Lezione 7
Il metabolismo
terminale e la
produzione di ATP
1
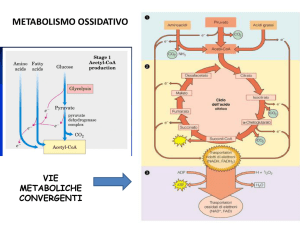
Il metabolismo terminale
Catabolismo Lipidi – Amminoacidi – Carboidrati
convergono sul metabolismo terminale,
e producono una stessa molecola: l’acetil-CoA
- ciclo di Krebs (o ciclo degli acidi tricarbossilici o
ciclo dell’acido citrico)
- fosforilazione ossidativa.
Entrambe le vie sono localizzate nei mitocondri
2
Relazione tra metabolismo
intermedio e terminale
3
Respirazione
Cellulare
I mitocondri
Coenzima A
Il coenzima A è sintetizzato a partire dalla vitamina
B5 (o acido pantotenico) e dall’aminoacido cisteina,
spendendo 4 ATP. La vitamina B5 è presente in molti
alimenti, specialmente uova, carne e latte. Il CoA è il
cofattore essenziale per la sintesi dell’acetil CoA.
7
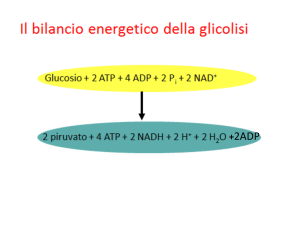
La piruvato deidrogenasi
3 enzimi
5 coenzimi
Trasforma il Piruvato in acetil-CoA , tramite DECARBOSSILAZIONE OSSIDATIVA del Piruvato.
Genera acetil Co-A che entra nel ciclo di Krebs
L’energia contenuta nel legame tioestere (che coinvolge uno zolfo) fra l’acetile e il
coenzima A viene usata per il legame con l’ossalacetato, uno degli intermedi del ciclo di
Krebs.
E’ una reazione IRREVERSIBILE (Importante dogma metabolico): i carboidrati possono
essere trasformati in lipidi , ma NON può avvenire il contrario.
In periodi di digiuno prolungato solo gli amminoacidi ( e non i grassi) possono essere
convertiti a glucosio
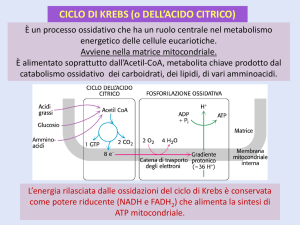
Il ciclo di Krebs (I)
Il ciclo di Krebs venne scoperto nel
1937 da H.A. Krebs, che lo
chiamò ciclo dell’acido citrico.
Successivamente, venne indicato
come ciclo degli acidi
tricarbossilici, dai tre intermedi
della fase di recupero dell’energia
(citrato, cis-aconitato e
isocitrato).
Hans Adolf Krebs ricevette il
premio nobel per la medicina
nel 1953
© Zanichelli editore, 2014
9
Il ciclo di Krebs (II)
A ogni giro del ciclo, un gruppo acetile (2C) entra
come acetil-CoA (punto di incrocio metabolico
fondamentale) e si condensa con ossalacetato (4C)
per formare il primo acido tricarbossilico (citrato).
Alla fine del ciclo il gruppo acetile è ossidato a 2 CO2 e
viene rigenerato ossalacetato. Quindi, teoricamente,
una sola molecola di ossalacetato basterebbe per
ossidare un numero illimitato di acetil-CoA.
Questo in realtà non succede perché l’ossalacetato e
altri intermedi vengono utilizzati anche come
precursori di molte biosintesi.
10
La struttura del ciclo di Krebs
Nel ciclo di Krebs possiamo osservare:
• La via degli elettroni, ovvero la generazione di potere
riducente attraverso la generazione di NADH e FADH2.
• La via dei carboni, ovvero il destino del gruppo acetile
attraverso le successive decarbossilazioni ossidative.
