Macromolecole Biologiche
Chimica Biologica
A.A. 2010-2011
Struttura e meccanismo
d’azione degli enzimi
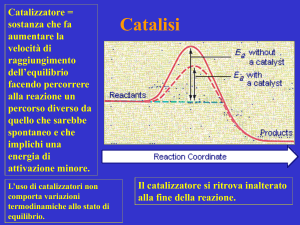
Catalisi chimica
Catalisi chimica: teoria dello stato di transizione
- descrizione microscopica del sistema:
le molecole di reagenti per trasformarsi prodotti procedono lungo una
traiettoria di reazione
⇒ questo progresso viene descritto da una coordinata di reazione
“diagramma dello stato di transizione” o
“diagramma della coordinata di reazione”
(correla l’energia libera alla coordinata di reazione)
il punto di massima G corrisponde allo stato di transizione (o complesso attivato)
Catalisi chimica
Catalisi chimica: teoria dello stato di transizione
A + B → X‡ → P
A, B = reagenti
P = prodotto
X‡ = stato di transizione
punto della reazione corrispondente
al max di energia libera
Coordinata di reazione: percorso di
minima energia libera lungo il quale
i reagenti si avvicinano l’un l’altro
ΔGreazione< 0 ⇒ reazione spontanea
ΔG‡ = energia libera di attivazione
Catalisi chimica
Correlazione tra velocità di una reazione ed energia di
attivazione
K‡
A, B = reagenti
P = prodotto
k’
⇀ X‡ → P
A+B↽
k
A+B→P
velocità della reazione: d[P] = k [A][B] = k’ [X‡]
dt
k = costante di velocità ordinaria della reazione elementare
k’ = costante di velocità di decomposizione di X‡ nei prodotti
assunzione: anche se instabile, X‡ è in rapido equilibrio con i reagenti
‡]
[X
⇒ K‡ =
[A][B]
costante di equilibrio ⇒
⇒ K‡ può essere espressa come: -RT ln K‡ = ΔG‡
Catalisi chimica
Correlazione tra velocità di una reazione ed energia di
attivazione
d[P] = k [A][B] = k’ [X‡] = k’ K‡ [A][B] = k’ exp (-ΔG‡/RT) [A][B]
dt
⇒ la velocità di reazione non dipende solo dalla concentrazione dei
reagenti ma diminuisce in modo esponenziale con ΔG‡ (differenza tra
l’energia libera dello stato di transizione e quello dei reagenti)
maggiore è ΔG‡ ⇒ meno stabile è lo stato di transizione ⇒ più lenta è la
reazione
k = k’ exp (-ΔG‡/RT) = (KBT/h) exp (-ΔG‡/RT)
KB = costante di Boltzmann = 1.3807 1023 J K-1
h = costante di Plank = 6.6261 10-34 J s
R = costante dei gas = 8.31 J mol-1 K-1
Catalisi chimica
Correlazione tra velocità di una reazione ed energia di
attivazione
K‡
k’
A+B ↽
⇀ X‡ → P
k’ = costante di velocità di decomposizione di X‡ nei prodotti
lo stato di transizione è stabilizzato da un legame debole che può rompersi
alla prima escursione vibrazionale
k’ = κν
ν = frequenza vibrazionale del legame che si spezza quando X‡ si decompone
κ = coeff. di trasmissione (0-1)
(probabilità che la rottura di sia nella direzione della formazione del prodotto)
Catalisi chimica
Correlazione tra velocità di una reazione ed energia di
attivazione
K‡
k’
A+B ↽
⇀ X‡ → P
k’ = κν
ν = ε/h
ε = energia media della vibrazione che porta alla decomposizione di X‡
h = costante di Plank
ad una temperatura T l’energia classica di un oscillatore è
ε = KBT (energia termica disponibile)
KB = costante di Boltzmann
⇒ k’ = κν = KBT/h ⇒ k = k’ exp (-ΔG‡/RT) = (KBT/h) exp (-ΔG‡/RT)
si assume κ = 1
all’aumentare di T e quindi dell’energia termica
disponibile, aumenta la velocità della reazione
Catalisi chimica
Correlazione tra velocità di una reazione ed energia di
attivazione
- passaggio attraverso lo stato di transizione: 10-13 -10-14 s
- la decomposizione dello stato di transizione nei prodotti (o viceversa, nei
reagenti) è il processo che determina la velocità di reazione totale
A→I→P
I = intermedio di reazione
2 stati di transizione
2 energie di attivazione
- la tappa con lo stato di transizione
a maggior energia libera determina
la velocità di reazione
Catalisi chimica
I catalizzatori riducono l’energia libera di attivazione
I catalizzatori si combinano temporaneamente con i reagenti promuovendo così
il loro ingresso nella condizione reattiva dello stato di transizione
