, il
che facilita la sua interazione con il ligando sull’endotelio.
Può servire come un recettore che indirizza i linfociti ad entrare nei linfonodi
legandosi alle venule endoteliale alte.
Essa agisce anche legando i neutrofili alle cellule endoteliali attivate dalle
citochine nelle sedi di infiammazione.
Tre ligandi delle cellule endoteliali possono legare la L-selectina:
Molecola di adesione cellulare contenente glicano 1
Molecola di adesione cellulare mucosa-addressina 1
CD34, un proteoglicano espresso sulle cellule endoteliali
Tutti questi ligandi sono modificati per mezzo dell’aggiunta di specifici
carboidrati che sono in realtà i residui riconosciuti dalla selectina.
La E-selectina o CD62E, definita in precedenza come molecola di adesione
leucocitaria-endoteliale 1, è espressa soltanto nelle cellule endoteliali
attivate dalle citochine.
Riconosce gruppi carboidrati sialilati complessi, correlati alla famiglia delle
Lewis X o Lewis A, un gruppo di molecole presente su varie proteine che si
trovano sulla superficie di granulociti, monociti, cellule T effettrici e cellule
della memoria attivate.
L’espressione delle E-selectine nelle cellule endoteliali è una caratteristica
dell’infiammazione acuta mediata da citochine e gli anticorpi anti E-selectina
possono bloccare l’accumulo dei leucociti in vivo.
La P-selectina è stata identificata per la prima
volta nei granuli secreti dalle piastrine, ma poi
anche nei granuli di secrezione delle cellule
endoteliali chiamati corpi di Weibel-Palade.
Quando le cellule endoteliali o le piastrine sono
stimolate, la P-selectina trasloca entro pochi
minuti sulla superfici cellulare. Una volta
raggiunta tale sede, la P-selectina media il
legame con i neutrofili, i linfociti T ed i
monociti.
I ligandi formati da carboidrati complessi
riconosciuti dalla P-selectina sono simili a quelli
per la E-selectina.
INTEGRINE
La superfamiglia delle integrine è costituita da 30 proteine strutturalmente omologhe,
che promuovono le interazione tra cellule o tra cellula e matrice.
Il nome deriva dall’ipotesi che esse coordinano (cioè “integrino”) i segnali trasmessi dai
ligandi extracellulari con mobilità citoscheletro-dipendente, cambiamenti di forma e
risposte fagocitarie.
Tutte sono proteine di superficie cellulare eterodimeriche composte da due catene
polipeptidiche associate in modo non covalente α e β. I domini extracellulari delle due
catene legano vari ligandi, che comprendono glicoproteine della matrice extracellulare,
componenti attivate del complemento e proteine della superficie di altre cellule. I
domini citoplasmatici interagiscono con componenti citoscheletriche (comprese la
vinculina, la talina, l’actina, l’α-actina e la tropomiosina).
Le integrine contenenti la catena β1
sono dette anche molecole VLA (Very
Late Activation) cioè molecole di
attivazione tardiva, sono ampiamente
espresse sui leucociti e su altre cellule
e mediano l’attacco delle cellule alle
matrici extracellulari.
La VLA-4 (α4β1) è espressa soltanto
sui leucociti e può mediare l’attacco di
queste
cellule
all’endotelio
interagendo con la molecola di
adesione cellulare vascolare 1 (VCAM1). La VLA-4 è una delle principali
proteine di superficie che mediano
l’indirizzamento dei linfociti verso
l’endotelio nelle sedi periferiche
dell’infiammazione.
Le integrine che contengono la catena β2
sono anche dette CD11a-cCD18, o famiglia
degli antigeni associati alla funzione
leucocitaria 1; CD11a-c si riferisce alle
diverse catene α e CD18 si riferisce alla
subunità comune β2. La CD11a-cCD18
svolge un ruolo importante nell’adesione dei
linfociti e degli altri leucociti alle altre cellule,
come le cellule presentanti l’antigene e le
cellule dell’endotelio vascolare. Altri membri
della famiglia comprendono CD11bCD18 e
CD11cCD18, che mediano l’attacco dei
leucociti alle cellule endoteliali e la
successiva fuoriuscita dai vasi.
