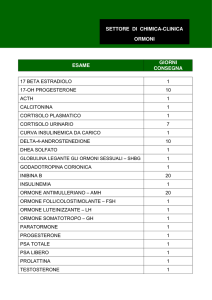
Endocrinologia, Prof. Pontecorvi, 18/10/06, 8.00-10.00
Prima ora
Il prof. Pontecorvi ci farà le lezioni assieme all’endocrinochirurgo, prof. Lombardi.
La parte pratica, per cui siamo stati divisi in gruppi, dovrebbe aver luogo dopo la fine delle lezioni
per protrarsi fino al 9 febbraio. Il tutor di ciascun gruppo dovrebbe mostrarci e guidarci nelle attività
scritte sul vangelino. L’accordo è quello di fare una settimana full immersion: tutte le mattine.
Uscirà una lista dei gruppi con il ricercatore affiancato che sarà il nostro tutor, e ci mettiamo
d’accordo per fare queste 21 ore. 3 ore al giorno per una settimana dovrebbe andar bene, oppure
dividiamo in modo alternativo. Può addirittura iniziare da subito.
Ripeteremo di ciascuna ghiandola la parte anatomica e fisiopatologica per arrivare poi alla parte
clinica e ai dati di laboratorio, fondamentali in endocrinologia per fare la diagnosi.
Le vere chiandole endocrine sono queste cinque:
- Al di sopra di tutto, l’ipotalamo, in realtà parte del sistema nervoso centrale che secerne
diversi fattori che regolano la secrezione ipofisaria. È la connessione fra il sistema nervoso e
quello endocrino. Tant’è vero che si parla di neuroendocrinologia per indicare questo campo
di studio.
- La “regina”, l’ipofisi, con i suoi ormoni che controllano il funzionamento di diverse altre
- La tiroide, che produce gli ormoni tiroidei e la calcitonina, e dietro al tiroide le quattro
(solitamente) paratiroidi, che regolano il metabolismo del calcio e del fosforo.
- I surreni, che producono i mineralocorticoidi e i glucocorticoidi nella corticale, e le
catecolamine nella midollare.
- Nella donna, le ovaie.
- Nell’uomo, il testicolo.
Questo è in realtà un modo vecchio di vedere le ghiandole endocrine, perché nel tempo sono stati
scoperti tanti nuovi ormoni e tante nuove cellule che li producono. Si è visto che ghiandole
endocrine possono essere sparse in diversi punti del nostro organismo, e in particolare
- lungo tutto il decorso della parete del sistema gastrointestinale.
- All’interno del pancreas, a livello delle insulae del Langerhans
Che secernono insulina, glucagone, gastrina e tutta una serie di peptidi anche di recentissima
identificazione che hanno ruolo soprattutto nel regolare l’apparato gastroenterico stesso, l’appetito,
la termoregolazione e così via.
Sempre cellule sfuse sono presenti altrove: per esempio
- a livello degli alveoli polmonari, cellule di origine neuroendocrina (cioè derivate dalle creste
neurali)
- a livello del cuore, dove è prodotto il peptide natriouretico atriale.
Così, oggi quando si parla di sistema endocrino si parla di qualcosa di diffuso, presente a livello di
un po’ tutti gli organi.
Queste cellule possono essere
- prodotte da neuroni (neurormoni)
- prodotte da cellule ghiandolari specializzate
- prodotte da cellule neuroendocrine
Interrelazioni tra sistema endocrino, SNC e sistema immunitario: il sistema
enuroimmunoendocrino. Il prof proietta uno schema che semplicemente mostra come ciascuno dei
tre sia in contatto con gli altri due, sottolineando però che il sistema immune comunica con
l’endocrino tramite sia immunomodulatori che autoimmunità
Prendiamo la reazione di fuga e allarme. Il sistema nervoso periferico, tramite la vista e l’olfatto,
stimola il sistema nervoso centrale a produrre a livello ipotalamico fattori che stimolano l’ipofisi a
secernere i fattori che aumentano la secrezione di glucocorticoidi e catecolamine.