• abbassano il ΔG‡
• non alterano ΔGreazione
(reazione spontanea diretta se
ΔGreazione < 0, altrimenti inversa)
• accelerano allo stesso modo la
reazione diretta e quella inversa
(la reazione raggiunge l’equilibrio
più velocemente)
ΔΔG‡cat = efficienza del catalizzatore = ΔG‡noncat -ΔG‡cat
Catalisi chimica
I catalizzatori riducono l’energia libera di attivazione
d[P] = k’ exp (-ΔG‡/RT) [A][B]
dt
aumento di velocità: rapporto fra velocità di reazione catalizzata e non catalizzata
exp(-ΔG‡cat /RT) / exp(-ΔG‡noncat /RT) = exp(ΔG‡noncat -ΔG‡cat)/RT = exp(ΔΔG‡cat)
dove ΔΔG‡cat = efficienza del catalizzatore
Esempio: a T = 25 °C (298 K)
aumento di velocità di 10 volte ⇒ ΔΔG‡ ~ 5.71 kJ/mol
(meno della metà dell’energia libera di un tipico legame a idrogeno)
aumento di velocità di 106 volte ⇒ ΔΔG‡ ~ 34 kJ/mol
(piccola frazione dell’energia libera della maggior parte dei legami covalenti)
Catalisi chimica
I catalizzatori riducono l’energia libera di attivazione
si consideri la conversione di A in P:
kF
⇀
A↽ P
kR
se in assenza di un catalizzatore la costante di velocità da A a P è, per
esempio, kF 10-4 s-1 mentre la costante di velocità della reazione inversa è
kR 10-6 s-1
⇒ la costante di equilibrio è K = kF / kR = [P]/[A] = 100
⇒ all’equilibrio la concentrazione di P è 100 volte quella di A
tale valore non cambia se la reazione è catalizzata, ma l’equilibrio si
raggiunge più in fretta
Proprietà generali degli enzimi
Catalisi enzimatica
- le diverse proteine svolgono funzioni biologiche cui si fa riferimento col
termine di attività biologica: ogni specie proteica ha una propria attività
biologica
- gli enzimi sono i catalizzatori biologici
- l’attività biologica di un enzima è una specifica attività catalitica:
capacità di accelerare una determinata reazione chimica
- gli enzimi aumentano la velocità delle reazioni chimiche abbassando la
barriera di energia libera che separa i reagenti dai prodotti
- i reagenti di una reazione catalizzata da un enzima sono detti substrati
di quell’enzima
Proprietà generali degli enzimi
Interazioni enzima-ligandi nella trasformazione del
substrato in prodotto
- gli enzimi agiscono legando transientemente substrati, intermedi, stati di
transizione, e prodotti della reazione catalizzata
- la reazione avviene con queste varie specie che si trasformano da
reagenti in prodotti mantenendosi in complesso con l’enzima
- l’enzima crea un ambiente in cui lo stato di transizione è stabilizzato
e in cui la reazione procede quindi più velocemente di quando avviene
quando le specie coinvolte sono in forma libera
Proprietà generali degli enzimi
Interazioni enzima-ligandi nella trasformazione del
substrato in prodotto
una reazione catalizzata enzimaticamente
procede con un meccanismo diverso da
quella non catalizzata, con stati di transizione
aventi una ΔG‡ inferiore
Proprietà generali degli enzimi
Catalisi chimica e catalisi enzimatica
- gli enzimi catalizzano specifiche reazioni chimiche attraverso
meccanismi di per sé molto simili a quelli operanti nella normale
catalisi chimica
- il meccanismo catalitico di un enzima è sempre una combinazione di
più meccanismi elementari di catalisi chimica
- il processo catalitico di un enzima interessa principalmente una porzione
della molecola della proteina enzimatica, detta sito attivo
- nel sito attivo sono presenti vari gruppi funzionali, detti gruppi
catalitici, disposti e orientati in modo opportuno per legare i substrati e
promuovere la loro trasformazione nei prodotti
Proprietà generali degli enzimi
queste reazioni di fatto non
- le reazioni catalizzate dagli enzimi sono: avvengono senza catalizzatore
• 106-1014 volte più veloci delle reazioni non catalizzate
• alcuni ordini di grandezza più veloci delle reazioni catalizzate chimicamente