Le altre integrine sono espresse su piastrine
e altri tipi cellulari e si legano alle proteine
della matrice extracellulare e alle proteine
coinvolte nella coagulazione.
IMMUNOGLOBULINE
La famiglia delle immunoglobuline comprende due molecole
di adesione endoteliale:
ICAM-1
VCAM-1
Entrambe fungono da ligandi per le integrine che si trovano
sui leucociti.
GLICOPROTEINE MUCINA-SIMILI
Le
glicoproteine
mucina-simili,
come
l’eparansolfato, fungono da ligandi per la molecola
di adesione leucocitaria chiamata CD44 e sono
presenti nella matrice extracellulare e sulle
superfici cellulari.
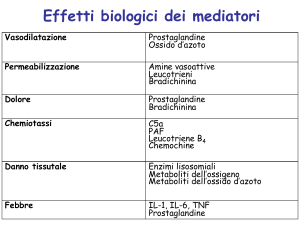
MEDIATORI CHIMICI DELL’INFIAMMAZIONE
I mediatori di origine cellulare sono sequestrati nei granuli intracellulari che devono
essere secreti (istamina) o sintetizzati ex novo in risposta ad uno stimolo
(prostaglandine e citochine)
MEDIATORI CHIMICI DELL’INFIAMMAZIONE
I mediatori di origine plasmatica (es. proteine del complemento, chinine) sono
presenti nel plasma in forma di precursori che devono essere attivati, da tagli
proteolitici, per acquisire le loro proprietà biologiche.
La produzione dei mediatori attivi è scatenata da prodotti microbici o da proteine
dell’ospite (es. C3a, C5c, fattori della coagulazione e chinine) che sono attivati a loro
volta da microbi o tessuti danneggiati.
I Mediatori esercitano il loro effetto biologico inizialmente legandosi a particolari
recettori posti sulla cellula bersaglio.
Un mediatore può stimolare il rilascio di altri mediatori da parte delle stesse
cellule bersaglio.
I mediatori possono agire su uno o pochi tipi di cellule bersaglio o possono avere
effetti diversi su differenti tipi cellulari.
Sono altamente specifici, hanno vita breve e sono potenzialmente dannosi.
AMINE VASOATTIVE
ISTAMINA
• Rilasciata dai granuli dei mastociti in risposta a lesioni fisiche (freddo o
traumi), a reazioni immunitarie che interessano i mastociti, alle
anafilotossine (frammenti del complemento), a proteine di derivazione
leucocitaria, a neuropeptidi e a citochine
• Causa la dilatazione delle arteriole, la contrazione della muscolatura
liscia e l’aumento della permeabilità vascolare
• Ha un effetto diretto sulle cellule endoteliali dei capillari venosi che si
contraggono liberando degli spazi tra le cellule adiacenti
• È un leggero chemioattrattore per gli eosinofili
SEROTONINA
• Rilasciata dalle piastrine in seguito all’ attivazione dell’aggregazione
piastrinica e dal fattore attivante delle piastrine (PAF) che deriva dai
mastociti durante le reazioni mediate dalle IgE.
PROTEINE PLASMATICHE
Proteine del sistema del complemento
Le proteine del complemento sono 9, sono presenti nel plasma in forma inattiva e sono
numerate da C1 a C9. L’attivazione della cascata del complemento può essere divisa in
fasi precoci e fasi tardive.
Nelle fasi precoci, tre vie diverse portano alla scissione del C3.
