Corso di Chimica delle Macromolecole
mmmm!!
Mi piacciono le
macromolecole a
pranzo!
parte del corso integrato di Biochimica
Aspetti organizzativi del corso di Chimica delle Macromolecole
-Testo di supporto: Campbell, Farrell "Biochimica" seconda (o terza) edizione.
EdiSES.
-Diapositive ed altro da http://biocfarm.unibo.it/samori/bioch_IF.html uno
strumento da usare durante il corso, non solo dopo.
-Ricevimento: previo appuntamento fissato per e-mail a [email protected]
o concordato durante le lezioni.
-Esame di profitto: il voto/verbalizzazione sarà unico per tutte le parti del corso
integrato. Saranno offerte prove scritte in itinere (esami parziali) riguardanti le varie
parti del corso. Il superamento delle prove parziali può sostituire l’esame finale. Gli
argomenti di questo corso (macromolecole) saranno comunque oggetto della parte
scritta della verifica, anche dopo l’esaurimento delle prove parziali.
L’acquisizione dei 13 CFU corrispondenti a BIOCHIMICA C.I. avverrà solo in seguito al
superamento dei colloqui parziali sui singoli corsi che lo compongono, che devono anche
essere sostenuti nel seguente ordine:
1. Chimica delle macromolecole (Prof. Zuccheri)
2. Biochimica (Prof. Farruggia-Fiorentini)
3. Biochimica della nutrizione (Prof. S. Hrelia)
Unità didattica: Lezione introduttiva alle
macromolecole
Oggetto della lezione:
Cosa sono le macromolecole. Introduzione ai concetti base delle macromolecole.
Le proteine, gli acidi nucleici, i lipidi, i polisaccaridi: cosa sono e a cosa servono.
Scopo della lezione:
Fornire i concetti di base per l’apprendimento dei temi del corso, anche
propedeutici agli insegnamenti degli altri docenti del C.I. e ad altri docenti del
corso di studi (corso di Microbiologia, corso di Chimica Farmaceutica).
Permettere agli studenti di rendersi conto della tipologia dei concetti trattati
durante il corso e di poter identificare quali concetti della chimica generale e della
chimica organica sono essenziali alla comprensione del corso, valutandone
eventualmente una revisione personale.
Verifica dell’apprendimento: Sapresti rispondere con facilità almeno alle
seguenti domande?
Cosa sono le macromolecole? In cosa si differenziano dalle molecole trattate dalla chimica organica
‘classica’ o dalla ‘chimica inorganica’? Cosa è un monomero? Cosa sono le proteine? A cosa servono?
Da quali unità ripetitive sono costituite? Cosa è la struttura delle proteine? Quali livelli gerarchici della
struttura delle proteine conosci? Cosa sono i lipidi? E gli acidi grassi? Come sono fatti i polisaccaridi?
Cosa sono gli acidi nucleici? Quali sono i costituenti base degli acidi nucleici? Quanti tipi di acidi nucleici
ci sono? Che struttura tridimensionale hanno?
Cosa sono le macromolecole?
•Sono molecole molto grandi
•Sono formate quando monomeri si legano per formare catene più lunghe, i
polimeri.
•Processi analoghi portano alla formazione o alla rottura dei polimeri (negli
organismi viventi).
L’etene, un monomero
Il polietilene, un polimero
Un parallelo ‘linguistico’
I polimeri stanno ai monomeri come le parole stanno alle lettere
In posizione gerarchica crescente:
segno
→
lettera →
重
parola
→
frase
→ lingua
“per correr miglior acque alza
le vele ormai la navicella del
mio ingegno.”
