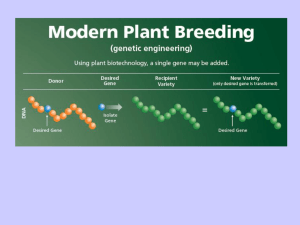
Trasformazione stabile e trasformazione transiente
Trasformazione stabile
Trasformazione transiente
Produzione di piante transgeniche:
Passaggi essenziali
Isolamento del gene di ‘interesse’ (es: inserimento in un
plasmide)
Preparazione di un costrutto per la trasformazione
genetica
Inserimento del costrutto nel tessuto vegetale
(Trasformazione genetica)
Selezione delle piante trasformate (uso dei marcatori di
selezione)
Rigenerazione del tessuto vegetale a pianta intera (PGM)
1
•Metodo Biologico
•Metodo Fisico
2
A. tumefaciens è efficiente ma ‘limitato’
alle dicotiledoni
Tra le più importanti piante coltivate:
frumento, riso, mais e orzo
Sono stati sviluppati metodi alternativi
per le monocotiledoni
Metodi Fisici di trasformazioni genetica negli Eucarioti
3
Metodo biolistico
Microparticelle d’oro (A) e di tungsteno (B)
Metodo biolistico
Particelle d’oro
ricoperte di DNA
Disco di plastica
su cui vengono
distribuite le
particelle d’oro
ricoperte di DNA
4
Metodo biolistico
Metodo biolistico
5
6
Agrobacterium
Un ingegnere genetico naturale
7
Agrobacterium spp sono batteri del suolo Gram (-)
responsabili di numerose malattie delle piante
A. tumefaciens - crown gall disease – galla del colletto
rose
grapevine
A volte il
tumore può
essere molto
esteso!
8
Come l’Agrobacterium causa la malattia?
Un megaplasmide (Ti plasmid -Tumour inducing,
circa 140 kb) contenuto nell’ A. tumefaciens è
essenziale per lo sviluppo della malattia
Induzione di tumori da parte dell’A. tumefaciens
Le cellule tumorali acquisiscono un gene per la sintesi di citochinine (e auxina)
Cellula della pianta
9
Infezione e formazione della galla
1. Riconoscimento pianta ospite-Agrobacterium
2. Ancoraggio di Agrobacterium sulla pianta ospite
3. Trasferimento e integrazione del T-DNA alla pianta
ospite
1. Riconoscimento pianta ospite-Agrobacterium
Normalmente Agrobacterium attacca la pianta ospite a
livello di una ferita fresca.
Il ferimento determina:
a) Il rilascio di composti fenolici associati alla parete
cellulare vegetale (induttori della virulenza) che
Agrobacterium percepisce e risponde attivando i geni
necessari al trasferimento (geni vir).
b) La formazione di una zona di cellule che si dividono
attivamente, in quanto il tessuto ferito va incontro ai
meccanismi di riparo.
10
I composti fenolici ‘induttori vir’
sono prodotti del metabolismo
fenilpropanoide;
Essi sono correlati a molecole che
vengono usate per rinforzare la
matrice polisaccaridica della
parete - es. cross-linkers fenolici
e lignina polimerica
acetosiringone
L’induzione vir è anche aumentata dalla presenza di
zuccheri semplici e da un pH acido – entrambi indicativi di
danno cellulare
2. Ancoramento di Agrobacterium sulla pianta ospite
Agrobacterium deve attaccarsi fisicamente alla parete
cellulare della pianta prima di poter trasferire il suo TDNA.
Sulla base dello studio di mutanti “attachment
defective”, l’attacco richiede la sintesi da parte del
batterio di un filamento polimerico di b-1,2-glucano.
Agrobacterium
b-1,2 glucano
Pianta a livello del tessuto ferito
Il target per b-1,2glucano sulla superficie
della pianta non è stato
ancora identificato.
11
Una volta stabilito il contatto iniziale, il batterio
elabora una rete di fibrille di cellulosa (b -1,4-glucano)
che aiutano il batterio ad ancorarsi alla parete cellulare
della pianta ospite.