Via classica: C1-Ag-Ac
Via alternativa: stimolata da molecole di
superficie dei microbi ma non è presente Ac
Via della lectina: C1 viene attivato
dal legame tra lectina e i
carboidrati presenti sui microbi
C3
C3 convertasi
C3a
C3b
C5 convertasi
C5a
Si lega alla molecola che
ha scatenato l’attivazione
C5b
Nelle fasi tardive, le tre vie convergono ed il prodotto di degradazione del C3, C3b,
attiva una serie di altre componenti del complemento.
Fenomeni vascolari: C3a e C5a stimolano i mastociti a rilasciare istamina
facendo aumentare la permeabilità vasale e inducendo vasodilatazione.
C5a nei neutrofili e nei monociti attiva la via lipossigenasica del
metabolismo dell’acido arachidonico provocando un ulteriore rilascio di
mediatori dell’infiammazione
Fenomeni di adesione, chemiotassi e attivazione: C5a è un potente agente
chemiotattico per neutrofili, monociti, eosinofili e basofili
Fagocitosi: C3b si comporta da opsonina e favorisce la fagocitosi da parte
dei neutrofili e macrofagi
PROTEINE PLASMATICHE
Sistema delle chinine
L’attivazione del sistema delle chinine porta al rilascio del peptide vasoattivo
bradichinina che aumenta la permeabilità vasale, causa vasodilatazione, contrazione
del muscolo liscio e dolore nella cute quando iniettata. L’azione è di breve durata
perchè viene inattivata dall’enzima chininasi e quella residua viene inattivata
dall’enzima angiotensina-convertasi.
PROTEINE PLASMATICHE
Sistema della coagulazione
Il sistema della coagulazione è diviso in due vie convergenti che culminano
nell’attivazione della trombina e nella formazione di fibrina.
La proteasi trombina fa da collegamento tra il sistema della coagulazione e
l’infiammazione, sia perchè è la principale proteasi della coagulazione sia perché si lega
ai recettori attivati dalla proteasi. Questo determina il reclutamento dei leucociti
mobilizzando P-selectina, producendo chemochine ed esprimendo molecole di adesione.
Il fattore XII oltre ad indurre la coagulazione, può attivare il sistema fibrinolitico che
controbilancia la coagulazione degradando la fibrina e solubilizzando il coagulo fibrinoso.
L’attivatore del plasminogeno scinde il plasminogeno in plasmina, una proteina
multifunzionale, che lisa i coaguli di fibrina, cliva il C3 producendo i frammenti di C3 e
scinde la fibrina per formare i prodotti della scissione della fibrina che inducono
permeabilità vasale.
CONCLUSIONI
I mediatori più importanti in vivo sono: bradichinina, C3a e C5a (per l’aumento della
permeabilità vasale), C5a (per la chemiotassi) e la trombina (che ha effetti sulle
cellule endoteliali)
Il C3a e il C5a possono essere prodotti da:
reazioni immunitarie che coinvolgono anticorpi e complemento (via classica);
attivazione della via alternativa o della via della lectina in assenza di anticorpi;
agenti esterni come la callicreina, plasmina o serino-proteasi
Il fattore di Hageman attivato (XIIa) dà inizio a quattro sistemi coinvolti nella
risposta infiammatoria:
il sistema delle chinine, che produce chinine vasoattive;
il sistema della coagulazione, che produce trombina, fibrinopeptidi e fattore X
il sistema fibrinolitico, che produce plasmina e degrada fibrina;
il sistema del complemento, che produca anafilotossine.
La callicreina può, tramite un meccanismo a feedback, attivare il fattore di Hageman
e amplificare massivamente gli effetti innescati dallo stimolo iniziale.
METABOLITI DELL’ACIDO ARACHIDONICO
L’ acido arachidonico è un acido grasso polinsaturo a 20 atomi di carbonio
che deriva da fonti alimentari o dalla conversione di un acido grasso
essenziale, l’acido linoleico.