Altro esempio: lo stress che altera il ciclo mestruale femminile, in questo caso dovuto ad anormale
ciclicità delle gondotropine ipofisarie.
Molti peptidi (citochine) che sono prodotti dalle cells del sistema immunitario (macrofagi, linfociti
T, linfociti B) hanno azione simile a quella degli ormoni. Può essere, come per gli ormoni, ad
azione paracrina o endocrina.
Anzi, alcune citochine, in alcune situazioni, svolgono una funzione assolutamente ormonale. Per
esempio, il rimodellamento osseo è ottenuto proprio tramite la liberazione in loco di citochine cme
l’IL2 e l’IL6, che qui svolgono un ruolo chiaramente diverso da quello nel sistema immunitario.
Molte malattie autoimmuni sono tipiche del sistema endocrino. La tiroidite di Hashimoto e il morbo
di Graves-Basedow sono modelli di malattie autoimmuni.
Dunque, il sistema nervoso centrale e quello immune dialogano con l’esterno, ciascuno facendo il
suo mestiere: il primo tramite i cinque sensi, il secondo reagendo ad agenti patogeni (salvo
fenomeni autoimmuni). Ed entrambi a loro volta dialogano, sul versante interno, con l’endocrino.
A. Funzioni del sistema endocrino regolare mediamnte l’azione di numerosi ormoni, il
differenziamento, la crescita e la funzione biologica dei tessuti e delle cellule dell’organismo
B. Ghiandole endocrine. Organi o tessuti formati da cellule endocrine ch eimmettono
direttamente nel ircolo periferico uno o più messaggeri chimici (ormoni)
C. Ormoni: messaggeri chimici prodotti dalle cellule endocrine che esercitano la propria
azione su di una cellula bersaglio a distanza, regolandone le funzioni biologiche
D. Cellule bersaglio: cellule contenenti specifici recettori capaci di interagire con l’ormone
mediandone l’effetto biologico ed inducendo una specifica risposta cellulare.
E. Recettori: molecola biologica, in genere proteina, che può essere situata sulla membrana
plasmatica, nel nucleo o, più raramente, nel citoplasma, che interagisce con l’ormone in
maniera esclusiva (il classico meccanismo chiave-serratura o in modo più complesso).
La patologia può essere dovuta
- alle variazioni quantitative dell’ormone, che possiamo misurare
- all’alterata interazione con i diversi tipi di recettori, che non possiamo misurare
Dalla rappresentazione quantitativa e qualitativa dei recettori che l’ormone trova a valle dipende
l’effetto endocrino!
F. Neurormoni: ormone prodotto da una cellula nervosa: es. ADH
a. Neurormone peptidico: es. “Releasing Hormones” ipotalamici
b. Neurormone non peptidico: es. Adrena,ina, Noradrenalina
c. Neurotrasmettitore: messaggero chimico che agisce a livello sinaptico
d. Neuromodulatore: ormone modulante la risposta di un neurone all’azione di un
altro ormone o neurotrasmettitore.
Modalità di rilascio ed azione ormonale
Via endocrina: l’ormone rilasciato dalla ghiandola o cellula ghiandolare è trasportato per via
ematica.
Via neuroendocrina: l’ormone rilasciato dal neurone è trasportato per via ematica.
Via neurocrina: il neurone contatta la cellula bersaglio tramite il suo assone ed il messaggero
chimico è rilasciato nello spazio sinaptico.
Via paracrina: l’ormone agisce sulle cellule vicine attraverso lo spazio extracellulare (es. tireociti
stessi in reazione agli ormoni tiroidei prodotti da un tireocita vicino!)
