GLI ORMONI
pag. 1
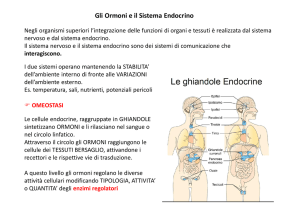
Ormoni
Definizione classica:
messaggeri chimici prodotti da tessuti specializzati (ghiandole
endocrine) che agiscono a distanza su cellule bersaglio (segnali
endocrini)
Definizione più ampia:
la capacità di produrre ormoni non è una proprietà esclusiva delle
ghiandole endocrine.
Gli ormoni possono anche agire localmente su cellule vicine per
diffusione (segnali paracrini)
o anche sulle stesse cellule che li hanno prodotti (segnali autocrini)
In questi casi l’ormone non è secreto in circolo
Secondo questa definizione più ampia la separazione tra
ormoni e neurotrasmettitori diventa labile
Certe molecole possono agire sia come ormoni che come
neurotrasmettitori (es: catecolamine, ormoni ippotalamici,
ipofisari, gastro-intestinali)
I neuroni possono rilasciare in circolo ormoni (neurormoni)
ossitocina
•
L'ossitocina è un ormone prodotto dai nuclei ipotalamici e secreto dalla
neuroipofisi
•
E’ un ormone peptidico, un nonapeptide, molto simile a un altro ormone ipofisario,
la vasopressina
•
L'azione principale dell'ossitocina è quella di stimolare le contrazioni della
muscolatura liscia dell'utero
•
Governa il "riflesso di eiezione fetale", "riflesso di eiezione del latte", "riflesso di
eiezione dello sperma”
•
Recettori dell'ossitocina si trovano anche nel sistema limbico del cervello
•
Esperimenti su animali hanno dimostrato l'importanza di tale ormone
nell'accoppiamento, nel comportamento parentale e sociale
GLI ORMONI-generalità
Gli ormoni peptidici e le catecolamine sono idrosolubili, non possono attraversare
la membrana plasmatica e trovano gli specifici recettori sulla superficie esterna
della membrana delle cellule bersaglio. Essendo “primi messaggeri” gli ormoni
producono i loro effetti grazie all’azione di un “secondo messaggero” intracellulare
l’AMP ciclico (AMPc)
derivato dall’ATP, che attiva allostericamente delle proteine cinasi in grado di
fosforilare con l’intervento di ATP altre proteine enzimatiche. La fosforilazione di
questi ultimi enzimi causa, a seconda dei casi, attivazione o si disattivazione.
L’azione di questi ormoni è rapida poiché agiscono modificando l’attività di enzimi
già presenti nelle cellule.
Gli ormoni tiroidei e steroidei sono invece in grado di attraversare la membrana
plasmatica, si legano a recettori citoplasmatici, raggiungono il nucleo ove attivano
la trascrizione, causando la sintesi di una proteina enzimatica.
L’azione di questi ormoni è lenta.
pag. 5
GLI ORMONI - Classificazione
- Ormoni di derivazione aminoacidica (tirosina)
catecolamine (adrenalina, nor-adrenalina), ormoni tiroidei
- Ormoni peptidici ( 3 - 200 AA)
ormoni dell’ipotalamo e dell’ipofisi, insulina, glucagone,
paratormone, calcitonina
- Ormoni steroidei (derivati del colesterolo)
ormoni della corteccia surrenale, androgeni, estrogeni
- Ormoni eicosanoidi (derivati dall’acido arachidonico)
prostaglandine
pag. 6
IL MECCANISMO D’AZIONE DEGLI ORMONI
PEPTIDICI E CATECOLAMINICI
ormone
Primo messaggero
Recettore proteico
Adenilico ciclasi
ATP
AMPc
PROTEINA
ENZIMATICA
Secondo messaggero
PROTEINA
ENZIMATICA
FOSFORILATA
pag. 7
IL MECCANISMO D’AZIONE DEGLI
ORMONI STEROIDEI
ormone
DNA
trascrizione
ormone
recettore
RNAm
traduzione
Sintesi della proteina enzimatica
pag. 8
L’INSULINA
L’insulina è prodotta nel pancreas dalle cellule delle isole del Langerhans e
viene secreta in risposta all’iperglicemia essendo il processo attivato da ioni
calcio. I tessuti bersaglio sono quello epatico, muscolare ed adiposo.
L’effetto dell’insulina è ipoglicemizzante.