L’acido arachidonico non si trova libero nelle cellule, ma è esterificato nei
fosfolipidi di membrana, dai quali viene rilasciato sotto l’azione di stimoli
meccanici, fisici, chimici o da altri mediatori.
I metaboliti dell’acido arachidonico, detti anche eicosanoidi, sono sintetizzati
da due classi di enzimi: ciclossigenasi e lipossigenasi; una volta formati si
legano ai recettori accoppiati alle proteine G presenti su molti tipi cellulari e
possono mediare ogni fase dell’infiammazione, si possono trovare negli
essudati infiammatori e la loro sintesi è aumentata in sede di infiammazione.
Via della ciclossigenasi: mediata da due enzimi (COX-1 e COX-2) porta
alla formazione di prostaglandine
Via della lipossigenasi: mediata da tre enzimi (il predominante è 5-LO)
porta alla formazione di leucotrieni
FATTORE ATTIVANTE LE PIASTINE (PAF)
Il PAF è un acetil-gliceril-etere-fosforilcolina, un fosfolipide con una tipica impalcatura di
glicerolo, un acido grasso a catena lunga in posizione A, una insolita catena corta in
posizione B e un radicale di fosfatidilcolina. E’ prodotto da basofili, piastrine, macrofagi e
cellule endoteliali e media il suo effetto tramite un recettore associato alla proteina G.
PAF
Vasodilatazione
Vasocostrizione
Aumenta la permeabilità
vasale
Broncocostrizione
CITOCHINE
TNF e IL-1
TNF-α
Prodotto da numerose cellule (monociti e linfociti T) come precursore
Attiva i macrofagi: induce fagocitosi, produzione di NO
Attiva le cellule endoteliali: aumenta la permeabilità ed induce
espressione di molecole di adesione
Induce espressione ed attivazione di cicloossigenasi e lipossigenasi
IL-1
Prodotta da monociti/macrofagi, cellule epiteliali, cellule B, fibroblasti
Viene espressa come precursore, agisce insieme al TNF
Induce espressione di molecole di adesione (E-selectina, ICAM-1)
Attiva i leucociti polimorfonucleati
Stimola la produzione di altre citochine (IL-2, IL-3, IL-4, IFNγ, TGFβ)
Aumenta la temperatura corporea
Stimola la proliferazione dei fibroblasti e l’angiogenesi
Effetti sistemici delle citochine
Le citochine agiscono su tessuti distanti per migliorare la risposta immunitaria
Fegato
Proteine della
fase acuta
(Proteine Creattive)
Attivazione del
Complemento,
Opsonizzazione
Midollo Osseo
Epitelio
Mobilizzazione
dei Neutrofili
Fagocitosi
Ipotalamo
Grasso, muscoli
Cellule
Dendritiche
Aumento della
Temperatura
corporea
Mobilizzazione di
proteine ed
energia per
favorire
l’incremeto della
temperatura
TNF- stimola
la migrazione ai
linfonodi e la
maturazione
Riduzione replicazione batterica e virale.
Aumentata elaborazione dell’antigene.
Incremento risposta immunitaria specifica
Inizio risposta
immunitaria
specifica
CHEMOCHINE
Sono una famiglia di circa 40 proteine con funzione chemiotattica per
monociti, linfociti, granulociti neutrofili ed eosinofili.
Vengono classificate in quattro gruppi in base alla disposizione dei residui di
Cys nelle proteine mature:
Chemochine C-X-C (α chemochine): IL-8 secreta dai macrofagi attivati e
dalle cellule endoteliali attiva i neutrofili e induce la loro chemiotassi
Chemochine C-C (β chemochine): proteina chemiotattica per i monociti
(MCP-1) e proteina infiammatoria dei macrofagi (MIP-1α) che attraggono
monociti, basofili, linfociti ma non neutrofili
Chemochine C (γ chemochine): mancano di due residui di Cys e sono
specifiche per i linfociti
Chemochine CX3C: unico elemento del gruppo è la fractalchina che esiste
in due forme, una legata alla superficie cellulare che favorisce l’adesione di
monociti e linfociti T; e una solubile con effetto chemiotattico di attrazione
per le stesse cellule.
