Allenamento pliometrico: teoria e pratica parte 1
Introduzione
La pliometria è un metodo di allenamento teso allo sviluppo della forza, specie di quella esplosiva,
reattiva ed esplosivo-elastico-riflessa, viste come attitudini di lavoro del sistema neuromuscolare
durante un carico dinamico, inteso come la capacità di passare rapidamente da un regime di
contrazione eccentrico ad uno concentrico attraverso lo sfruttamento del ciclo stiramentoaccorciamento (CSA) o stretch-shortening-cycle (SSC), il quale crea i presupposti per stimolare la
tensione muscolare attraverso l'assorbimento di energia cinetica da parte del corpo e la restituzione
sotto forma di lavoro.
La capacità reattiva può essere espressa dalla formula: R=R2/R1
Dove R, è qualsiasi capacità di lavoro muscolare nelle fasi di stiramento e di contrazione attiva, R1
è la forza nella fase di spinta e R2 è il valore di forza nella fase di ammortizzazione, e dove la
capacità reattiva è tanto maggiore quanto più è alto tale rapporto.
Storia
Il padre di tale metodo è Yury Verkhoshansky, che notò, quando negli anni ‘50 era allenatore della
squadra universitaria "Burevesnik" di Mosca, che nel salto triplo e in modo più specifico nel
secondo balzo, lo "step", in una frazione di 0,125 secondi vi era un'applicazione di forza su un solo
arto pari a circa 300 Kg. Da qui l'idea di usare come sovraccarico non il comune bilanciere ma
l'energia cinetica della caduta e sfruttare il peso dell'atleta. Battezzò tale metodo dapprima "jump
training" e in seguito "metodo d'urto".
Propose inizialmente 3 tappe per introdurre tale metodologia sui campi d'atletica:
1. Periodo preparatorio, in cui si ricercava un incremento delle capacità generali ed un
adattamento al lavoro.
2. Periodo in cui si affiancavano esercizi che sfruttavano il CSA con esercizi classici con
sovraccarichi.
3. Periodo che sfruttava esercizi con CSA ad alta intensità.
Nel 1967 cominciò a sfruttare altezze di caduta comprese tra 0,75 e 1,15 m, da allora in poi furono
numerosi gli allenatori, autori, preparatori atletici che introdussero tale metodica in ogni tipologia di
sport, dando così forte impulso alle applicazioni e alla bibliografia in merito.
Ricordiamo Zatsiorsky, Margaria, Piron, Bosco, Zanon, Asmussen, Wilt.
Oggi si può notare ancora un piccolo fraintendimento riguardo all'uso dei termini "metodo d'urto" e
pliometria, dove, nel primo metodo vi è uno sfruttamento del regime stiramento accorciamento, nel
secondo invece della sola contrazione eccentrica, anche se oggi entrambe le nomenclature sono
usate come sinonimi.
Anche nello scrivere la parola pliometria vi è un dibattito in merito all'utilizzo della "i" o della "y" (
pliometria o plyometria, la "y" è tipica dell'espressione Anglosassone, la "i" di quella Europea).
Esiste inoltre ancora la distinzione tra esercizi definiti a "carattere pliometrico" ed "esercizi
pliometrici propriamente detti", dove alla prima categoria appartengo esercizi che sfruttano il CSA
non in modo massimale, alla seconda invece quelli in cui si fa ricorso all'utilizzazione massiva
dell'energia elastica, dispiegando in tutte le possibilità il sistema nervoso e muscolare attraverso
l'utilizzazione di tempi di contatto al suolo minimi.
Maglia connettivale
Il muscolo si compone di diversi elementi che formano a mo' di puzzle la sua compagine. Le
diverse unità funzionali sono i sarcomeri o inocommi, vere e proprie unità funzionali del
movimento. Non è argomento di questo articolo spiegare né le funzioni né i processi che sono alla
base del funzionamento del sarcomero.
Per capire però come avvenga la restituzione dell'energia elastica durante un CSA, è necessario
avere chiara la disposizione della maglia proteica che sottende alle funzioni del muscolo stesso,
cosa modula a livello del sistema nervoso tali processi e le relazioni fisiche che ne sono alla base.