Proprietà generali degli enzimi
Differenze rispetto ai normali catalizzatori chimici
(1) velocità di reazione più elevate
(2) condizioni di reazione più blande: T < 100 °C, P = Patm, pH ~ neutro
(una catalisi chimica efficiente invece può richiedere T e P elevate e pH estremi)
(3) capacità di regolazione:
- controllo allosterico
- modificazione covalente degli enzimi
- variazione della quantità di enzima sintetizzato
(4) elevata specificità di reazione: nel riconoscimento dei substrati e prodotti
(raramente si hanno prodotti collaterali)
Proprietà generali degli enzimi
(4) elevata specificità di reazione
i substrati (ed altre molecole) si legano ai siti attivi
(cavità interne o fessure superficiali) degli enzimi
mediante interazioni non-covalenti
- complementarità geometrica
- complementarità elettronica
- modelli per l’interazione enzima-substrato:
• modello chiave-serratura
• modello dell’adattamento indotto
- in realtà il sito attivo più che essere complementare al substrato, è
complementare allo stato di transizione
Proprietà generali degli enzimi
• Modello chiave-serratura
siti di legame pre-strutturati, complementarità fra sito attivo enzima e
substrato e nessun cambio conformazionale dovuto al legame del substrato
• Modello dell’adattamento indotto
siti di legame pre-strutturati, ma cambi conformazionali dovuti al legame
del substrato
Proprietà generali degli enzimi
(4) elevata specificità di reazione
(4a) stereospecificità
- specificità nel legare substrati chirali e nel catalizzare le loro reazioni
- intrinseca asimmetria dei siti attivi (formati solo da L-AA)
- quasi tutti gli enzimi che partecipano a reazioni chirali sono assolutamente
stereospecifici
Esempio: Aconitasi (ciclo dell’acido citrico)
Citrato = molecola prochirale
⇒ diventa chirale per sostituzione di uno dei 2
gruppi carbossimetilici –CH2COO⇒ interazione asimmetrica (su 3 punti) fra citrato ed enzima
Proprietà generali degli enzimi
(4) elevata specificità di reazione
(4b) specificità geometrica
- selettività nel riconoscimento dell’identità dei gruppi chimici dei substrati
- gli enzimi possono avere una specificità geometrica molto variabile
- “preferenza” (più che specificità geometrica) quando i substrati accettati
sono svariati (esempio: gli enzimi digestivi)
Esempio: Chimotripsina
mancanza di specificità anche per il tipo di reazione catalizzata (eccezione alla
regola) ⇒ idrolisi di legame peptidico ed estere
Proprietà generali degli enzimi
Nomenclatura degli Enzimi
IUBMB (International Union of Biochemistry and Molecular Biology):
organismo internazionale che si occupa della classificazione funzionale e
sistematica degli enzimi e della loro denominazione
http://www.iubmb.unibe.ch
- classificazione e denominazione in base alla reazione catalizzata
- ad ogni enzima sono assegnati 2 nomi e 1 numero:
Nome d’uso raccomandato
(nome comunemente usato, spesso di origine “storica”)
Nome sistematico
(nome dei substrati + termine con suffisso –asi, che definisce il tipo di reazione)
Numero di classificazione
Proprietà generali degli enzimi
Nomenclatura degli Enzimi
Numero di classificazione
formato: (EC X.Y.Z.W)
esempio:
EC 1.1.1.1
“EC”: Enzyme Commission
X: Classe
Y: Sottoclasse
Z: Sotto-sottoclasse
W: numero individuale
- esistono 6 classi principali di reazioni enzimatiche
Proprietà generali degli enzimi
Nomenclatura degli Enzimi
Classe di enzimi
Reazioni catalizzate
1. Ossidoriduttasi
Reazioni di ossidoriduzione
2. Transferasi
Trasferimento di gruppi funzionali
3. Idrolasi
Reazioni di idrolisi
Reazioni di eliminazione con formazione di
doppi legami
Isomerizzazioni
Formazione di legami accoppiata all’idrolisi
di ATP
4. Liasi
5. Isomerasi
6. Ligasi
Proprietà generali degli enzimi
Nomenclatura degli Enzimi
Proprietà generali degli enzimi
Nomenclatura degli Enzimi