OSSIDO NITRICO
L’Ossido Nitrico (NO) è un gas solubile prodotto dalle cellule endoteliali e
dai macrofagi. Viene sintetizzato dall’enzima ossido nitrico sintetasi (NOS) a
partire dalla L-arginina. Esistono tre tipi di NOS:
eNOS e nNOS espressi costitutivamente a bassi livelli e attivati da un
aumento delle [Ca2+]; iNOS indotto quando i macrofagi sono attivati da
citochine (TNF o INF-γ) o da altri agenti.
NO è un potente vasodilatatore, riduce l’aggregazione e l’adesione, funge da
regolatore endogeno del reclutamento dei leucociti.
RADICALI LIBERI DELL’OSSIGENO
macrofago
LPS
IFN
iNOS
L-a
rgi
nin
a
NO
NO + O2●
ONOO- + H+
Anione
perossinitrito
ONOOH
●OH
+ NO2 ●
Stimolano la produzione di citochine,
chemochine e molecole di adesione
endoteliale dei leucociti, amplificando
la cascata che evoca la risposta
infiammatoria
Radicali
liberi
provocano danno alle
cellule endoteliali con
conseguente aumento
della permeabilità
vascolare
inattivano gli inibitori
della proteasi
possono essere
dannosi per altre cellule
COSTITUENTI LISOSOMIALI DEI LEUCOCITI
Sono presenti nei neutrofili e nei monociti.
Nei neutrofili ne esistono due tipi:
• Granuli azzurrofili che contengono fattori battericidi (lisozima), idrolasi,
proteasi e mieolperossidasi
• Granuli specifici che contegono lisozima, collagenasi, gelatinasi, istaminasi
e fosfatasi alcalina.
Entrambi possono svuotarsi nei vacuoli di fagocitosi che avvolgono il
materiale inglobato, oppure il contenuto può essere riversato nello spazio
extracellulare.
Gli enzimi presenti nei granuli svolgono varie funzioni:
• le proteasi acide degradano i batteri e i detriti all’interno dei fagolisosomi
• le proteasi neutre degradano varie componenti extracellulari (collagene,
fibrina, elastina) e scindono anche il C3 e C5 liberando le anafilotossine.
Poiché queste proteasi sono molto nocive, nel siero e nei liquidi dei tessuti
sono presenti delle proteine che ne inibiscono l’attività, la più importante è
l’ α- antitripsina.
NEUROPEPTIDI
Sostanza P e Neurochinina A sono prodotte nel sistema nervoso centrale e
periferico, in particolare la sostanza P è presente nelle fibre nervose del
polmone e del tratto gastrointestinale.
Hanno molte funzioni biologiche:
Aumentano la permeabilità vascolare
Regolano la pressione sanguigna
Trasmettono segnali dolorosi
Stimolano la secrezione delle cellule endocrine
ESITI DELL’INFIAMMAZIONE ACUTA
L’infiammazione acuta può presentare tre esiti:
Risoluzione completa:
Avviene quando il danno è lieve e di breve durata, le cellule possono
rigenerarsi, i mediatori chimici vengono inattivati, la permeabilità vascolare
rientra nella norma, i neutrofili muoiono, i leucociti, le proteine e gli agenti
estranei vengono rimossi.
Guarigione tramite fibrosi:
Avviene quando il danno è lieve e di lunga durata, le cellule non sono in
grado di rigenerarsi o vi sia un abbondante essudato ricco di fibrina.
Infiammazione Cronica:
L’infiammazione non si risolve a causa della persistenza dell’agente lesivo o a
causa di qualche interferenza con il normale processo di guarigione.