Effetti metabolici
-Promuove l’ingresso del glucoso nel (fegato), nel muscolo e nel tessuto
adiposo
- Stimola la glicogenosintesi nel fegato e nel muscolo
- Stimola la glicolisi acetil-CoA sintesi acidi grassi nel fegato e nel
tessuto adiposo
- Favorisce la sintesi di VLDL nel fegato
- Inibisce la glicogenolisi nel fegato e nel muscolo
- Inibisce la gluconeogenesi nel fegato
L’abbassamento della glicemia causa diminuzione di emissione di insulina
con un meccanismo a feed-back.
pag. 9
IL GLUCAGONE
Il glucagone è prodotto nel pancreas dalle cellule delle isole del Langerhans;
viene secreto in risposta a ipoglicemia; il suo bersaglio principale è il fegato,
ma agisce anche sul tessuto adiposo.
Il suo effetto è iperglicemizzante.
Effetti metabolici
- Attiva la glicogenolisi (fegato)
- Stimola la lipolisi (tessuto adiposo)
- Attiva la gluconeogenesi (fegato)
- Inibisce la glicolisi (fegato, tessuto adiposo)
- Inibisce la glicogenosintesi (fegato)
Il glucagone è in grado di causare aumento della glicemia grazie alla presenza,
esclusivamente nel fegato, dell’enzima glucoso 6-fosfatasi che permette la
defosforilazione del glucoso 6-P e la fuoriuscita nel sangue di glucoso libero.
pag. 10
IL FEGATO E LA REGOLAZIONE DELLA
GLICEMIA
GLUCOSO DAL SANGUE
GLUCOSO LIBERATO NEL SANGUE
glucoso 6 fosfatasi
G6-P
glicogeno
glicogeno
fonte energetica
sintesi di acidi
grassi
G6-P
gluconeogenesi
lattato, AA
acidi grassi
utilizzati
come
combustibile
VLDL al
tessuto adiposo
INSULINA
DOPO UN PASTO
acidi grassi
dal tessuto
adiposo
GLUCAGONE
DOPO UNA NOTTE DI DIGIUNO
pag. 11
LE CATECOLAMINE
Le catecolamine, adrenalina e noradrenalina, sono prodotte nella midollare
delle surrenali ed anche nel cervello e nel tessuto nervoso, dove operano come
neuro-trasmettitori. Lo stimolo nervoso ne provoca l’emissione nel sangue e la
loro concentrazione da 0,06 microgrammi/litro si eleva, in pochi secondi, di
1000 volte. Si possono legare a recettori alfa e beta. La sintesi delle
catecolamine parte da tirosina DOPA Dopamina.
Effetti fisiologici: aumento del battito cardiaco e della pressione sanguigna. Il
bersaglio principale di questi ormoni è il muscolo, ma agiscono anche sul
fegato e sul tessuto adiposo.
Effetti metabolici
- Stimolano la glicogenolisi nel fegato aumento della glicemia
- Stimolano la glicogenolisi muscolare glicolisi (lattato) ATP
- Stimolano la gluconeogenesi nel fegato aumento della glicemia
- Mobilitano gli acidi grassi dal tessuto adiposo
- Stimolano la secrezione di glucagone
- Inibiscono la glicogenosintesi nel fegato e nel muscolo
- Inibiscono la secrezione di insulina
pag. 12
IL METABOLISMO DI VARI
ORGANI E TESSUTI
pag. 13
IL FEGATO: UN ORGANO ALTRUISTA
Il fegato:
presenta un numero di enzimi costitutivi
molto superiore agli altri organi
ha caratteristiche embrionali ed è rigenerante
contiene una riserva di glicogeno pari
a 400 kcal.
rifornisce i tessuti periferici di carburanti metabolici
detossifica l’organismo
Vie metaboliche e attività
Carburanti preferiti:
specifiche:
prodotti di degradazione
gluconeogenesi
di AA
chetogenesi
acido lattico
ureogenesi
sintesi di acido glucuronico
turnover rapido proteine plasmatiche
modulazione della glicemia
sintesi VLDL
pag. 14
IL FEGATO E LA REGOLAZIONE DELLA
GLICEMIA
GLUCOSO DAL SANGUE
GLUCOSO LIBERATO NEL SANGUE
glucoso 6 fosfatasi
G6-P
glicogeno
glicogeno
fonte energetica
sintesi di acidi
grassi
G6-P
gluconeogenesi
lattato, AA
acidi grassi
utilizzati
come
combustibile
VLDL al
tessuto adiposo
INSULINA
DOPO UN PASTO
acidi grassi
dal tessuto
adiposo
GLUCAGONE
DOPO UNA NOTTE DI DIGIUNO
pag. 15
IL MUSCOLO SCHELETRICO
Il muscolo scheletrico:
contiene elevate quantità di mioglobina
ha una riserva di glicogeno di 1200 kcal.