atomo → molecola → macromolecola → interazioni intermolecolari → cellula/vita
H
重
Alcune lettere ‘vivono’ e hanno significato come tali, senza formare parole
Le macromolecole nella cellula
I polimeri (o macromolecole) sono sostanze
ad alto peso molecolare, costituite da unità
monomeriche ripetitive
COMPOSTI MACROMOLECOLARI O POLIMERICI
MACROMOLECOLE
104
M
109
ORGANICI OD INORGANICI
NATURALI
Poliisoprene
Polisaccaridi
Acidi nucleici
Proteine
Argille
Asbesto
Quarzo
Polifosfati
SEMISINTETICI
SINTETICI
Polivinilici
Polivinilidenici
Poliesteri
Poliammidi
Poliuretani
Resine fenoliche
Polieteri
Polisilossani
Poliimmidi
Polifosfazene
Classificazione di composti macromolecolari
su basi composizionali
COMPOSTI MACROMOLECOLARI
OMOPOLIMERI
COPOLIMERI
Strutture
macromolecolari
ad una sola unità
ripetente
Strutture
macromolecolari
a più di una unità
ripetente
[A-A-(A)n-A]
COPOLIMERI
Alternati
A blocchi
Statistici
(AB)n
(A)n
ABAAABABBA
(B)m
• Proprietà ultime dei materiali strettamente dipendenti dalla
distribuzione delle unità monomeriche
Classificazione dei polimeri secondo
la struttura delle macromolecole
LINEARI RAMIFICATI RETICOLATI
Come si ottengono i polimeri in laboratorio/nell’industria
POLIMERI
POLIMERIZZAZIONE A
CATENA
lineare
ramificata
• Meccanismo di crescita
delle macromolecole a
“catena”
coinvolgente
l’apertura di doppi legami
o tripli legami o di cicli a
3-7 termini
• Meccanismo di crescita
delle macromolecole a
“stadi” con perdita di
molecole
piccole
(policondensazione) o no
(poliaddizione).
• Composizione
centesimale del polimero
coincidente con quella
del monomero
• Eteroatomi presenti nella
catena principale
• Catena principale in
genere
di
natura
poliidrocarburica
reticolata
POLIMERIZZAZIONE
A STADI
• Composizione centesimale
del polimero diversa da
quella
dei
precursori
monomerici nel caso di
policondensazione
Reazioni di condensazione
• Considerare, ad esempio, monomeri con
gruppi –OH terminale
• Questi monomeri possono essere collegati
mediante un processo chiamato
condensazione nel quale si forma un legame
covalente tra le due molecole e si perde una
molecola d’acqua.
• Questa reazione può essere catalizzata (da un
enzima, se in un sistema vivente)
• Questo stesso tipo di reazione può avvenire
per diverse classi di polimeri: proteine,
carboidrati, acidi nucleici (e polimeri sintetici).
Un esempio
La reazione di idrolisi
• Polimeri di vario tipo possono essere
depolimerizzati mediante la reazione di
idrolisi. Una molecola di acqua è
addizionata per rompere un legame
covalente (con l’aiuto di una idrolasi, un
enzima specifico, ad esempio). In questo
modo si possono rilasciare i monomeri.
Quattro tipi di macromolecole nei
viventi
•
•
•
•
Lipidi
Carboidrati
Acidi Nucleici
Proteine
30 sono i più comuni monomeri nella cellula
Molecula
Tipi
diversi
Nome della molecola
Amminoacidi
20
Ala (A)
Asp (N)
Phe (F)
Gln (Q)
Thr (T)
His (H)
Basi
aromatiche
5
Adenina Timina Citosina
Uracile Guanina
Monomeri degli acidi
nucleici
Zuccheri
2
Riboso
Componente gli acidi
nucleici
Glucoso
Metabolismo energetico
Colina
Componente i fosfolipidi
Lipidi
3
Leu (L) Arg (R) Lys (K)
Met (M) Asp (D)
Cys (C) Pro (P)
Ser (S) Glu (E)
Gly (G) Trp (W)
Tyr (Y) Ile (I) Val (V)
Ruolo
Glicerolo
Palmitato
Monomeri delle proteine
Proteine
• Più della metà del peso secco
del corpo umano.