3. Trasferimento e integrazione del T-DNA
alla pianta ospite
E’ la fase più complessa e coinvolge numerose molecole il
cui ruolo non è stato ancora completamente chiarito
12
Plasmide Ti
3 importanti regioni per il trasferimento del DNA e
l’induzione del tumore
T-DNA
LB
1. T-DNA
RB
E
2. T-DNA borders
3. Regione Vir
Oncogeni e
Opina sintasi
D
C
G
vir
B
Ti
A
Catabolismo
opine
ori
1. T-DNA
contiene geni per la biosintesi di fitormoni
(auxina e citochinina)
contiene geni per la biosintesi delle opine
•questi geni vengono trascritti soltanto nella pianta
Opine
coniugati di amminoacidi e acidi organici/zuccheri
normalmente non presenti nelle piante
Possono essere metabolizzate dall’ Agrobacterium ma
non dalla pianta
nopalina = arginina + alpha-chetoglutarato
octopina = arginina + piruvato
mannopina = glutammina + mannosio
agrocinopina A = saccarosio + arabinosio
13
reazione
della
nopalina
sintasi
reazione
della
octopina
sintase
2. T-DNA borders
•Sequenze di 24-25 bp a ripetizioni dirette
su entrambi i lati del T-DNA
•soltanto il bordo destro sembra essere
essenziale
14
3. Regione vir
•Regione di ~35 kb contenente 6-9 unità
trascrizionali (virA, B, C, D, E, F, G, H, J)
•Alcuni loci vir hanno ORFs multiple (es. virB ha
11 ORF)
• ~23 potenziali proteine sono codificate
•Alcuni prodotti dei geni vir agiscono nella cellula batterica
mentre altri sono trasportati e agiscono nella cellula ospite
•Quattro loci vir sono essenziali per la formazione del
tumore (A, B, D and G)
•La maggior parte dei geni vir sono silenti in assenza
dell’ospite, ma la virA e vir G sono trascritti
costitutivamente a bassi livelli
•Sistema di controllo gerarchico - virA e virG regolano
l’espressione degli altri geni vir
15
•VirA è associata con la membrana batterica interna, in
un complesso con il prodotto del gene cromosomico ChvE
INNER MEMBRANE
CITOPLASM
PERIPLASM
LINKER
KINASE
COOH
SENSOR
DOMAIN
NH2
Schema della proteina VirA
RECEIVER
by E.C.
•Il legame di una apposita molecola di induttore fenolico
(es. acetosiringone) con il complesso VirA causa
l’attivazione della sua latente attività chinasica, e quindi la
VirA si autofosforila
(Questa risposta della VirA è aumentata dal legame di uno zucchero
con il prodotto del gene ChvE)
•La VirA-fosforilata recluta e attiva la VirG
fosforilandola
•La VirG fosforilata è un attivatore trascrizionale per
gli altri loci vir
16
zucchero
1.
Composto fenolico
ChvE
VirA
3.
2.
ADP
P
ATP
autofosforilazione
VirA attiva
DNA genomico
4.
VirG attiva
P
VirG inattiva
P
ATP
ADP
LB
RB
E
D
P
5.
C
G
B
A
vir
Ti
ori
17
•I prodotti genici virD (VirD1 e VirD2) si legano al
bordo destro e tagliano un filamento all’estremità 5’
del T-DNA;
-Questa attività è aumentata dai prodotti
genici virC che si legano alle vicine sequenze
enhancer.
18
LB
overdrive
RB
5’
5’
Induzione vir
LB
overdrive
RB
D2
D1
C1
C2
LB
overdrive
RB
nick
D1
D2
•La proteina VirD2 rimane attaccata covalentemente
all’estremità 5’ del filamento di T per tutto il processo di
trasferimento
•La proteina VirE2 è una ‘single-stranded DNA
binding protein’; più copie si legano al filamento T.
Anche la virE2 passa nella cellula ospite insieme al
filamento.
LB
RB
E1
overdrive
Sintesi di DNA
E1
E1
D1
D2
19
E1 E1 E1 E1 E1 E1 E1
E1 E1 E1 E1 E1 E1 E1
D2
Filamento T con la proteina “cap” VirD2 attaccata covalentemente
Questa associazione può già avvenire nella cellula batterica,
ma alcune evidenze suggeriscono che il filamento T inciso + D2
È trasferito all’ospite, separatamente dalle proteine E1, e il
filamento ricoperto di E1 si forma nel citoplasma della cellula
ospite
•Il prodotto del gene VirB contribuisce nella
formazione di una struttura di secrezione di “tipo IV”,
che potrebbe coivolgere il pilus batterico come
condotto o struttura guida
20
Fig. 1. The six functions of VirE2. The vir regulon, encoding the major loci virA-E and virG-H, is expressed on detection of plant wound
signals. VirD2 and VirD1 liberate the T-strand and VirD2 remains covalently bound to the 5' end. 1: VirE2 coats the T-strand, protects it
from degradation, and maintains it in a transportable conformation. 2: VirE2 associates with VirE1, required for VirE2 export. 3: VirE2
exits Agrobacterium via the type IV exporter independently, or as part of the T-complex. Alternatively, VirE2 may exit by an alternate
pathway (pink; ref. 4). 4: VirE2 forms a pore in the plant plasma membrane allowing passage of the T-complex and coats the T-strand in
the plant cytoplasm. 5: VirD2 and VirE2 interact with plant cytoplasmic chaperones (RocA, Roc4, and CypA). Other factors (AtKAP ,
VIP1) may target the T-complex to the nucleus. 6: VirE2 interacts with nuclear factors (VIP2) that mediate interaction with chromatin and
facilitate integration of the T-strand.