Via autocrina: l’ormone agisce sulla stessa cellula di origine
Proprietà generali degli ormoni
- presenti in circolo a concentrazioni molto basse (pM o nM), tanto che hanno potuto essere
misurati per la prima volta solo all’inizio degli anni ’60, quando furono inventati i metodi
immunologici. Con il tempo, poi, i sistemi di misurazione si sono raffinati attraverso l’uso di
anticorpi monoclonali, di traccianti non radioattivi ma fluorescenti e immunoenzimatici.
- Breve emivita: da pochi minuti (PTH) ad alcuni giorni (tiroxina)
- Funzioni generali di controllo su:
o Crescita e differenziamento cellulare
o Sviluppo sessuale e riproduzione
o Omeostasi metabolica ed energetica
o Contrattilità della muscolatura liscia
o Comportamento animale (neuropsicoendocrinologia)
Origine e classificazione degli ormoni
Dalle proteine:
- Proteine (es. GH, prolattina)
- Glicosilazione (TSH, LH)
- clivaggio peptidi (ACTH)
- clivaggio aminoacidi (Tiroxina o T4)
Dagli aminoacidi (nel senso che non vanno a dare veri polipeptidi)
- modificazione (es. adrenalina)
- piccoli peptidi (es. TRH)
Colesterolo
- steroidi (es. cortisolo, vitamina D)
Acidi grassi
- retinoidi (es. acido trans-retinoico)
- eicosanoidi (es. prostaglandina E1)
Classificazione degli ormoni: struttura e localizzazione recettoriale
1. Peptidi e glicoproteine
-
derivazione: aminoacidi
-
-
localizzazione recettoriale: membrana cellulare (perché non possono attraversarla, per
ragioni sia di massa che di carica), con la struttura recettoriale tipicamente con elemento
esterno (che riconosce l’ormone) e uno interno (che trasmette all’interno il messaggio)
origine: tutte le ghiandole endocrine escluse surrene e tiroide
esempi: insulina (51 aa), TSH, LH, ormone crescita, prolattina, ecc.
2. Steroidi
- derivazione: colesterolo
- localizzazione recettoriale: citoplasma e nucleo (perché possono attraversare la membrana,
essendo lipofili e non troppo grandi), anche se ci sono eccezioni come il progesterone, che
ha anche dei recettori di membrana; l’effetto ultimo è quello di regolare l’espressione genica
del DNA, il che può essere fatto da
o una catena di signaling intracellulare a partire dalla membrana e attraverso l’uso di
secondi messaggeri
o direttamente dal complesso recettore-ormone, come accade più spesso per gli ormoni
steroidei
- origine: corticosurrene, ovaio e testicolo
- esempi: testosterone, estradiolo, aldosterone, vitamina D
3. Amine e ormoni tiroidei
Sono molecole aminoacidiche come i peptidi e le glicoproteine, ma molto più piccoli di questi!
Quindi, possono attraversare le membrane biologiche nonostante la carica
-
derivazione: aminoacido tirosina
-
A. catecolamine
epinefrina: prodotta dalla midollare del surrene
noradrenalina: prodotta dalle terminazioni nervose simpatiche
localizzazione recettoriale: recettori di membrana
-
B. Ormoni tiroidei: prodotti dalla tiroide
T4 = tiroxina (biologicamente inattiva)
T3 = triiodotironina (biologicamente attiva)
Localizzazione recettoriale: nucleare
4.
-
Neurotrasmettitori
Derivazione: neuroni con rilascio nello spazio intersinaptico
Origine: neuroni
Localizzazione recettoriale: membrana postsinaptica
Esempi: acetilcolina, dopamina, serotonina ecc.
5. Prostaglandine
- Derivazione: acidi grassi, ma nonostante questo sono molecole cariche e quindi incapaci di
attraversare la membrana cellulare (salvo alcuni)
- Origine: numerosi tessuti
- Localizzazione recettoriale: membrana cellulare
- Esempi: PGE2, PGF2, ecc.