Da un punto di vista semplicistico le proteine che compongono il sarcomero le possiamo dividere in
tre categorie:
1. Proteine contrattili: Actina e Miosina.
2. Proteine regolatrici: Troponina e Tropomiosina.
3. Proteine strutturali: Titina, Nebulina, Desmina ecc..
Se si osserva poi un preparato muscolare al microscopio si può facilmente notare la presenza di
zone di diverso colore, chiamate dagli studiosi bande o zone (fig. 1).
Ritroviamo così principalmente:
Dischi Z - Delimitano il sarcomero, sono i punti di ancoraggio per le proteine, attraversano
tutte le fibre e si attaccano al sarcolemma, sono sede di lesioni durante lavoro muscolare.
Banda A - corrisponde alla lunghezza del filamento di Miosina.
Banda I - corrisponde a due file di Actina in due sarcomeri contigui.
Banda H - corrisponde alla zona compresa tra due file di Actina nello stesso sarcomero.
Linea M - linea che divide il sarcomero in due porzioni simmetriche, si posiziona a metà
della banda scura.
Fig. 1 – Bande del tessuto muscolare.
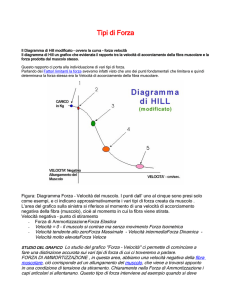
Schema di Hill
Hill negli anni '50 suppose che tale maglia connettivale avesse un ruolo, nella tensione del muscolo
a riposo, nella protezione contro le elongazioni eccessive, ed un ruolo nella restituzione dell' energia
elastica accumulata durante il lavoro muscolare.
L'autore divise il muscolo da un punto di vista "fasciale" in tre zone (fig. 2):
1. Componente contrattile (CC).
2. Componente elastica in serie (CES). Tendini e ponti acto-miosinici.
3. Componente elastica in parallelo (CEP). Guaine muscolari.
Fig. 2 – Schema di Hill
Per Hill (1950), Chapman (1985), Huijing (1992), il maggior responsabile della restituzione di
energia è il CES ( testa S2 della miosina e tendine), invece la generazione di forza dipenderebbe da
CC (ponti acto-miosinici) e CEP (sarcolemma), il CEP interviene solo per escursioni ampie, per cui
non è attivato nei movimenti naturali, ma dovrebbe avere solo funzione protettiva.
Elementi elastici non smorzati - Hill ipotizzò nel 1950 che gli elementi elastici fossero presenti
anche nella materia contrattile, li battezzò elementi elastici non smorzati. Huxley e Simmons (1971)
avvalorarono tale supposizione.
Tendini - Sono formazioni connettivali a fibre parallele, composti da polipeptidi avvolti in modo
elicoidale, che hanno la funzione di collegare principalmente il muscolo all'osso. La loro morfologia
consente che sotto carico aumenti la loro lunghezza e ne diminuisca il diametro.
Ponti acto-miosinici - Supponiamo una contrazione isometrica ideale, ovvero senza avvicinamento
dei dischi Z. Nonostante questa ipotesi la testa della Miosina agganciandosi comunque ruoterebbe e
allungherebbe la coda del ponte. Ciò farebbe dedurre la presenza di una componente elastica a tale
livello e il fatto che quest'ultima si possa ripercuotere alle estremità del muscolo.
L'accorciamento del muscolo da un valore di massima contrazione massimale fino a 0, è pari a circa
lo 0,4% per mezzo sarcomero, poiché la componente tendinea s'infiltra nella compagine del
muscolo a diversi livelli avremo diversi gradi di allungamento, dallo 0,4%, al 2% per il sartorio di
rana, al 4% per il gastrocnemio di rana ( Lavagna, Citterio,1974).
Da cui Chapman (1985) e Huijing (1992), hanno potuto affermare che nel riuso elastico il tendine
immagazzina il 72% e la testa S2 della Miosina il 28% dell'energia totale. Inoltre quando vi è un
accumulo di energia nel pacchetto tendine-muscolo, il valore dell'energia immagazzinata è
proporzionale alla forza esercitata e alla deformazione indotta. Detto questo, siccome muscolo e
tendine sono in serie, la forza applicata ad entrambi è uguale, ma data la diversa rigidità dei due
tessuti, costante nel tendine, variabile nel muscolo, dove più alta sarà la tensione maggiore sarà la
stiffness e tanto maggiore la sua resistenza all'allungamento. A partire da queste considerazioni si
evince che vi è un maggiore accumulo di energia a livello tendineo (Kreamer).