Altri prodotti dei geni vir
VirF - fattore che condiziona il range delle piante
ospiti; potrebbe interagire con proteine nucleari
che regolano il ciclo di divisione cellulare?
VirH - (PinF) possibile citocromo P-450 idrossilasi
funzione di difesa? Fattore per il range d’ospite?
VirJ - possibile binding protein del filamento T; aiuta
l’esportazione del filamento T?
21
Formazione del tumore
•Il T-DNA del ceppo wild-type di Agrobacterium
codifica due tipi di funzioni- produzione di fitormoni e
sintesi di opine.
•I geni del T-DNA hanno strutture promotrice e
‘codon usage’ che gli consente di essere trascritti
efficientemente nelle cellule vegetali ma non nei
batteri.
•La produzione di fitormoni è catalizzata da due
prodotti genici (Tms1 e Tms2) che convertono il
triptofano in acido 3-indol-acetico (IAA), e un
prodotto genico (Tmr/Ipt) che converte l’adenina in
isopentinil adenina (citochinina).
22
L-triptofano
O2
IaaM
Ammide -2-indol acetico
CO2
IaaH
NH3
Acido-3-indol acetico (IAA)
2-isopentinil-PP
adenosina - 5’-PO4
Tmr1
Isopentinil adenine-5-PO4
PPi
Non tutti I geni del T-DNA sono stati
caratterizzati.
ORF 6b, per esempio è stato soltanto recentemente
riportato codificare una proteina che ha le
caratteristiche di un fattore di trascrizione
eucariotico (DNA-binding domain)
A. rhizogenes trasporta un differente megaplasmide
(Ri) ma possiede la stessa funzionalità.
Il T-DNA di Ri sembra mancare di un gene per la
biosintesi delle citochinine, che probabilmente è
responsabile della morfologia ‘radicante’ del tumore
indotto dall’ A. rhizogenes.
23
E’ interesante notare che geni omologhi al T-DNA
tms1 e 2, e tmr1 sono presenti in altri batteri
fitopatogeni
Pseudomonas savastanoi, per esempio, causa una
malattia che produce una galla chiamata ‘rogna
dell’olivo’. I fattori di virulenza noti comprendono
la capacità di sintetizzare IAA (codificato da
iaaM e iaaH) e citochinina (codificato da ptz).
Comunque, P. savastanoi manca della capacità di
trasferire questi geni al genoma ospite. Produce e
secerne i fitormoni al sito di infezione.
Il tessuto colpito risponde con divisione e
espansione cellulare, che determina la formazione
della galla.
N.B. Questo implica differenze nel controllo trascrizionale...
Spettro d’ospite
•A. tumefaciens ha un ampio spettro d’ospite
nell’ambito delle dicotiledoni.
•Le monocotiledoni e le gimnosperme non sono di solito dei
buoni ospiti
•La mancanza di infettività può essere dovuto:
-alla non produzione di un appropriato composto
fenolico induttore (es. la colonizzione avviene se
l’induttore viene fornito esogenamente)
-alla presenza di fattori dell’ospite non
ancora caratterizzati biochimicamente.