Meccanismi di regolazione della secrezione ormonale
1. Connessioni ipotalamo-ipofisarie
- Ipofisi posteriore (neuroipofisi): rilascio diretto dalle terminazioni nervose ipotalamiche di
ADH e ossitocina
- Ipofisi anteriore (adenoipofisi): “releasing hormones” ipotalamici stimolano la secrezione
ormonale ipofisaria
2. Simpaticoadrenergico
- Innervazione simpatica diretta della midollare del surrene
3. Innervazione autonomica di ghiandole endocrine
- I.e.: ipofisi anteriore e posteriore, pancreas, tiroide, gonadi ecc.
Attenzione: si sa poco di quello che fa!
Seconda ora
Con calma.
Il feedback negativo
Consideriamo l’asse ipotalamo-ipofisario in riferimento agli ormoni tiroidei.
Un ormone stimola la secrezione di un altro ormone e quest’ultimo inibisce la secrezione del primo
ormone.
Esempio: la T4, rilasciata dalla tiroide sotto lo stimolo della tireotropina (TSH), inibisce a sua volta
la sintesi e la secrezione
- di TSH a livello dell’ipofisi anteriore
- e di TRH nell’ipotalamo.
Immaginiamo che per un motivo come l’asportazione della tiroide il T4 cali: avremo ipotiroidismo
(bassi livelli di T4) e di conseguenza un aumento di TSH (vanno sempre in senso inverso!).
Allora l’endocrinologo interviene somministrando per bocca T4. E il TSH scende. Se la dose,
tuttavia, di ormoni tiroidei somministrata dall’endocrinologo è eccessiva, avremo T4 troppo alto
(ipertiroidismo, qui iatrogenico) e TSH troppo basso.
Se invece la tiroide inizia a produrre un eccesso di ormoni tiroidei, come nella malattia di GravesBasedow, anticorpi attivano il recettore del TSH stimolando la tiroide mimando il TSH stesso. E
avremo dunque ormoni tiroidei alti e TSH basso, tando da essere indosabile.
E il TSH è così sensibile a queste fini variazioni di ormoni tiroidei prodotti che si alza o si abbassa
ancor prima che noi riusciamo a misurare variazioni degli ormoni tiroidei circolanti. Abbiamo
situazioni iniziali, per esempio di ipofunzione tiroidea, in cui, se andiamo a misurare la T4, la
troviamo perfettamente nel range di normalità, ma con il TSH già più elevato. Ciò vuol dire che il
tireotropo ha già sentito che gli ormoni tiroidei stanno diminuendo.
Sono, queste, le condizioni subcliniche della patologia endocrina, laddove proprio questa sensibilità
è usata per rilevare le variazioni indosabili e sublciniche dei livelli di diversi fattori.
L’ormone tiroideo, però, è in grado di inibire non solo la secrezione di TSH ma anche quella di
TRH. E si verifica un doppio feedback che ha la stessa funzione: regolare continuamente la
secrezione di ormoni tiroidei.
Ripetiamolo: nel 99% dei casi i livelli di TSH e TRH variano inversmente a quelli degli ormoni
tiroidei.
Consideriamo l’esposizione al freddo intenso: il sistema nervoso centrale è stimolato a produrre una
grande quantità di TRH a livello ipotalamico, che determina produzione di TSH, che aumenta il
rilascio degli ormoni tiroidei, che ovviamente da un lato aumentano il metabolismo basale,
dall’altro stimolano molti sistemi di consumo ATP, e inoltre funzionano da disaccoppianti.