Esempi:
alcool deidrogenasi
alcool:NAD+ ossidoriduttasi
EC 1.1.1.1
(nome raccomandato)
(nome sistematico)
(numero di classificazione)
carbossipeptidasi A
peptidil-L-amminoacido-idrolasi
EC 3.4.17.1
(nome raccomandato)
(nome sistematico)
(numero di classificazione)
Cofattori
Cofattori e Coenzimi
i gruppi funzionali delle proteine sono molto adatti per:
- partecipare a reazioni acido-base
- formare legami covalenti transitori
- partecipare ad interazioni carica-carica
meno adatti per:
- catalizzare reazioni di ossidoriduzione
- catalizzare processi con trasferimenti di gruppi
Cofattori
⇒ molti enzimi agiscono con l’ausilio di specifiche sostanze non peptidiche:
i cofattori
Cofattori
Cofattori e Coenzimi
Ioni metallici (Cu2+, Fe3+, Zn2+)
Cofattori
Cosubstrati
Coenzimi (molecole organiche)
Gruppi prostetici
Cosubstrati: sono associati transientemente all’enzima e quindi funzionano
da cosubstrati
Gruppi prostetici: sono permanentemente associati all’enzima, spesso anche
con legami covalenti
“Oloenzima”= complesso enzima-cofattore cataliticamente attivo
“Apoenzima”= proteina inattiva a seguito della rimozione del cofattore
Cofattori
Ioni metallici
- effetti tossici di alcuni ioni metallici: Cd2+ e Hg2+ possono sostituire lo
Zn2+ (stesso gruppo della tavola periodica ) nel sito attivo di certi enzimi
inattivandoli
Cofattori
Coenzimi
a) I coenzimi devono essere rigenerati
- per completare un ciclo catalitico, i coenzimi che vengono modificati
chimicamente devono tornare al loro stato originale
cosubstrati ⇒ la reazione può essere catalizzata da un enzima diverso
gruppi prostetici ⇒ la reazione avviene come fase (separata) della sequenza
della reazione enzimatica
Cofattori
Coenzimi
Esempio: NAD+ e NADP+
trasportatori intracellulari
di elettroni
Riduzione:
trasferimento di 2 atomi di idrogeno H·
o di uno ione idruro H:- e di un protone H+
ADH = alcool deidrogenasi
Cofattori
Coenzimi
b) Molte vitamine sono precursori di coenzimi
- molti organismi non sono in grado di sintetizzare alcuni coenzimi o
parti di essi
⇒ tali parti sono introdotti con la dieta attraverso le vitamine
- negli organismi superiori i meccanismi di sintesi delle vitamine sono
andati persi evolutivamente
- solo le vitamine idrosolubili sono precursori di cofattori (non quelle
liposolubili come la vitamina A e D)
Cofattori
Coenzimi
b) Molte vitamine sono precursori di coenzimi
Cofattori
Coenzimi
b) Molte vitamine sono precursori di coenzimi
Meccanismi di Catalisi
Meccanismi di Catalisi
•
i catalizzatori riducono l’energia libera dello stato di transizione ΔG‡
stabilizzando lo stato di transizione della reazione catalizzata
•
l’efficienza degli enzimi è dovuta alla loro specificità nel legare i
substrati combinata alla disposizione dei gruppi catalitici
classificazione meccanismi di catalisi
1) Catalisi acido-base
2) Catalisi covalente
3) Catalisi favorita da ioni metallici
4) Effetti di prossimità e orientamento
5) Legame preferenziale dello stato di transizione
Meccanismi di Catalisi
Catalisi acido-base
•
Catalisi acida generale
l’energia libera dello stato di transizione di una reazione viene abbassato
a seguito del trasferimento temporaneo di un protone da un acido
Esempio: tautomerizzazione cheto-enolica
reazione non catalizzata
reazione lenta: elevata energia libera
dello stato di transizione, simile ad un
carbanione
catalisi acida generale
donazione di un protone all’atomo
di O ⇒ riduzione del carattere
carbanionico dello stato
di transizione
⇒ aumento velocità reazione
Meccanismi di Catalisi
Catalisi acido-base
•
Catalisi basica generale
l’energia libera dello stato di transizione di una reazione viene abbassato
a seguito della sottrazione temporanea di un protone da parte di una base
Esempio: tautomerizzazione cheto-enolica
reazione non catalizzata