•Alcuni casi di apparente incompatibilità sono stati
superati ‘coltivando’ l’Agrobacterium con specifici tessuti
(es. piccioli cotiledonari in Brassica)
24
Altri determinanti dello spettro d’ospite possono
essere:
i) La sensibilità o specificità della VirA;
ii) altri geni vir difettosi (necessari più per alcuni
ospiti che per altri); o
iii) oncogeni difettivi (es. incapacità di produrre la
correta miscela di fitormoni nella cellula ospite)
Agrobacterium
e
Ingegneria Genetia vegetale
25
Osservazioni chiave
Il T-DNA è permanentemente incorporato nel genoma
della cellula ospite
Nessuno dei geni codificati dal T-DNA sono necessari
per il trasferimento: fitormoni, opine
Soltanto i bordi del T-DNA e le funzioni vir sono
necessari
T-DNA
LB
RB
Oncogeni e
Opina sintasi
E
D
C
G
B
A
vir
Ti
Catabolismo
opine
ori
Inoltre…..Limitazioni del plasmide Ti
-Dimensioni del plasmide troppo grandi per la manipolazione
-Produzione di auxine e citochinine previene la
rigenerazione della pianta intera
26
Due approcci per ottenere plasmidi Ti ingegnerizzati
•Sistema del Vettore binario
•Sistema del Vettore cointegrato
•Sistema del Vettore binario
1) Plasmide di piccole dimensioni
ad ampio spettro d’ospite:
•Due Ori: di E. coli e di Agrobacterium
•RB (e LB)
•Marcatori per la selezione e
mantenimento in E. coli e A. tumefaciens
2) Plasmide Ti helper
(Ti disarmato)
•Privo del T-DNA
•Contiene i geni Vir
•Marcatore di selezione per le piante
•Gene di interesse
27
•Sistema del
Vettore binario
Fig. 15.6 Watson et al DNA ricombinante
•Sistema del
Vettore binario
Fig. 15.6 Watson et al DNA ricombinante
28
•Sistema dei Vettori cointegrati
•Due plasmidi ricombinano per formare un vettore
cointegrato
-Vettore ‘spoletta’
•Ori di E. coli (no Ori di Agrobacterium)
•RB (e LB)
•Marcatori per la selezione e mantenimento in E. coli
e A. tumefaciens
•Marcatore di selezione per le piante
•Gene di interesse
•DNA omologo al plasmide Ti
-Ti helper Ti disarmato
Vettore ‘spoletta’
Plasmide Ti cointegrato
29
•Sistema del
Vettore
cointegrato
Fig. 15.5 Watson et al DNA ricombinante
•Sistema del
Vettore
cointegrato
Fig. 15.6 Watson et al DNA ricombinante
30
plasmidi Ti ingegnerizzati
•Sistema dei Vettori binari
-due plasmidi
•geni vir in un plasmide
•T-DNA in un altro plasmide
•Sistema dei Vettori cointegrati
-inizia con due plasmidi
-ricombina per avere un singolo plasmide
•Geni vir e T-DNA finiscono sullo stesso plasmide
Trasformazione con Agrobacterium: Vettori Binari
Possibili svantaggi:
Possibile instabilità del plasmide ad ampio spettro
d’ospite, dovuto alla contemporanea presenza delle
origini di replicazione per E. coli e A. tumefaciens
Rispetto ai vettori co-integrati presenta diversi
vantaggi:
•Non richiede ricombinazione tra le molecole implicate
•Utilizza vettori di dimensioni piccole molto più
efficienti dei grandi plasmidi Ti disarmati nel
trasferiemnto da E. coli ad A. tumefaciens
31
Trasformazione con Agrobacterium: Vettori cointegrati
Possibili svantaggi:
•Richiede estese regioni di omologia tra il Ti e il plasmide di
E.coli
•Trasferimento genico meno efficiente rispetto ai vettori
binari
Floral dip
transformation
I fiori di
Arabidopsis
possono
essere
semplicemente
immersi in con
una soluzione
contenente
Agrobacterium
con il
costrutto
genico
32
Con il Floral dip transformation viene completamente eliminata la
necessità di produrre tessuto callificato per la trasformazione
Trasformazione transiente
33
Espressione transiente con sistemi virali
In piante di Nicotiana benthamiana come
sistema eterologo utilizzando il virus X
della patata (PVX)
166 kd
25kd 8kd
GDI
CP
12kd
34
Agroinfezione
E’ possibile inserire il DNA di un genoma virale tra i
bordi del T-DNA di un plasmide Ti e usare il processo
di trasferimento dell’Agrobacterium tumefaciens per
incorporare il DNA virale nella cellula vegetale.
Una volta inserito, le funzioni virali possono
trascrivere e replicarsi, stabilendo un’infezione
virale.
Esempi di utilizzo:
•per studiare virus che non infettato senza l’aiuto di
vettori (afidi), es. Luteovirus
•Per migliorare l’efficienza di infezione per produrre
proteine ricombinanti
•Per identificare fattori di virulenza
Esempio di analisi funzionale mediante Agroinfiltrazione
Cold Spring Harbor Laboratory Press
Torto T A et al. Genome Res. 2003;13:1675-1685
35
Sintomi su N. benthamiana dopo agroinfiltrazione con i cDNA
crn1 e cnr2
16 dai
12 dai
PVX vector pGR106
pGR106 crn1 cDNA
pGR106 crn2 cDNA
Cold Spring Harbor Laboratory Press
Torto T A et al. Genome Res. 2003;13:1675-1685
• Il sistema PVX è stato proposto anche
come possibile soluzione al rischio di
produrre molecole di interesse
farmacologico nelle piante
36