• La via del Coenzima A, in cui una molecola di CoA viene
liberata all’inizio dall’acetil-CoA, per essere poi utilizzata e
nuovamente rigenerata nella sequenza del ciclo:
a-chetoglutarato-> succinil-CoA -> succinato
11
Acetil-CoA+3NAD+ +FAD +GDP+Pi+2H2O = 2 CO2+ 3NADH + 3H+ +FADH2 +GTP+ CoA
Perché avvenga è necessario ossigeno
perché solo così la catena respiratoria
è attiva e può rifornire il ciclo di
coenzimi ossidati continuamente
necessari
Ciclo di Krebs
Le sequenze di reazioni del ciclo
di Krebs
Possiamo dividere idealmente il ciclo di Krebs in due blocchi
di reazioni:
1. La serie da citrato a succinato serve a operare le due
decarbossilazioni ossidative successive che rimuovono
i 2C entrati nel ciclo all’inizio come acetil CoA.
2. La serie da succinato a ossalacetato serve a
ripristinare il composto di partenza (ossalacetato) per un
nuovo giro.
3. In entrambe le serie di reazioni si ottiene energia sotto
forma di 4 coppie di elettroni e una molecola di GTP. Ma
solo nella prima serie viene prodotta CO2.
13
Ciclo di Krebs
14
Ciclo di Krebs
BILANCIO COMPLESSIVO
Acetil CoA + 3 NAD++FAD+GDP + Pi+2H2O= 2CO2+3 NADH +3 H+ +FADH2 + GTP
Ossidazione completa dei due carboni dell’Acetil CoA a anidride carbonica
Riduzione di 3 NAD e 1 FAD (per ciascuna molecola di AcetilCoA)
CONTROLLO
La velocità del ciclo è regolata dalla necessità di ATP da parte della cellula
Il principale ENZIMA REGOLATORE è la citrato sintasi che catalizza la reazione sotto
riportata. Il Citrato prodotto inibisce in maniera competitiva l’enzima.
1. Formazione del citrato
Acetil CoA reagisce con
l’ossalacetato (acido 2-oxo1,4-butandioico) per
formare il citrato (acido 3carbossi-3-idrossi-1,5pentandioico)
.
16
La decarbossilazione ossidativa
L’ossidazione dell’acetil-CoA, che forma 2 molecole di
CO2 (con liberazione di una coppia di elettroni per
ciascuna molecola) avviene grazie a due
deidrogenasi che agiscono su
-isocitrato > a-chetoglutarato (reazione 3)
- a-chetoglutarato > succinil- CoA (reazione 4).
17
La decarbossilazione ossidativa
18
Ripristino Ossalacetato
Per formare il Succinato, si rompe il
legame alta energia del Succinil CoA,
fosforilando la GTP che poi forma ATP.
Succinato > Fumarato > Malato >
Ossalacetato.
Formazione di FADH2 e NADH + H+
19
Il ciclo di Krebs è una via
anfibolica (I)
Il ciclo di Krebs fornisce numerosi precursori per le
vie biosintetiche:
- ossalacetato e a-chetoglutarato per la sintesi di
amminoacidi,
- citrato come fonte di acetil-CoA per la biosintesi di
acidi grassi,
- succinil-CoA per la sintesi dell’eme,
20
Il ciclo di Krebs è una via anfibolica (II)
21
Reazioni anaplerotiche (I)
Continuamente vengono sottratti intermedi dal ciclo di
Krebs per la biosintesi. Questo potrebbe rallentare la
respirazione se non venissero rimpiazzati.
Le reazioni dedicate a fornire sempre sufficienti
intermedi al ciclo sono dette anaplerotiche (di
riempimento).
La principale è la carbossilazione del piruvato ad
ossalacetato ad opera della piruvato carbossilasi
mitocondriale:
piruvato+CO2+ATP -> ossalacetato + ADP+Pi
22
Destino del piruvato in condizioni
aerobiche: ossalacetato
- importante intermedio del ciclo di Krebs,
- substrato per la gluconeogenesi: forma il fosfoenolpiruvato.