catalisi basica generale
Meccanismi di Catalisi
Catalisi acido-base
•
reazioni acido-basiche catalizzate in modo concertato
reazioni soggette contemporaneamente a catalisi acida e basica generale
•
le catene laterali di Asp, Glu, His, Cys, Tyr e Lys hanno un valore di
pK nell’ambito del pH fisiologico
⇒ possono agire come catalizzatori acidi e/o basici
•
l’attività catalitica di questi enzimi è sensibile al pH (influenza sullo
stato di protonazione delle catene laterali )
⇒ andamento a campana della velocità della
reazione catalizzata in funzione del pH
⇒ molti enzimi sono attivi nell’ambito di un
ristretto valore di pH (5-9)
Meccanismi di Catalisi
Catalisi acido-base
•
un gruppo ionizzabile sarà più efficiente nel trasferire protoni a pH
prossimi al suo pK
tuttavia
il pK di un gruppo può variare di diverse unitàda quello atteso, a seconda
del microambiente in cui si trova
•
il pH, oltre che sullo stato di ionizzazione delle catene laterali, ha
influenza anche su:
- il legame del substrato all’enzima
- la ionizzazione del substrato
- le variazioni strutturali dell’enzima
Meccanismi di Catalisi
Residui implicati nella catalisi acido-base
pKa
~4
~10-12.5
~8.3
~6
~10
Meccanismi di Catalisi
Esempio di reazione acido-base concertata
RNasi A (pancreas bovino)
•
enzima digestivo (secreto dal pancreas nell’intestino tenue)
•
idrolisi di RNA nei nucleotidi componenti
•
velocità di reazione pH-dipendente ⇒ coinvolgimento di 2 residui
ionizzabili con pK 5.54 e 6.4
(2 residui His)
RNasi S (forma attiva, legame
peptidico 20-21 idrolizzato) +
analogo del substrato non digeribile
Meccanismi di Catalisi
Esempio di reazione acido-base concertata
Reazione 1:
His 12 (base generale) sottrae un protone dal gruppo ossidrile 2’ dell’RNA,
promuovendo l’attacco nucleofilico all’atomo di fosforo
His 119 (acido generale) promuove la scissione del legame protonando il gruppo
uscente ⇒ formazione di un intermedio poliribonuleotidico contenente un gruppo
fosfodiestere 2’,3’-ciclico all’estremità 3’-terminale
Reazione 2: dopo che il gruppo uscente si allontana entra H2O
His 12 (acido generale), His 119 (base generale)
Meccanismi di Catalisi
Catalisi covalente
•
la reazione viene accelerata mediante la formazione transitoria di un
legame covalente fra il catalizzatore (gruppo nucleofilico) ed il substrato
(gruppo elettrofilico) ⇒ “catalisi nucleofilica”
3 tappe concettuali:
1. formazione del legame covalente catalizzatore-substrato
mediante reazione nucleofilica
2. perdita di elettroni dal centro di reazione ad opera del
catalizzatore (ora elettrofilico)
3. eliminazione del catalizzatore (reazione inversa della 1.)
Meccanismi di Catalisi
Catalisi covalente
•
la nucleofilicità di una sostanza è correlata alla sua basicità
⇒ la catalisi covalente assomiglia a quella basica salvo il fatto che si
forma un legame covalente invece della privazione di un H+ dal substrato
•
più stabile è il legame covalente più sarà difficile romperlo e proseguire
nella catalisi
⇒ un “buon catalizzatore covalente” deve combinare 2 proprietà
apparentemente contraddittorie:
(a) alta nucleofilicità
(b) capacità di formare un gruppo che abbandoni facilmente l’enzima
•
gli intermedi covalenti formano il prodotto desiderato reagendo in un
secondo passaggio della reazione con:
(a) una molecola di acqua
(b) un secondo substrato
Meccanismi di Catalisi
Catalisi covalente
Gruppi nucleofilici:
- carichi negativamente
- contengono coppie di e- non condivisi in grado di formare facilmente
legami covalenti con centri poveri di elettroni
Esempi:
Proteine
gruppo amminico non protonato (Lys),
gruppo carbossilico (Asp),
gruppo imidazolico (His),
gruppo tiolico (Cys),
gruppo ossidrilico (Ser)
Coenzimi
tiamina pirofosfato (fermentazione alcolica)
piridossal fosfato (deamminazione a.a.)