– Polimeri degli amminoacidi
– Ricoprono diversi ruoli: enzimi
per la catalisi, elementi
strutturali, recettori di segnale,
trasportatori, …
Proteine (Polipeptidi)
•
Amminoacidi (20 tipi diversi) legati insieme da legami peptidici
(polipeptidi)
•
Sei funzioni delle proteine:
1. Riserva:
albumina (albume dell’uovo)
2. Trasporto:
emoglobina
3. Regolazione: ormoni
4. Movimento:
muscoli
5. Struttura:
membrane, capaelli, unghie
6. Enzimi:
reazioni cellulari
Gli enzimi sono i catalizzatori proteici
senza enzima
con l’enzima
Solo un effetto sulla cinetica della reazione
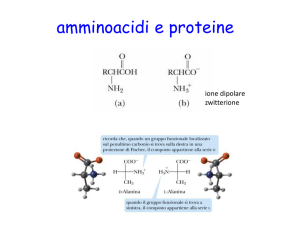
Gli amminoacidi
H
carbonio α
R
R
catena
laterale
R
Cα
+
H 3N
COO –
gruppo
gruppo
amminico carbossilico
COO –
NH 3
COO
–
modello sfere e
bastoncini
NH 3
Gli amminoacidi sono
strutture tetraedriche
Gli amminoacidi sono molecole chirali (quasi tutti)
Ecco la reazione di condensazione di due amminoacidi:
la formazione del legame peptidico
Proteine
I livelli di organizzazione che determinano la struttura
Livello di struttura
Base della struttura
Legami coinvolti
Primaria
Sequenza degli
amminoacidi
Legami covalenti
Secondaria
RIpiegamento in αeliche o foglietti β o
come gomitolo
statistico
Legami idrogeno
Terziaria
Ripiegamento
tridimensionale di una
catena polipeptidica
Legami idrogeno e
ponti disolfuri,
interazioni
elettrostatiche e
idrofobiche
Quaternaria
Associazione di 2 o più
subunità ripiegate
Come per la terziaria
Livelli della struttura proteica I
• Primaria (1o)
– Sequenza degli amminoacidi
– Determinati dall’informazione genetica
– Tenuti insieme da legami covalenti peptidici
• Secondaria (2o)
– Ripiegamento delle regioni dei polipeptidi
– α-eliche
o foglietti β
– Tenuti insieme da deboli legami idrogeno
Livelli della struttura proteica II
• Terziaria (3o)
– Ripiegamento della proteine intera in una forma
caratteristica
– Può essere globulare (enzimi o fibrosa (capelli)
– Può essere bloccata in una forma da ponti disolfuro
• Quaternaria (4o)
– Interazione di 2 o più polipeptidi. Ad esempio,
l’emoglobina, la proteina che trasporta l’ossigeno nel
sangue è fatta di 4 polipeptidi tenuti insieme
Proteine
• Struttura primaria
– Derivata dall’informazione genetica
– La sequenza degli amminoacidi contiene
l’informazione appropriata per il ripiegamento
corretto della proteine
– Una sostituzione singola può risultate in una
proteina che non si ripiega in modo corretto
(come succede per l’emoglobina)
Struttura primaria
• Scritta dal terminale amminico a quello
carbossilico
• Codificata dalla sequenza nucleotidica
dell’mRNA, con i ‘comandi’ contenuti nel
DNA
• La sequenza determina le strutture
secondaria, terziaria e quaternaria, e
anche le interazioni proteina-proteina e
proteina-acidi nucleici
Proteine
• La struttura secondaria
– Coinvolge interazioni relativamente locali tra gli amminoacidi
– Le interazioni portano alla formazione di eliche o foglietti β
– L’α-elica ha 3.6 amminoacidi per spira e questo porta in
prossimità gli amminoacidi ad un intervallo di 4
– Leu, Met e Glu hanno una propensione a formare eliche
– Gly e Pro interrompono l’elica e sono coinvolti in
ripiegamenti e deformazioni delle eliche
– Si formano legami idrogeno tra l’azoto imminico di un
amminoacido ed il carbonile di un altro amminoacido
Un modello di α-elica
Proteine
• Il foglietto β
– Massimizza i legami idrogeno, collegando due
polipeptidi o due sementi diversi di un
polipeptide
– Ile, Val e Phe hanno una certa propensione a
formare foglietti
– I foglietti possono essere paralleli (N->C dei
polipeptidi allineati)i
I foglietti beta
È un’altra struttura secondaria stabile e molto diffusa delle proteine, stabilizzata dalla
formazione cooperativa di un grande numero di legami idrogeno. È della foglietto
ripiegato β o struttura β.
Nella struttura, i carboni α stanno nelle pieghe delle ‘strisce’, i C=O puntano in una
direzione e gli N-H che li legano nella direzione opposta. Tutti i gruppi formano legami H.
Antiparallel β-sheet
La ribonucleasi A bovina (bovine ribonuclease A) è una piccola proteina
(14.6 kD, 129 residui) che contiene alcune eliche corte, una sezione
importante di β-sheets antiparalleli e alcuni ripiegamenti β, oltre ad alcuni
segmenti senza struttura definita.