contiene fosfocreatina in concentrazione 6
volte superiore all’ATP (CPK isoenzima MM)
ha l’enzima miocinasi che rigenera
rapidamente 1 ATP da 2 ADP
allenato intensamente utilizza acidi grassi e
corpi chetonici al posto dei carboidrati
Carburanti preferiti:
dopo un pasto:
in digiuno di breve termine:
glucoso
acidi grassi dall’adiposo
corpi chetonici Leu, Ile, Val ( AA ramificati)
In condizioni di anossia si attivano:
glicogenolisi
glicolisi anaerobia
elevata liberazione di lattato
in digiuno prolungato:
corpi chetonici
lattato
Sotto sforzo si verifica:
incremento di consumo di O2
incremento di β-ossidazione di acidi grassi
incremento di fosforilazione ossidativa
elevata liberazione di lattato
pag. 16
IL CUORE
Il miocardio:
contiene elevatissime quantità di mioglobina per
sostenere l’intensa respirazione cellulare necessaria alla
contrazione incessante
consuma per il 50-90% del suo metabolismo acidi grassi
I carburanti preferiti si differenziano a
seconda delle condizioni, come nel muscolo
scheletrico:
dopo un pasto:
glucoso
corpi chetonici
in digiuno di breve termine:
acidi grassi dall’adiposo
Leu, Ile, Val (ramificati)
In condizioni di anossia si attivano:
glicogenolisi
glicolisi anaerobia
in digiuno prolungato:
corpi chetonici
lattato
Sotto sforzo si verifica:
incremento del consumo di O2
incremento della β-ossidazione
incremento della fosforilazione
ossidativa
pag. 17
IL TESSUTO ADIPOSO
Il tessuto adiposo bianco ha un ruolo centrale nel
metabolismo energetico. I trigliceridi contengono
principalmente: acido oleico (45%), palmitico
(20%), linoleico (10%) e stearico (6%).
Vie metaboliche attive:
• glicolisi
• sintesi di acidi grassi
• via dei pentosi
• termogenesi
La lipoproteina lipasi (adipociti) idrolizza i
trigliceridi dei chilomicroni e delleVLDL: ingresso
di acidi grassi poi esterificati negli adipociti.
I trigliceridi vengono idrolizzati durante il digiuno,
per rifornimento energetico.
Il tessuto adiposo bruno, ricco di mitocondri,
attua termogenesi nel neonato, l’adiposo
bianco nell’adulto.
La termogenesi è stimolata dall’adrenalina
che attiva la lipolisi e gli acidi grassi liberi
agiscono da disaccoppianti, annullando il
gradiente di protonico della catena
respiratoria, con dispersione di energia sotto
forma di calore.
pag. 18
IL RENE
La parte corticale e quella midollare del rene presentano
metabolismo molto differente:
la corticale presenta metabolismo aerobio con grande
consumo di ossigeno. E’ attiva la gluconeogenesi per
fornire glucoso alla midollare
la midollare è caratterizzata da metabolismo anaerobio
Carburanti principali
acido palmitico (60-80% energia)
Reazione caratteristica (ammoniogenesi)
Glutamina glutamato + NH4+
acido lattico
glutamina
glucoso
corpi chetonici (digiuno)
pag. 19
IL CERVELLO
Il cervello consuma:
nell’adulto il 25% dell’ossigeno corporeo
nel neonato e nel bambino (4 anni) il 50 %
dell’ossigeno corporeo
glucoso come unico combustibile
corpi chetonici a digiuno
L’anossia e l’ipoglicemia protratte sono letali
Il contenuto di AA liberi nel cervello è almeno 8 volte superiore a quello
plasmatico, per la sintesi di molti neurotrasmettitori.
Il livello di Asp e Glu è 300 volte superiore a quello plasmatico.
pag. 20
I POLMONI
I polmoni:
presentano attiva glicolisi anaerobia con produzione
elevata di lattato
consumano per sé pochissimo ossigeno per non
sottrarlo ai tessuti
in digiuno consumano acidi grassi e il metabolismo
fortemente aerobio sottrae ossigeno agli altri tessuti
presentano attiva via dei pentosi (NADPH + H+ )
sintetizzano attivamente acidi grassi
Sintesi caratteristica attuata dagli pneumociti di Tipo II:
sostanza surfactante, miscela di fosfolipidi e 3 tipi di lipoproteine,
responsabile della bassa tensione superficiale degli alveoli polmonari,
protettiva verso agenti ossidanti ( ossigeno, ozono, radicali liberi).
Il deficit di surfactante è causa di collasso alveolare.
pag. 21
LA CUTE
La cute:
è circa il 10% del peso del corpo umano
utilizza l’80% del glucoso nella glicolisi
contiene lattato in quantità 10 volte superiore
al sangue
sintetizza colesterolo per il rivestimento
superficiale
Reazioni caratteristiche:
attiva sintesi di cheratina
trasformazione del 7-deidrocolesterolo in colecalciferolo (vitamina D3)
sintesi di melanina a partire dall’aminoacido tirosina
pag. 22