Riflesso da stiramento e riflesso del Golgi
Un alto numero di attività del nostro organismo avvengono senza controllo cosciente. Tutte queste
capacità di mantenimento omeostatico sono chiamate "attività riflesse". Tra queste il più semplice è
l'arco riflesso o monosinaptico, costituito da cinque elementi fondamentali:
1.
2.
3.
4.
5.
un recettore sensibile agli stimoli prodotti dai mutamenti esterni.
un neurone afferente che guida l'impulso dal recettore al Sistema Nervoso Centrale.
una sinapsi all'interno della quale spesso è posizionato un interneurone.
un neurone efferente che guida gli impulsi di risposta.
un neurone effettore che si attiva per mantenere l'omeostasi.
Gli organi dell'apparato propriocettivo sono due: Fusi Neuromuscolari e gli Organi Tendinei del
Golgi. Queste due strutture sono protagoniste del lavoro pliometrico, anche se in modo inverso in
quanto il riflesso miotatico tende ad aumentare la risposta, invece il riflesso del Golgi tende a far
diminuire la prestazione. Vediamoli più nel dettaglio.
Fusi neuromuscolari
Recettori così chiamati per la loro morfologia, sono formati da una guaina, che contiene da 8 a 12
(in media) fibre muscolari (intrafusali), sparse e poste parallele a quelle extrafusali (fig.3).
Abbiamo due tipi di fibre intrafusali:
1. a sacco nucleare
2. a catena nucleare.
Dove le prime, hanno i nuclei raggruppati nella parte centrale, le seconde lungo l'asse longitudinale.
Entrambi i tipi di fibre hanno la zona centrale non contrattile e quella periferica contrattile e striata.
Dalla parte centrale origina una terminazione anulospirale che si avvolge attorno alla fibra, da cui
originano le fibre afferenti Ia per stiramenti veloci e brevi, ma anche per quelli tonici. Tra centro e
periferia originano le terminazioni secondarie dalle quali dipartono le fibre afferenti di tipo II, per
gli stiramenti tonici. Tale tipo di fibre intrafusali prendono il nome di gamma, in quanto innervate
da gamma motoneuroni. Tali formazioni servono ad informare sulle condizioni dello stiramento
muscolare, inviando segnali agli alfa motoneuroni del muscolo agonista, e attraverso neuroni della
zona grigia intermedia del midollo spinale, agli alfa motoneuroni del muscolo antagonista, che
viene inibito.
Fig. 3 – Fusi neuromuscolari.
Organi tendinei del Golgi
Recettori posti in serie alla fibra, nella giunzione mio-tendinea. Sono arborizzazioni nervose che
avvolgono fasci di fibre di tessuto connettivo-collageno del tendine. Hanno forma allungata, di 1
mm e diametro di 0,1 mm, ed hanno una capsula che li contiene. Sono innervati da un singolo
assone (fibra di tipo Ib), che perde la mielina all'interno della capsula, dividendosi in tante
diramazioni interposte tra le fibre. Dagli OTG partono fibre afferenti di tipo Ib che vanno nel
midollo agli alfa motoneuroni (fig. 4).
Il tempo che intercorre tra stiramento iniziale e l'aumento di forza è compreso tra 50 e 55 ms, dove
40-45 sono dovuti al riflesso ed il restante 12-14% a ritardo elettromeccanico. Per Schimidtbleicher
(1986) ci sono due forme di impulso che usano lo SSC:
1. SSC lento- con marcato spostamento articolare ed attività elettrica di durata di 300-500ms.
2. SSC rapido- con movimenti angolari ridotti e tempi di contatto di 100-200 ms.
Molti movimenti sportivi sono assimilabili ad entrambi i tipi di SSC.
La fase di preattivazione ha un ruolo fondamentale durante tutti i salti con SSC rapido. Essa
dipende dall'intensità dello sforzo, è regolata dal controllo visivo ed aumenta la rigidità muscolare
prima del contatto al suolo. Altra peculiarità della preattivazione è quella di aumentare la sensibilità
dei fusi neuromuscolari. Circa 20-30 ms per gli arti superiori e 30-45 ms per gli arti inferiori dopo il
contatto al suolo appare un picco sul tracciato EMG, che corrisponde all'intervento del riflesso da
stiramento.