23
Il ciclo di Krebs genera elettroni
Nel ciclo di Krebs non viene prodotto direttamente
ATP. Un ATP è prodotto indirettamente a partire dal
GTP generato nella reazione 5.
Il ciclo invece genera 4 coppie di elettroni sotto
forma 3 NADH e 1 FADH2. Questi potranno essere
spesi per la sintesi di molte molecole di ATP.
il ciclo di Krebs è una specie di motore che usa
gruppi acetili come combustibile per trasformarli in
un flusso di elettroni.
24
La catena di trasporto degli elettroni (I)
Le coppie di elettroni conservate nel NADH e FADH2 devono
essere rese disponibili per la produzione di ATP.
Questo rigenererà le forme ossidate (NAD+ e FAD) che
potranno ricevere nuovi elettroni, mantenendo in moto il ciclo.
Gli elettroni sono ceduti da NADH e FADH2 a un sistema di
proteine presenti nella membrana interna del mitocondrio,
esposte verso la matrice, dove si svolge il ciclo di Krebs e
quindi facilmente accessibili.
Queste proteine costituiscono la catena di trasporto degli
elettroni o catena respiratoria.
25
La catena di trasporto degli elettroni (II)
26
Flusso degli elettroni nella catena
(I)
La catena respiratoria comprende quattro complessi
proteici:
• Complesso I: NADH deidrogenasi
• Complesso II: succinato deidrogenasi
• Complesso III: Q-citocromo c ossidoreduttasi
• Complesso IV: citocromo c ossidasi
27
Flusso degli elettroni nella catena
(II)
Questi complessi contengono proteine specializzate
nell’accettare e donare elettroni (flavoproteine,
proteine Fe-S, citocromi).
La capacità di trasferire elettroni di queste proteine
dipende dalla presenza di particolari coenzimi
(ubichinone) o di gruppi prostetici, ovvero strutture
molecolari in grado di legare elettroni, quali il gruppo
eme (nei citocromi) i gruppi ferro-zolfo (nelle
corrispondenti proteine), il gruppo
flavinmononucleotide (nella NADH deidrogenasi).
28
La catena di trasporto degli
elettroni (III)
Ossido-riduzioni
La catena respiratoria è una sequenza di ossido-riduzioni, in
cui le proteine dei vari gruppi (nella sequenza I-IV) accettano gli
elettroni (riducendosi) e li cedono immediatamente al gruppo
successivo (ossidandosi).
30
Ossigeno: accettore finale di eÈ necessario che gli elettroni alla fine vengano
eliminati: è questa la funzione dell’ossigeno che
funziona da accettore finale.
Una molecola di O2 accetta 4 elettroni dal complesso
IV, formando 2 molecole di acqua.
Quindi gli elettroni generati in ogni ciclo di Krebs (8)
necessitano di 2 molecole di ossigeno per essere
smaltiti.
31
Funzionamento della catena di trasporto
La catena respiratoria non genera direttamente ATP, bensì
energia, sotto forma di forza proton-motrice, che servirà per
32
la fosforilazione ossidativa.
Il gradiente elettrochimico
protonico (I)
Il trasferimento degli atomi di idrogeno attraverso la
catena respiratoria comporta la separazione tra ogni
atomo di H e il suo elettrone.
Gli ioni H+ così generati vengono pompati verso lo
spazio intermembrana.
L’ossidazione del NADH + H+ libera 2 elettroni e 2
protoni (H+). Gli elettroni entrano nella catena nel
complesso I e vengono trasportati dai complessi I-IV
all’ ossigeno.
I protoni vengono pompati attraverso la membrana
interna nello spazio intermembrana.
33
Il gradiente elettrochimico
protonico (II)
La presenza di una concentrazione elevata di H+ da un
solo lato della membrana farà sì che, nel momento in
cui lo spazio intermembrana (elevata concentrazione
di H+) venga messo in collegamento con la matrice
(bassa concentrazione di H+) si crei un gradiente
elettrochimico, in cui i protoni fluiscono dal
compartimento ad alta concentrazione verso quello a
bassa.