Proteine
• Ci sono motivi strutturali ricorrenti (associazioni di
strutture secondarie)
– β−α−β
– Hairpin-turn
– Helix-turn-helix
Proteine
• Struttura terziaria
– Dipende più dai gruppi R che dai gruppi
immino e carbonile
– I domini idrofobici tendono ad associare gli
uni con gli altri e sono spesso ritrovati nelle
zone interne delle proteine
– I domini idrofobici ed idrofilici determinano la
struttura “nativa,” che può formarsi
spontaneamente o grazie all’aiuto degli
Chaperon
Varie interazioni concorrono a stabilizzare la struttura terziaria
Come per la trioso fosfato isomerasi, le proteine possono presentare anche
strutture molto complesse e simmetriche.
(d) T rioso fosfato isomerasi
La natura ricicla, quando può: similitudini di struttura globale non devono fare
pensare a similitudini di funzione per le proteine.
Proteine
• Stabilizzazione della struttura terziaria
– Legami non covalenti
• Legami idrogeno tra gruppi R opportuni
• Interazioni elettrostatiche tra gruppi R carichi
• Interazioni idrofobiche tra gruppi R non polari
– Legami covalenti
• Ponti disolfuro tra due residui di Cys
• I legami disolfuro sono importanti per stabilizzare sia la
struttura terziaria che quella quaternaria
Reticolazione mediante legami disolfuro
L’insulina è un esempio di polipeptide ricco in disolfuro
Insulina
Proteine
• Fibrose
– Hanno una struttura secondaria molto estesa, con
eliche o foglietti molto lunghi
– Esempi: Fibroina, cheratina, collagene, elastina
• Globulari
– La maggior parte delle proteine coinvolte nelle
funzioni cellulari
– Le eliche ed i foglietti sono presenti e permettono il
ripiegamento della catena polipeptidica
Le strutture cellulari hanno spesso
un’organizzazione gerarchica
ottenuta mediante interazioni tra
catene diverse
Proteine
• Molte proteine globulari consistono di diversi
domini
• Un dominio è una unità discreta di struttura
terziaria ripiegata indipendentemente
– 50-350 residui amminoacidici
– Spesso ogni dominio ha una funzione precisa nella
proteina
– Ad esempio, certi enzimi hanno domini in grado di
legare gli ioni metallici
Proteine
• Domini
– Le proteine globulari piccole hanno
solitamente un solo dominio
– Le grandi proteine globulari hanno più di un
dominio
Singoli domini di proteine globulari
Proteine
• Struttura quaternaria
– È il livello di organizzazione correlato
all’interazione e all’assemblaggio delle
subunità
– Esempio: le Integrine (recettori nella matrice
extracellulare) contengono subunità α e β
Alpha 2 beta 1 integrin domain 1
IgG Fab
Acidi Nucleici
• Immagazzinano, trasportano e agevolano
la trasmissione dell’informazione genetica
• Sono polimeri lineari dei nucleotidi
• Differiscono tra loro nel tipo di zucchero
che contengono (sempre il pentoso
chiamato riboso)
– Acido desossiribonucleico (DNA)
– Acido ribonucleico(RNA)
Acidi nucleici
• Acido desossiribonucleico
– L’idrogeno sostituisce l’ossidrile al 2’
– Quattro basi nucleotidiche (ATCG)
– Il gruppo fosfato
• Acido ribonucleici
– Riboso
– Quattro basi nucleotidiche (AUCG)
– Il gruppo fosfato
Il ruolo e le forme degli acidi nucleici
RNA
Nucleoside1
Nucleotide2
•Adenina
Adenosina
AMP
•Guanina
Guanosina
GMP
•Citosina
Citidina
CMP
•Uracile
Uridina
UMP
Base
Purine
Piyrimidine
1base
+ zucchero
2base
+ zucchero + fosfato
DNA
Nucleoside1
Nucleotide2
•Adenina
Desossiadenosina
dAMP
•Guanina
Desossiguanosina
dGMP
•Citosina
Desossicitidina
dCMP
•Timina
Timidina
dTMP
Base
Purine
Pirimidine
1base
+ zucchero
2base
+ zucchero + fosfato
Polimeri
• Polimeri lineari si formano mediante il
legame fosfodiestere
– Il fosfato è legato mediate un fosfoestere al
carbonio 5’ di un nucleotide o
desossinucleotide
– E forma un legame con il carbonio 3’ del
nucleotide o desossinucleotide successivo
– Si forma un fosfodiestere 3’, 5’
• Per la formazione è necessaria l’energia
fornita da un nucleotide trifosfato
Le catene di DNA e
RNA: questa è una
descrizione delle
connessioni, non
della geometria
dell’elica
Acidi nucleici
• Il riconoscimento tra le molecole dipende
dalle caratteristiche di purine e pirimidine
– Gruppi carbonili ed azoti sono capaci di
formare legami idrogeno
– I legami idrogeno si formano tra le coppie A:T
o A:U o G:C
Le catene di DNA e
RNA: questa è una
descrizione delle
connessioni, non
della geometria
dell’elica
La duplicazione del DNA: la
catena del DNA è costituita in
modo tale da i) avere ridondanza
nell’informazione contenuta e ii)
essere intrinsecamente adatta
per essere duplicata, facendo
essa stessa da stampo per le
catene nuove che è necessario
sintetizzare
DNA
• Gli esperimenti di diffrazione su fibra
hanno mostrato che il DNA può esistere in
varie conformazioni. La conformazione più
comune in soluzione è detta B-DNA. Il BDNA è un’elica destrogira con un solco
stretto ed uno largo. Le basi sono
perpendicolari all’asse molecolare. È la
geometria più comune.
