Principali difficoltà nello studio dei caratteri genetici
nell’uomo
Non si possono programmare gli incroci
Il tempo di generazione dello sperimentatore è
uguale al tempo di generazione della specie oggetto
di studio
Le fratrie sono di piccole dimensioni
Vantaggi
Per l’uomo le conoscenze ‘mediche’ (anatomia,
fisiologia, patologia ecc.) sono molto più avanzate
rispetto a quelle sugli altri organismi
Anche fenotipi molto rari è difficile che sfuggano
all’osservazione
Lo studio dei caratteri mendeliani nell’uomo viene
effettuato attraverso lo studio dei pedigree
Non sempre è facile dimostrare l’ereditarietà
genetica di un carattere membri della stessa
famiglia oltre a condividere una certa quota del loro
patrimonio genetico condividono anche l’ambiente
(alimentazione, stile di vita, interessi, ecc.) e non
sempre è possibile avere informazioni dettagliate e
complete sui rapporti di parentela e sul fenotipo dei
vari membri di una famiglia
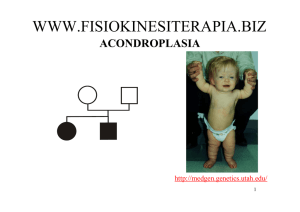
Albero genealogico o pedigree figura che
rappresenta in modo schematico individui con almeno
un ascendente in comune, i loro discendenti, i loro
coniugi e le relazioni di parentela che intercorrono
tra di essi
Possono essere utilizzati:
per accertare le modalità di trasmissione di un
carattere ereditario
nella consulenza genetica (es. per calcolare il
rischio di generare figli affetti o il rischio di
sviluppare una determinata malattia genetica)
sono fondamentali per la mappatura genetica
Se si utilizzano i pedigree per stabilire le modalità di
trasmissione di un carattere genetico è importante
considerare la frequenza con cui il carattere in esame si
presenta nella popolazione
se un CARATTERE è RARO possiamo assumere che sia
entrato nel pedigree una sola volta e, tra tutte le modalità
di trasmissione compatibili con il/i pedigree in esame,
considerare come più verosimile quella che ipotizza il minor
numero di portatori indipendenti (cioè non imparentati)
viceversa per CARATTERI COMUNI tale assunzione non può
essere fatta
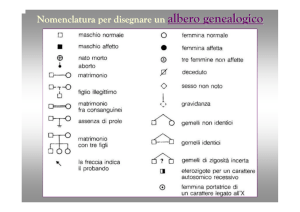
I PRINCIPALI SIMBOLI USATI PER LA
COSTRUZIONE DEI PEDIGREE
Il probando è l’individuo attraverso cui si è arrivati allo studio di
quel pedigree, viene indicato con una freccia
Ogni individuo è identificato in modo non ambiguo da una
coppia di numeri (un numero romano ed uno arabo)
PRINCIPALI MODALITA’ DI TRASMISSIONE DI UN
CARATTERE MENDELIANO
autosomica dominante
autosomica recessiva
X-linked dominante
X-linked recessiva
Y-linked
mitocondriale
dovuta a un gene soggetto a imprinting
EREDITA’ AUTOSOMICA DOMINANTE
EREDITA’ AUTOSOMICA RECESSIVA
EREDITA’ X-LINKED RECESSIVA
U
Dipartimento di Biologia
Università degli Studi di Roma “Tor Vergata”
EREDITA’ X-LINKED DOMINANTE
Tra gli affetti il rapporto maschi:femmine è 1:2
EREDITA’ Y-LINKED
trasmissione maschio-maschio
non si conoscono (e probabilmente non esistono) patologie legate
a geni del cromosoma Y (escluse quelle legate alla fertilità)
FATTORI CHE COMPLICANO L’ANALISI DI
PEDIGREE
penetranza incompleta – La penetranza è la percentuale di
individui che hanno il genotipo-malattia e che sono affetti.
Affermare che una malattia genetica è a penetranza
incompleta equivale a dire che esiste una certa quota di
individui che non manifestano la malattia pur avendo il
genotipo-malattia.