Fig. 4 – Organi tendinei del Golgi.
Circuito di Renshaw
Vedendo il soma di un motoneurone ed il suo assone, noteremo che quest'ultimo emette dei
ramuscoli collaterali che fanno sinapsi con delle cellule presenti nel corno anteriore della sostanza
grigia, chiamate cellule di Renshaw (fig. 5), che a loro volta inviano un'innervazione ricorrente al
soma del motoneurone stesso, che serve a dare informazioni inibitorie nel caso vi sia un'attività
efferente in eccesso del motoneurone. Tale circuito è denominato di Renshaw.
Fig. 5 – Cellula di Renshaw.
Quindi il circuito gamma è un apparato di rilevazione e modulazione dell'attività muscolare.
I motoneuroni alfa del muscolo agonista, sono controllati da funzioni inibitorie che contribuiscono a
riportare il sistema in condizioni di equilibrio, grazie alle afferenze dei fusi del muscolo antagonista,
dagli OTG e del circuito ricorrente di Renshaw.
Relazioni fisiche
Le relazioni fisiche che legano le condizioni muscolari nel lavoro pliometrico sono diverse:
Relazione forza lunghezza
Il picco di forza dipende dal grado di accavallamento delle proteine contrattili. Una fibra a riposo ha
più o meno lunghezza di 2,5 micrometri, con possibilità per il sarcomero di raggiungere lunghezze
che si piazzano intorno ai 3,65 micrometri, in quanto i filamenti spessi sono 1,6 e quelli sottili 1. Il
picco di forza si ha quando il sarcomero è in una lunghezza compresa tra 2 e 2,2 micrometri.
Relazione forza velocità
Negli anni '40 il fisiologo Hill trovò la relazione che legava la forza e la velocità. Dal grafico
rappresentante tale relazione si può evincere che la velocità è massima a carico zero e la forza è
massima a velocità zero. Il miglior compromesso che lega tali due parametri si situa al 30-40%
dell'1 RM. Tale curva ha un carattere iperbolico e non può essere modificata con l'allenamento (fig.
6).
Fig. 6 – Curva di Hill.
Relazione velocità lunghezza
La forza muscolare è proporzionale al diametro trasverso della fibra, la velocità invece dipende dal
numero di fibre in serie lungo il decorso della fibra. Per cui se ipotizzassimo un accorciamento
Delta L e avessimo 1000 sarcomeri in serie, la velocità generata dall'accorciamento totale sarebbe
1000xDelta L/Delta t. Più lunghi sono i muscoli e più hanno a disposizione traiettorie di
accelerazione. La forza nella fase eccentrica aumenta sensibilmente con l'aumentare della velocità
nelle fasi iniziali per poi stabilizzarsi (negli atleti esperti).
Relazione velocità ipertrofia
La forza è proporzionale al diametro trasverso del muscolo, l'eccessiva ipertrofia aumenta le
viscosità interne e la retrazione connettivale da qui non favorisce i movimenti di tipo esplosivo-
balistici. Da tale relazione è anche deducibile la maggiore forza eccentrica dei Bodybuilders in
quanto l'esasperata ipertrofia crea attriti più alti nei movimenti cedenti.
Fisiologia della pliometria
Il riuso elastico è presente in tutti i gesti, dalla semplice locomozione, alla corsa, al sollevamento
pesi.
L'ammontare della restituzione dell'energia elastica nella fase concentrica dipende da:
1.
2.
3.
4.
Stiramento esclusivamente attivo del muscolo.
Ampiezza dello stiramento.
Velocità di stiramento.
Coupling time.
Stiramento esclusivamente attivo del muscolo. L'immagazzinamento dell'energia elastica può
avvenire solo in tal caso, perché in caso di attivazione l'allungamento è a carico della CC e della
CEP, mentre la CES essendo più rigida non subisce variazioni di lunghezza.
Ampiezza dello stiramento. Quando il muscolo è sottoposto a un carico pari alla sua massima
forza isometrica, si è visto che l'ampiezza dell'accorciamento aumenta con l'ampiezza dello
stiramento.
Quando tale effetto è massimo si possono distinguere quattro fasi dell'accorciamento:
1° fase - accorciamento elastico simultaneo con la caduta di forza.
2° fase - rapido accorciamento isotonico.