B-DNA
B-DNA
DNA
• Il DNA si può anche trovare nella forma A
in cui il solco maggiore è molto profondo
ed il solco minore molto superficiale
• Una forma insolita è quella del DNA Z. I
due nucleotidi che costituiscono l’unità
strutturale base di questa forma hanno
diversa conformazione ed impartiscono un
andamento a zig-zag (globalmente
levogiro) a questa elica.
A-DNA
Z-DNA
RNA
• L’RNA è un polimero che contiene riboso
invece che desossiriboso. La
composizione normale delle basi contiene
guanina, adenina, citosina, uracile
• Anche l’RNA può formare catene a doppia
elica. Queste sono nella conformazione A
poiché la presenza dell’OH in 2’ non
consente la conformazione B. Ma l’RNA è
più comunemente a catena singola e può
assumere strutture molto inusuali e strane.
RNA
• Ad esempio l’RNA transfer è una molecola
chiave per il processo di traduzione del
materiale genetico nelle proteine. È
costituito di circa 70 basi ripiegate per
formare steli con basi appaiate e anelli
spaiati. La forma globale di un tRNA può
essere assimilata a quella della lettera L.
Double stranded RNA
tRNA
t-RNA
siRNA vegetale legato ad un suo repressore virale
Complessi tra acidi nucleici e proteine
Carboidrati
Formula: (CH2O)n
I
or H - C - OH
I
Monosaccaridi – zuccheri semplici con gruppi H multipli.
In funzione del numero di atomi di carbonio, (3, 4, 5, 6),
un monosaccaride si dice trioso, tetroso, pentoso, esoso.
Disaccaridi - 2 monosaccaridi legati covalentemente: un
dimero.
Oligosaccaridi – alcuni monosaccaridi legati
covalentemente.
Polisaccaridi – polimeri che consistono di catene di
monosaccaridi o disaccaridi.
Struttura dei monosaccaridi
• Un ossidrile è attaccato ad ogni
carbonio, ad eccezione di uno, quello
del carbonile (doppio legame C=O)
Carboidrati
funzione:
Come deposito di energia
- Come deposito intermedio (amido per
le piante, glicogeno per gli animali)
- Come componente strutturale delle
cellule (cellulosa nella parete cellulare
delle piante e chitina nell’esoscheletro
di insetti e artropodi.
Monosaccaridi
Aldosi (es: glucoso) hanno Chetosi (es: fruttoso)
un gruppo aldeidico ad una hanno un carbonile al C2
estremità
H
C
O
CH2OH
C
O
HO
C
H
OH
H
C
OH
OH
H
C
OH
H
C
OH
HO
C
H
H
C
H
C
CH2OH
CH2OH
D-glucose
D-fructose
D vs. L
La designazione
degli isomeri D e L
è basata sulla
configurazione del
carbonio
asimmetrico della
gliceraldeide.