Pulsatilità e ritmo circadiano
Gli ormoni hanno fasi della giornata in cui sono secreti in maggiore quantità e altre in cui sono
secreti in minor quantità. Tipicamente,
- cortisolo, che di solito ha il punto più basso intorno alle 23.00-24.00, tanto che il prelievo
alle 24.00 si usa per escludere o confermare la diagnosi di sindrome di Cushing: se non
diminuisce molto a mezzanotte, c’è il sospetto di ipersecrezione di cortisolo (il ritmo,
tuttavia, varia da individuo a individuo)
- LH
- Una serie di ormoni ipofisari che tendono ad aumentare durante la notte (e in particolare
durante le fasi REM) e diminuire durante il giorno (ormone della crescita, TSH). È noto il
detto delle nonne che i bambini crescono quando dormono.
Trasporto plasmatico
1. Ormone libero vs. ormone legato
- gli ormoni steroidei e tiroidei ed alcuni ormoni peptidici sono trasportati nel sangue
prevalentemente legati a specifiche proteine plasmatiche di trasporto (“binding globulins”)
- soltanto l’ormone non legato o libero (“free”) è capace di raggiungere le cellule bersaglio
esercitando la specifica azione biologica: quindi [free hormone] < [total hormone]
esempio: [free T4 o FT4] = 0.03% [T4 totale]
- la quota di ormone libero e quella legata sono in equilibrio fra loro secondo la legge di
azione di massa
Questa forma di trasporto legata alle proteine degli ormoni rappresenta dunque da un lato una sorta
di riserva circolante, in equilibrio con l’altra molecola legante che è il recettore della forma libera,
la quale è contesa fra i due.
Per molti ormoni, quali i tiroidei, la parte libera e quindi capace di interagire con le cellule bersaglio
è una quota irrisoria del totale.
Ci sono situazioni in cui la quantità totale di T4 aumenta enormemente, per esempio la gravidanza:
la TBG è prodotta dal fegato, e la sua produzione è stimolata dagli estrogeni. Per cui le donne in
gravidanza, soprattutto nel primo trimestre (con grande quantità di estradìolo) hanno alti livelli di
TBC e troveremo alta la quantità totale di T4, quasi a livello ipertiroideo, mentre troveremo
normale la quantità libera.
2. Sinergiscmo vs antagonismo ormonale
- sinergismo = azione biologica combinata di due ormoni è maggiore della somma
dell’azione singola di ciascun ormone
- antagonismo = due ormoni svolgono effetti biologici opposti
Esempio: insulina e ormone della crescita (GH)
- entrambi stimolano la sintesi proteica, sono cioè anabolizzanti (e in questo sono sinergici)
- l’insulina stimola il trasporto del glucosio nel muscolo, mentre il GH lo inibisce (e in questo
sono antagonisti)
3. I recettori ormonali
-
I recettori ormonali sono molecole proteiche ad elevata specificità di struttura e funzione,
localizzati a livello delle cellule bersaglio dell’azione ormonale
Essi sono capaci di interagire con una specifica molecola ormonale, di interpretare il segnale
ormonale e di trasferirlo all’interno della cellula bersaglio convertendolo in una specifica
azione biologica (attivazione o inibizione genica)
-
Proprietà recettoriali.
o Sensibilità tissutale: maggiore è la capacità di legame ormonale (i.e., maggior
numero di recettori disponibili) più sensibile sarà il tessuto bersaglio ad una stessa
concentrazione ormonale
o Un meccanismo di protezione con cui la cellula si protegge dall’eccesso di un
ormone è la diminuzione della densità di recettori (per internalizzazione e
degradazione) in conseguenza di un carico eccessivo dell’ormone).
o Come già detto, spesso gli ormoni non trovano un solo tipo di recettore, ma diverse
isoforme recettoriali. Dell’insulina ci sono vari recettori: A, B e così via; non solo,
ma il recettore dell’insulina è simile a quello dell’IGF1 o somatomedina, tanto che si
possono formare recettori ibridi, fatti per metà da recettore dell’insulina e per metà
da recettore del’IGF1, e che sono capaci di legare l’uno e l’altro ormone, ovviamente
con proprietà diverse da quelle della versione “pura” di ciascuno dei due recettori. E
questo ha risvolti notevoli anche per la crescita neoplastica. All’interno delle cellule,
dunque, c’è in questo caso una regolazione quantitativa e non solo qualitativa.