Il collegamento è creato da un complesso
transmembrana: l’ATP sintasi.
34
Il flusso elettronico attraverso la membrana interna dei mitocondri è
accompagnato da un pompaggio di protoni dalla matrice allo spazio
intermembrana.
Ciò genera una differenza di potenziale elettrico e una differenza
di pH tra i due lati della membrana rendendo quello che guarda la
matrice più alcalino.
Questa concentrazione protonica e questa separazione di cariche
determinano una energia elettrochimica che prende il nome di forza
motrice protonica che rappresenta la forma di conservazione dell'
energia prodotta e liberata nelle ossidazioni biologiche.
La membrana mitocondriale è impermeabile ai protoni che non
possono così rientrare per annullare il gradiente elettrochimico.
Ma quando questi potranno fluire attraverso i canali protonici specifici
posti nel complesso Fo dell'ATPsintetasi in direzione del gradiente
elettrochimico, si renderà disponibile una grande quantità di energia.
Il complesso F1 dell'ATPsintetasi condenserà un fosfato all'ADP
per generare ATP.
Il gradiente di H+ è la forza
che permette la
fosforilazione di ADP che
procede con la rimozione di
H2O.
ADP+Pi = ATP + H20
ADP + Pi = ATP + H+ (interno) +
OH- (esterno)
Fosforilazione ossidativa: ATP
sintasi
La teoria chemiosmotica (1962, P.Mitchell) spiega la
sintesi di ATP a partire dalla forza proton-motrice del
gradiente elettrochimico.
Il flusso di protoni che ritorna verso la matrice, infatti,
passa attraverso un canale costituito dalle proteine
della subunità F0 del complesso della ATP sintasi.
Questo canale, in realtà è un rotore, le cui componenti
vengono mosse dal flusso di H+.
38
Meccanismo della ATP sintasi
Questo movimento si
trasmette alle proteine della
testa (subunità F1) che
sporge verso la matrice e
dove ha sede la sintesi di
ATP a partire da ADP e Pi,
generando acqua. Gli
elettroni ceduti da una
molecola di NADH + H+
portano alla sintesi di 2
ATP, mentre quelli di un
FADH2 a 1,5 ATP.
39
Considerazioni finali sulla
produzione di ATP
La produzione di ATP (la moneta energetica della
cellula) quindi procede attraverso complesse
sequenze di reazioni connesse tra di loro:
1. Il catabolismo della macromolecole è connesso al
ciclo di Krebs grazie all’ acetil-CoA.
2. Il ciclo di Krebs è connesso alla catena
respiratoria grazie ai trasportati di elettroni ridotti
FADH2 e NADH+H+.
3. La catena respiratoria è connessa all’ATP sintasi
grazie alla forza proton motrice del gradiente
elettrochimico.
© Zanichelli editore, 2014
40
Resa energetica: 65% di 679 kcal
Emoglobina, O2, CO2 e
respirazione (I)
È possibile ora capire il nesso tra la respirazione
intesa come processo organo-meccanico di
inspirazione ed espirazione e la respirazione come
processo biochimico che porta alla sintesi di ATP.
Inspirando immettiamo l’ossigeno che viene captato
dall’emoglobina degli eritrociti e veicolato alle cellule,
dove funge da accettore finale degli elettroni della
fosforilazione ossidativa.
42
Emoglobina, O2, CO2 e
respirazione (II)
La quantità notevole di CO2 generata dalla
decarbossilazione ossidativa nel ciclo di Krebs e nelle
varie reazioni cellulari, viene ceduta all’emoglobina,
riportata ai polmoni ed emessa con l’espirazione.
La respirazione è quindi un bell’esempio di come
nell’organismo, ogni aspetto macroscopico sia
legato a precisi eventi molecolari.
43