Proiezioni di Fischer
D=R
CHO
CHO
H
C
OH
HO
L-glyceraldehyde
CHO
H
C
OH
CH2OH
D-glyceraldehyde
H
CH2OH
CH2OH
D-glyceraldehyde
C
CHO
HO
C
H
CH2OH
L-glyceraldehyde
Zuccheri D e L
Per gli zuccheri con più
di un centro chirale, la
designazione D o L si
riferisce al carbonio
asimmetrico più lontano
dal gruppo aldeidico o
chetonico.
La maggioranza degli
zuccheri naturali è D.
O
H
C
H – C – OH
HO – C – H
O
H
C
HO – C – H
H – C – OH
H – C – OH
HO – C – H
H – C – OH
HO – C – H
CH2OH
D-glucose
Gli zuccheri D e L sono enantiomeri (imagini speculari)
CH2OH
L-glucose
Nomenclatura degli zuccheri
•Gli zuccheri D e L hanno lo stesso nome es: Dglucoso e L-glucoso.
•Gli altri stereoisomeri hanno nomi caratteristici.
•Ci sono 2n stereoisomeri, ove n è il numero di
stereocentri.
•Gli aldosi 6-C hanno 4 carboni asimmetrici per
cui ci sono 16 stereoisomeri (8 zuccheri D e 8 L).
Formazione di acetali e ketali
Un aldeide può
reagire con un
alcol per formare
un emiacetale
Un chetone può
reagire con un
alcol per formare
un emichetale
H
C
H
O
+
R'
OH
R'
O
R
OH
R
aldehyde
alcohol
hemiacetal
R
C
C
R
O
+
"R
OH
R'
ketone
"R
O
C
R'
alcohol
hemiketal
OH
I pentosi e gli
esosi possono
ciclizzare se i
carbonili
reagiscono con
un OH distale.
Il glucoso forma
un emiacetale
intramolecolare,
un anello
piranosico a 6
membri.
1CHO
H
2
C
HO 3C
H
OH
H
D-glucose
C
4
OH
H 5C
OH
(linear form)
6 CH2OH
6 CH2OH
6 CH2OH
5
H
4
OH
H
OH
3
H
O
H
H
1
2
OH
α-D-glucose
OH
5
H
4
OH
H
OH
3
H
O
OH
H
1
2
OH
β-D-glucose
Gli zuccheri ciclici sono spesso presentati come
proiezioni di Haworth.
H
6 CH2OH
6 CH2OH
5
H
4
OH
O
H
OH
3
H
H
2
OH
α-D-glucose
H
1
OH
5
H
4
OH
H
OH
3
H
O
OH
H
1
2
H
OH
β-D-glucose
La ciclizzazione produce un nuovo stereocentro
al C1. I 2 stereoisomeri sono chiamati anomeri,
α e β.
Nelle proiezioni di Haworth:
α (OH sotto l’anello)
β (OH sopra l’anello).
Legami glicosidici
L’ossidrile anomerico ed un ossidrile di un altro zucchero
possono condensare (perdendo acqua o altra molecola
piccola) e formare un legame glicosidico:
R-OH + HO-R' Æ R-O-R' + H2O
Es., Il metanolo può reagire con l’OH e fare un metil
glucoside.
H OH
H OH
H2O
H O
HO
HO
H
H
OH
H
+
CH3-OH
HO
HO
H
H
OH
α-D-glucopyranose
H O
methanol
H
OH
OCH3
methyl-α-D-glucopyranose
Disaccaridi:
Maltoso, un prodotto di
scissione dell’amido, è
un disaccaride con un
legame glicosidico
α(1Æ4) tra C1-OH e
C4-OH di 2 molecole di
glucoso. Questo
significa che è
coinvolto l’anomero α.
6 CH2OH
6 CH2OH
H
4
O
5
H
OH
OH
3
H
H
H
1
H
4
4
maltose
OH
H
H
H
H
H
1
OH
2
OH
O
1
4
5
O
H
OH
H
H
3
H
6 CH2OH
O
H
OH
H
OH
3
OH
5
O
O
2
6 CH2OH
H
5
2
OH
3
cellobiose
H
2
OH
1
H
OH
Cellobioso, un prodotto della scissione della cellulosa.
L’anomero β (O su C1 verso l’alto) lega l’OH sul C-4. Il legame
glicosidico β(1Æ4) è rappresentato ella proiezione come un zigzag, ma si tratta ovviamente di un artificio grafico (il secondo
glucoso è ribaltato rispetto al primo).