3° fase - inflessione del tracciato lunghezza tempo, dovuto alla riduzione e all'inversione della
velocità.
4° fase - accorciamento isotonico.
Per Iles (1977), Gottliebeg e Agarwall (1979), il riflesso da stiramento si attua entro 40-50 ms
dall'inizio della fase di stiramento, da cui Bosco (1977), indicò che se la fase eccentrica fosse stata
troppo lunga, l'azione del riflesso miotatico sarebbe ricaduta all'interno della fase eccentrica stessa.
Influenza della velocità di stiramento. La contrazione eccentrica deve essere immediata altrimenti
si assisterà ad una caduta di forza prima rapida e poi lenta. Per Abbott e Aubert (1952), la velocità
di caduta della forza è tanto maggiore quanto più veloce è stato lo stiramento.
Coupling time. Sulla base di ciò bisogna dedurre che l'atleta debba possedere un buon bagaglio
motorio e coordinativo per cercare di restare in equilibrio e quindi di poter sfruttare l'energia
elastica accumulata, specie durante l'inversione del movimento (coupling time). Quindi la fase di
inversione del movimento deve essere inferiore al tempo medio di vita di un ponte actomiosinico,
ossia 120-150 ms nella FTb.
Per Komi e kollhorer (1997) lo stretch shortening cycle (SSC) o ciclo stiramento accorciamento,
richiede tre condizioni fondamentali:
1. Una pre-attivazione prima della fase eccentrica.
2. Una fase eccentrica corta e rapida.
3. Passaggio eccentrico-concentrico immediato.
Questo conduce ad una espressione della forza superiore alla massima isometrica del 150-200%
fino al 500-600%, riconducibile a sua volta ad altri tre fattori quali:
1. intervento di fattori neuromuscolari:
o reclutamento spaziale delle unità motorie.
o frequenza di scarica massima.
o maggiore sincronizzazione delle unità motorie.
2. Elasticità tendine muscolo.
3. Riflesso da stiramento.
Da ciò si è dimostrato che lo sfruttamento del CSA aumenta il rendimento del gesto esclusivamente
muscolare, ovvero supportato da esclusiva fonte biochimica grazie alla restituzione dell'energia
elastica accumulata nel sistema tendine muscolo.
Per Paillard le unità motorie si sincronizzano per due motivi:
1. Perché sottomesse allo stesso ordine centrale.
2. Per effetto "ephatique", ovvero per gli effetti del campo elettrico, per vicinanza di tali
motoneuroni nel nucleo motore.
Per Zatsiorsky la sincronizzazione delle unità motorie avviene in determinate condizioni, quali:
Lavoro esplosivo.
Alti carichi.
Isometria lunga.
Lavori misti.
Piron ha proposto tre condizioni per l'allenamento pliometrico sulle quali si può agire:
1. posizione (piazzamento) - ovvero ricercare quella specifica della disciplina.
2. Spostamento sull'appoggio - senza variare l'angolo al ginocchio.
3. Variazione della tensione - aumentando o diminuendo l'altezza di caduta oppure utilizzando
altre modalità di azione muscolare.
Le sedute di lavoro pliometrico possono essere principalmente di cinque tipi:
1.
2.
3.
4.
5.
con balzi orizzontali.
con balzi verticali.
con sedute miste.
sedute di balzi ripetuti, in cui si associano diversi tipi di balzi.
metodo Bosco Pittera. Metodo introdotto dai due studiosi nel 1982, su giocatori nazionali di
calcio e pallavolo, consiste nello scendere giù dal plinto con un angolo al ginocchio di 90°,
per poi atterrare in un range articolare compreso tra i 90° e i 110°. Tale metodo ha come
caratteristica principale di avere un momento sul braccio di leva del ginocchio inferiore del
30% rispetto alla pliometria normale, il tutto con una tensione muscolare più bassa. La
particolarità di tale metodo è quello di lavorare ad angoli più usuali per il calcio. Tutto
questo confermato dagli studi di Bobbert, che vide inoltre che vi è era minore sollecitazione
di ginocchia e caviglie, con una potenza meccanica espressa nell'esercizio maggiore,
evidenziabile grazie al tracciato EMG. Bosco e Pittera provarono ciò da un'altezza di caduta
di 40-50 cm a due gambe e 20-30 cm ad una gamba.