Esempio 1. malattia AD con una penetranza dell’80%
100 soggetti Aa, 80 sono malati e 20 sono sani
Esempio 2. malattia AR con una penetranza del 70%
100 soggetti aa, 70 sono malati e 30 sono sani
ESEMPIO DI MANCATA PENETRANZA
madre e figlia di II-2 presentano la stessa patologia possiamo escludere
che il gamete da cui è nata la figlia di II-2 portasse una mutazione fresca
U
Dipartimento di Biologia
Università degli Studi di Roma “Tor Vergata”
Penetranza incompleta
La penetranza incompleta è una caratteristica
maggiormente frequente nelle malattie AD
Il motivo per cui individui con il genotipomalattia non sono malati può essere:
influenza dell’ambiente
azione di altri geni (gene principale + geni
modificatori)
ESPRESSIVITA’ VARIABILE
individui portatori dello stesso allele
malattia presentano caratteristiche cliniche e
gravità diverse
Per le malattie X-linked l’espressività variabile, nelle
femmine, può essere particolarmente accentuata. Questo
è dovuto al fenomeno di inattivazione del cromosoma X:
la gravità del quadro clinico dipenderà dalla proporzione
di cellule che hanno mantenuto attivo il cromosoma X
con l’allele anormale
Espressività variabile e penetranza incompleta: sono un
continuum dello stesso fenomeno.
La penetranza incompleta è una forma estrema di
espressività variabile.
ESPRESSIONE FENOTIPICA
OGNI OVALE RAPPRESENTA UN INDIVIDUO
INDIVIDUI DELLA STESSA RIGA HANNO UGUALE GENOTIPO
PENETRANZA INCOMPLETA
ESPRESSIVITA’ VARIABILE
PENETRANZA INCOMPLETA E ESPRESSIVITA’ VARIABILE
ETEROGENEITÀ GENETICA (eterogeneità di locus)
fenomeno per cui malattie clinicamente uguali sono
dovute a mutazioni in geni diversi
Esempi classici sono la sordità non sindromica e l’albinismo
cioè fenotipi alla cui determinazione concorrono numerosi eventi sequenziali,
ognuno controllato da un gene, quindi il non verificarsi di uno qualsiasi di essi
compromette del tutto il fenotipo finale
Altri esempi:
sindrome di Ehlers-Danlos (lassità di pelle e legamenti) e retinite
pigmentosa: varie forme clinicamente indistinguibili ma
geneticamente distinte (eredità AD, AR e X-linked recessiva);
Quando si cerca di stabilire la modalità di trasmissione di
una particolare malattia o si vuole mappare il genemalattia bisogna tenere presente questa possibilità e
considerare che in pedigree diversi la patologia può
essere causata da mutazioni in geni diversi
ESEMPIO DI COMPLEMENTAZIONE NELL’UOMO
(Sordità autosomica recessiva)
Malattie recessive che presentano eterogeneità di
locus mostrano il fenomeno della complementazione
ETEROGENEITÀ GENETICA (eterogeneità allelica)
mutazioni diverse dello stesso gene causano malattie
diverse
talvolta la differenza è quantitativa (es. Fibrosi cistica
con e senza insufficienza pancreatica; distrofie di
Duchenne e di Becker);
altre volte è qualitativa (alleli diversi dello stesso gene
causano patologie molto diverse (es. insensibilità
agli androgeni e atrofia muscolo-spino-bulbare)
INSORGENZA TARDIVA
penetranza dipendente dall’età
1
probabilità
0,8
0,6
0,4
0,2
0
0
20
40
età (anni)
60
80
I
II
1
2
3
4
III
II-1, II-3 e II-4 non hanno ricevuto l’allele malattia o
non hanno ancora manifestato la malattia?
malattia X-linked dominante letale nei maschi
in alcuni casi si osserva poliabortività
il rapporto sessi è alterato ma, a causa delle piccole dimensioni
delle famiglie umane, generalmente è difficile dimostrarlo
malattia X-linked recessiva: mutazione fresca o
segregazione di un allele malattia ?
Mosaicismo
Presenza in un individuo di due linee cellulari
geneticamente diverse ma derivanti da un unico zigote
Mosaicismo somatico
Mosaicismo germinale
Mosaicismo somatico E germinale
Mosaicismo funzionale nelle femmine dovuto a
inattivazione del cromosoma X
Le malattie X-linked dominanti in genere mostrano,
nelle femmine, una notevole variabilità di espressione
dovuta alla diversa percentuale di cellule che hanno
inattivato il cromosoma X con l’allele mutante
L’inattivazione del cromosoma X può anche essere
responsabile dell’insorgenza di malattie X-linked recessive
in femmine portatrici.
pedigree di un carattere autosomico recessivo comune
carattere X-linked comune: apparente trasmissione maschio-maschio
esempio: daltonismo, freq. del carattere nei maschi (q) 0.08, freq. di
femmine portatrici (2pq) = 0.147, quindi la probabilità di un matrimonio
maschio daltonico x femmina portatrice = 0.08 x 0.147 = 0.012
EREDITA’ MITOCONDRIALE
CARATTERE DETERMINATO DA UN GENE SOGGETTO A
IMPRINTING NELLA MADRE (è attiva solo la copia fornita
dal padre): figli di madri malate non sono mai malati; figli di
padri malati hanno una probabilità del 50% di ricevere la
malattia