Disaccaridi
Altri disaccaridi comuni:
Saccaroso, lo zucchero da tavola, ha un legame
glicosidico tra gli ossidrili anomerici di glucoso e
fruttoso.
La configurazione del glucoso è α (O verso il
basso), quindi il legame è detto α(1Æ2).
Il nome completo del saccaroso è α-Dglucopiranosil-(1Æ2)-β-D-fruttopiranoso.)
Lattoso, lo zucchero del latte, composto di
galattoso e glucoso, con un legame β(1Æ4)
dell’ossidrile anomerico del galattoso. È il β-Dgalattopiranosyl- (1Æ4)-α-D-glucopiranoso
Glucoso di riserva nelle piante
CH 2OH
H
O
H
OH
H
OH
H
OH
H
H
1
O
6 CH OH
2
5
O
H
4 OH
H
3
H 1
H
H
O
O
H
OH
CH 2OH
CH2OH
CH2OH
H
H
H
O
O
H
OH
H
2
OH
H
OH
H
OH
H
H
O
O
H
OH
H
H
OH
H
amylose
Le piante immagazzinano glucoso come amilosio o
amilopecina, polimeri del glucoso chiamati collettivamente
amido. Immagazzinare glucoso in una forma polimerica
minimizza di effetti osmotici.
Amiloso: è un polimero con legami α(1Æ4).
Il terminale di un polisaccaride non coinvolto in un legame
glicosidico è detta terminate riducente.
L’amiloso ha una conformazione ad elica.
OH
CH2OH
CH2OH
O
H
H
OH
H
H
OH
H
O
OH
CH2OH
H
H
OH
H
H
OH
H
H
OH
CH2OH
O
H
OH
O
H
OH
H
H
O
O
H
OH
H
H
OH
H
H
O
4
amylopectin
H
1
O
6 CH2
5
H
OH
3
H
CH2OH
O
H
2
OH
H
H
1
O
CH2OH
O
H
4 OH
H
H
OH
H
H
O
O
H
OH
H
H
OH
H
OH
Amilopectina è un polimero con principalmente legami
α(1Æ4), ma anche ramificazioni del tipo α(1Æ6). Le
ramificazioni producono una struttura compatta e molti
terminali ai quali possono avvenire reazioni
enzimatiche.
CH2OH
CH2OH
O
H
H
OH
H
H
OH
H
O
OH
O
H
OH
H
OH
H
H
OH
H
H
OH
CH2OH
CH2OH
H
O
H
OH
H
H
O
O
H
OH
H
H
OH
H
H
O
4
glycogen
H
1
O
6 CH2
5
H
OH
3
H
CH2OH
CH2OH
O
H
2
OH
H
H
1
O
O
H
4 OH
H
H
OH
H
H
O
O
H
OH
H
H
OH
H
OH
Il Glicogeno, è lo zucchero di deposito degli
animali, è simile all’amilopectina ma ha più
ramificazioni α(1Æ6).
La struttura fortemente ramificata consente il
rilascio rapido del glucoso dai depositi di glicogeno
(durante l’attività fisica): questa esigenza è sentita
più fortemente dagli animali che dalle piante .
CH2OH
H
O
H
OH
H
OH
H
1
O
H
H
OH
6CH OH
2
5
O
H
4 OH
3
H
H
H 1
2
OH
O
O
H
OH
CH2OH
CH2OH
CH2OH
H
O
H
O
H
OH
H
OH
O
H
O
H
OH
H
OH
OH
H
H
H
H
H
H
H
OH
cellulose
La Cellulosa, è il costituente principale delle pareti cellulari
delle piante: consiste di catene lineari di glucoso con legami
β(1Æ4).
Un glucoso ogni due è ribaltato, a causa del legame β.
Questo comporta legami idrogeno intra e inter-catena ed
interazioni di van der Waals. Queste fanno si che le catene di
cellulosa possano essere dritte e interagire lateralmente per
formare fasci. La cellulosa impartisce rigidità e resistenza alle
pareti cellulari. Gruppi di Celluloso Sintasi sulla membrana
plasmatica producono fasci di catene parallele di cellulosa
che patono dalla superficie della cellula vegetale.
La cellulosa
Carboidrati di deposito
Carboidrati strutturali
Gli oligosaccaridi delle glicoproteine e dei glicolipidi possono essere lineari
o ramificati. Spesso includono zuccheri modificati.