Meccanismi di Catalisi
Catalisi covalente
Centri elettrofilici dei substrati:
-
gruppi carichi positivamente
guscio elettronico di valenza non completo
atomi elettronegativi
Esempi:
gruppo fosforico,
gruppo acilico,
gruppo glicosilico
Meccanismi di Catalisi
Catalisi favorita da ioni metallici
•
~ 1/3 degli enzimi richiede ioni metallici per l’attività catalitica
metalloenzimi: metalli di transizione (Fe2+, Fe3+, Cu2+, Zn2+, Mn2+, Co2+)
legati saldamente all’enzima ⇒ ruolo catalitico
enzimi attivati da metalli: metalli alcalini, alcalini-terrosi (Na+, K+, Mg2+, Ca2+)
legati debolmente all’enzima ⇒ ruolo strutturale
•
3 diversi possibili ruoli catalitici:
1) si legano al substrato orientandolo correttamente per la reazione
2) partecipano a reazioni di ossido-riduzione mediante cambiamento reversibile
del loro numero di ossidazione
3) stabilizzano elettrostaticamente o proteggono le cariche negative (o l’aumento
di densità elettronica) che possono svilupparsi durante la reazione
Meccanismi di Catalisi
Catalisi favorita da ioni metallici
- in molte reazioni si comportano come un protone che neutralizza una carica
negativa, ma sono più efficienti perché possono essere più concentrati a pH
neutro (dove [H+] = 10-7M) e con carica > +1.
- rendono più acide le molecole d’acqua ad essi legate, cioè le rendono fonte di
ioni idrossido OH− nucleofilici anche a valori di pH minori della neutralità
Esempio: Anidrasi carbonica
CO2 + H2O
HCO3− + H+
Meccanismi di Catalisi
Catalisi favorita da effetti di prossimità e orientamento
I reagenti devono venire in contatto nella giusta relazione spaziale
1) gli enzimi portano i substrati in contatto con i gruppi catalitici e, in
reazioni a più substrati, i substrati fra loro
⇒ aumento velocità di un fattore ~5
2) gli enzimi legano i substrati nelle orientazioni più produttive affinché
la reazione abbia luogo
⇒ aumento velocità di un fattore ~100
(diversa reattività delle molecole a seconda della loro orientazione.
Meccanismi di Catalisi
Catalisi favorita da effetti di prossimità e orientamento
3) gruppi carichi possono aiutare a stabilizzare lo stato di transizione
della reazione (“catalisi elettrostatica”) e possono guidare substrati
polari verso il sito attivo
4) gli enzimi stabilizzano/bloccano i movimenti (traslazionali e
rotazionali) relativi tra i substrati ed i gruppi catalitici (nello stato di
transizione) ⇒ aumento di velocità di un fattore fino a ~107)
Penalizzazione entropica:
diminuzione di entropia dovuta all’avvicinamento reciproco ordinato di
substrati e gruppi catalitici con formazione di orientamento reattivo
⇒ l’energia libera necessaria per superare tale diminuzione di entropia è
fornita dall’energia di legame del(i) substrato(i) all’enzima (valori
negativi di ΔΗ)
Meccanismi di Catalisi
Catalisi favorita dal legame preferenziale dello stato di
transizione
- un enzima può legare lo stato di transizione con una affinità maggiore
rispetto a quella dei suoi substrati e prodotti
⇒ ne aumenta la concentrazione e quindi proporzionalmente la velocità
di reazione
-
gli enzimi forzano i substrati verso la geometria dello stato di
transizione mediante siti di legame nei quali i substrati non distorti non
si legano perfettamente
ΔΔG‡cat = efficienza del catalizzatore
= ΔG‡noncat -ΔG‡cat = ΔG‡N -ΔG‡E
exp(ΔΔG‡cat /RT) = aumento di velocità
di reazione
Meccanismi di Catalisi
Catalisi favorita dal legame preferenziale dello stato di
transizione
aumento di velocità di 106 volte a 25 °C
⇒ ΔΔG‡ ~ 34 kJ/mol
se lo stato di transizione viene stabilizzato
con soli 2 legami a H in più, non presenti
fra enzima e substrato
⇒ affinità di legame per lo stato di transizione 106 volte > che al substrato
- un buon substrato non deve necessariamente legarsi con alta affinità al suo
enzima, ma lo farà dopo essere stato attivato a stato di transizione
- gli analoghi dello stato di transizione sono potenti inibitori dell’enzima