o Regolazione qualitativa. Il recettore alfa 2 degli ormoni tiroidei non lega l’ormone
ma lega il sito genico che è target dell’ormone tiroideo, impedendo ad esso di
legarlo, e dunque ha un’attività inibitoria. Si è visto che la quantità di questi recettori
diminuisce nell’arco della vita, cosicchè il cervello di un anziano è molto meno
responsivo agli ormoni tiroidei del cervello di un fanciullo. Questo spiega la relativa
resistenza del sistema nervoso centrale dell’anziano all’azione degli ormoni tiroidei.
Ripetiamolo: ciò che conta non è solo la quantità totale dell’ormone, ma solo quella attiva; e non
conta neanche solo questa, ma la quantità di recettori e la loro qualità. Addirittura, nello stesso
soggetto potremo avere, per esempio, ipotiroidismo nel fegato e ipertiroidismo nel cuore, se in quel
soggetto la distribuzione tissutale delle varie isoforme del recettore è sproporzionata rispetto al
normale.
Secondi messaggeri
-
Il più famoso e frequente è l’AMP ciclico. Esempi: epinefrina e norepinefrina, glucagone,
LH, FSH, TSH, caclitonina, PTH, ADH
Attività protein chinasica: insulina, ormone della crescita, prolattina, ossitocina,
eritropoietina, molti fattori di crescita
Calcio e/o fosfoinositidi: epinefrina e norepinefrina quando si legano ad alcuni tipi di
recettori, angiotensina II, ADH, GnRH, TRH
GMP ciclico: peptide natriuretico atriale, ossido nitrico
Tipi chimici di recettori di membrana
- attraversano una sola volta la membrana: es. recettore dell’EGF (fosforilazioni in
Serina/Treonina)
- dimeri, cioè fatti da due subunità che attraversano ciascuna una volta la membrana,
accostate l’una all’altra: es. recettore dell’insulina (fosforilazioni in tirosina)
- attraversano sette volte la membrana e hanno poi una lunga coda intracitoplasmatica. Sono i
recettori legati alle proteine G e nella maggior parte dei casi usano come secondo
messaggero l’AMP ciclico: es. recettore beta-adrenergico di tipo 1 (il primo ad essere stato
identificato)
Il meccanismo d’azione dei GPCR = recettori accoppiati alle proteine G
Sono recettori a sette domini transmembrana che prendono contatto con le proteine G, trimeriche,
con subunità alfa, beta e gamma, che hanno la capacità di legare il GTP trasformandolo in GDP.
In situazione quiescente, la proteina G è legata al versante intracellulare del recettore, e la subunità
alfa lega GDP.
Quando arriva l’ormone, si lega al recettore provocandone una mutazione conformazionale, e allora
la subunità alfa rilascia il GDP e lega GTP, le subunità inibitorie beta e gamma si staccano dalla
alfa, e la alfa libera e legante GDP attiva l’adenilato ciclasi che usa ATP come substrato per
produrre AMP ciclico.
La principale molecola bersaglio del cAMP è la PKA = protein chinasi A: il cAMP lega la subunità
catalitica inattiva, spiazza la subunità regolatoria, cosicché la subunità catalitica è attiva e stimola
fosforilazione di varie proteine, dando origine a varie risposte cellulari. Studi italiani dimostrano
che la subunità catalitica stessa può arrivare direttamente dentro il nucleo.
Il meccanismo d’azione del recettore dell’insulina
Ciascun monomero è dato da una subunità alfa extracellulare e una beta transmembrana. Il legame
dell’ormone al dimero determina autofosforilazione su residui di tirosina del versante
intracitoplasmatico, il che fa partire una cascata di signaling intracellulare.