Gli oligosaccaridi delle glicoproteine possono avere semplicemente un
legame glicosidico da uno zucchero ad una Ser o Thr.
Legami all’azoto
Il legame può
avvenire anche con
un’Asn.
CH2OH
O
O
H
H
OH
HN
C
HN
CH2
C
H
H
OH
H
HN
CH
C
CH3
O
N-acetylglucosamine
Initial sugar in N-linked
glycoprotein oligosaccharide
Asn
O
HN
HC
R
C
O
X
HN
HC
C
R Ser or Thr
O
Lipidi
• Derivati degli idrocarburi
– Includono i grassi, gli oli, le cere, i
fosfolipidi, gli steroidi.
– Sono componenti strutturali delle
membrane, depositi energetici,
pigmenti, isolanti e segnali
intracellulari.
– Derivano dai carboidrati.
Non sono polimeri, in larga misura
Lipidi
Lipidi sono non-polari (idrofobici), solubili nei
solventi organici.
La maggior parte delle membrane lipidiche sono
anfipatiche, cioè hanno un terminale non polare
ed uno polare.
Gli acidi grassi, i lipidi più semplici, consistono
di una catena idrocarburica con un carbossile ad
una estremità. CH3(CH2)14-COOnon-polare
polare
Lipidi
6 funzioni:
1. deposito di energia a lungo termine
2. Isolamento termico
3. protezione dagli shock meccanici
4. protezione dalla disidratazione
5. messaggeri chimici (ormoni)
6. componenti principali delle membrane
(phospholipids)
O
C
β
Notazione abbreviata
γ
α
−
3
O
1
di un acido grasso
4
2
con 16 carboni, un
doppio legame cis
Acido grasso con un
tra C9 e C10 è 16:1
legame doppio cis-Δ9
cis Δ9.
Alcuni esempi:
14:0 acido miristico
16:0 acido palmitico
18:0 acido stearico
18:1 cisΔ9 acido oleico
18:2 cisΔ9,12 acido linolenico
20:4 cisΔ5,8,11,14 acido arachidonico
I doppi legami
O
β
γ
α C
degli acidi
−
3
O
1
4
2
grassi sono
solitamente
Acido grasso con un
nella
legame doppio cis-Δ9
configurazione
cis
La maggior parte degli acidi grassi naturali ha un
numero pari di carboni.
Il doppio legame produce una piegatura della catena
Quando i doppi legami sono in trans, la piegatura
non si forma.
I grassi
A temperatura ambiente le
molecole degli acidi grassi
saturi sono impacchettate
efficientemente e formano un
solido
Le molecole di un grasso
insaturo non possono
impacchettarsi
efficientemente, per cui
restano in forma liquida
Glicerofosfolipidi (fosfolipidi)
I glicerofosfolipidi sono i
comuni costituenti delle
membrane cellulari. Hanno lo
scheletro del glicerolo.
Gli ossidrili al C1 e C2 sono
esterificati con acidi grassi,
spesso diversi (per lunghezza o
doppi legami)
CH2OH
H
C
OH
CH2OH
glycerol
L’altro carbonio del glicerolo è
esterificato con un fosfato, a sua
volta spesso esterificato con l’OH di
una molecola polare, come la
serina, la colina, l’etanolammina,
l’inositolo, il glicerolo
polar
"kink" due to
double bond
non-polar
O
O
R1
C
H2C
O
O
R2
O
CH
H2C
C
O
P
O−
CH3
O
CH2
CH2
+
N CH3
CH3
phosphatidylcholine
La fosfatidilcolina, è un comune lipide di
membrana
O
O
R1
C
H2 C
O
O
CH
H2 C
C
R2
O
O
P
O
O−
OH
phosphatidylinositol
H
OH
H
OH
OH
H
H
H
H
OH
fosfatidilinositolo, è un lipide di membrana ed ha
un ruolo come molecola di segnale cellulare.
• Modello
molecolare
L’autoassemblaggio dei fosfolipidi per formare la membrana
cellulare
Lipid bilayer (da simulazione)
Gli steroidi
Colesterolo
HO
Cholesterol
Il Cholesterol ha un sistema policiclico rigido e
una catena breve e ramificata. È idrofobico con un
gruppo ossidrilico polare (anfipatico).
Si trova nelle membrane, è il precursore per la
sintesi degli ormoni steroidei e della vitamina D.
Le macromolecole interagiscono tra loro
Il ribosoma