Recettori nucleari
Prendiamo l’esempio degli steroidi o degli ormoni tiroidei: li possiamo già incontrare nel
citoplasma. Lo steroide arriva, entra nella cellula attraverso la membrana plasmatica, incontra il suo
recettore nel citoplasma, ci si lega, insieme i due entrano nel nucleo, e lì si influenza la trascrizione
genica. Nonostante questo percorso sia semplificato, è chiaro che abbiamo bisogno di molte meno
mediazioni.
Con gli ormoni tiroidei, come dicevamo, il recettore è già sul DNA.
Tutti i recettori che agiscono direttamente sul DNA si somigliano molto strutturalmente:
superfamiglia dei recettori steroidei e tiroidei:
- una parte centrale è molto conservata, tant’è che essi sono stati clonati proprio sfruttando
questa similitudine a livello dell aparte centrale, che è la parte che interagisce con il gene
bersaglio sul DNA
- la parte carbossiterminale è il sito di legame dell’ormone: come è ovvio, è questa la parte
che varia di più da recettore a recettore
- la parte amminoterminale è regolatoria, in grado di interagire con altri fattori trascrizionali;
nella regolazione, è importante per trasferire l’attivaizone all’RNA polimerasi di tipo II. Dal
recettore all’RNA polimerasi di tipo II c’è tutto un discorso fatto ttraverso la mediazione di
fattori recettoriali di trascrizione.
Il DNA-binding domain possiamo immaginarlo fatto come una spina elettrica. Ha due protuberanze.
Si chiama struttura a dita di Zinco = Zinc finger, perché un atomo di Zn le lega.
Questa struttura del DNA varia da recettore a recettore in base al numero di nucleotidi che separano
le due dita di zinco. Ce ne saranno di un certo tipo, altre che sono più larghe, e così via: è la
distanza, il passo fra le due protuberanze che differenzia una spina dall’altra.
Sul gene bersaglio di questi recettori ci sono delle sequenze nucleotidiche molto caratteristiche
denominate HRE = Hormone Response Element (TRE, ERE, ARE e così via per ormoni come,
rispettivamente, i tiroidei, gli estrogeni, gli androgeni). L’HRE possiamo immaginarlo come la
presa della corrente. Anche l’HRE ha un suo passo: degli elementi in grado di interagire con i
recettori che in mezzo possono essere distanziati da un numero variabile di nucleotidi. Quindi, in
base al passo della presa si realizza la specificità di interazione. Capiamo bene, però, che su queste
situazioni c’è possibilità di overlapping. E questo ci spiega perché gli steroidi tendono ad avere
azioni simili. Lo sterolo ha azione mineralcorticoide, i progestinici sono maschilizzanti, e così via.
Una sorta di tendenza alla bassa specificità del sistema, quando un ormone è presente in quantità
eccessva e inizia ad agire su recettori di ormoni simili (es. donne che prendono la pillola e hanno
un’enorme ritenzione idrica perché l’ormone mima i mineralcorticoidi).
POST SCRIPTUM
Abbiamo chiesto le slidesssss al prof, che ci ha assicurato che le metterà sul blackboard. Non
contenti di questa soluzione e smaniosi di appropriarcene, io e Davide abbiamo deciso di insistere
affinché il prof ce le dia subito alla fine di una delle prossime lezioni su una delle nostre penne
USB, affinché possiamo poi diffonderle con sorprendente velocità con il solito metodo (cioé:
nell’assai frequente caso che io venga a lezione, chi vuole viene a controllare sul mio PC se io non
abbia niente di nuovo, e si prende le cose che non ha ancora...). Adesso andate pure a dare un senso
alla vostra giornata (o alla vostra settimana, così vi togliete il pensiero per i prossimi sette giorni, un
po’ come gli aggiornamenti dell’antivirus). Ciao a tutti!
Luca Valerio