43
Scheda: integrazione del metabolismo 2
Metabolismi specifici: il fegato, il tessuto adiposo e il muscolo scheletrico
Negli organismi pluricellulari e quindi anche nell'uomo, al quale faremo riferimento, ogni tessuto svolge
le vie metaboliche studiate in maniera specifica. Vi è quindi una "divisione del lavoro metabolico" che
sarà poi integrato e coordinato dall'azione degli ormoni adrenalina, glucagone ed insulina, che
tratteremo a parte. [vedi controllo ormonale].
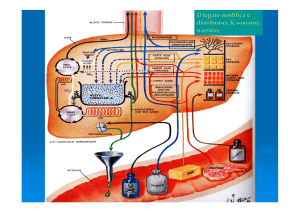
Il fegato
Il fegato è l'organo centrale del metabolismo in quanto gli epatociti, dopo l'assorbimento intestinale,
assumono la quasi totalità degli zuccheri e degli aminoacidi. Solo una piccola quota dei trigliceridi viene
utilizzata dagli epatociti mentre la gran parte, dopo l'assorbimento, entra nel tessuto linfatico e poi nel
tessuto adiposo. IL fegato elabora i nutrienti e li ridistribuisce ai tessuti, attraverso il flusso
sanguigno, in una miscela, "calcolata" per i fabbisogni extraepatici a seconda dell'intervallo di tempo tra
un pasto e l'altro, dell'attività dell'intero organismo e dal fabbisogno energetico.
Vediamo in dettaglio ora l'azione del fegato sugli zuccheri
e sugli aminoacidi. [seguire lo schema]
Il glucosio entrato nell'epatocita viene fosforilato dalla
glucochinasi, enzima con una Km molto elevata ma non
inibito dal suo prodotto, come l'esochinasi.
Il glucosio 6-P, come si vede in figura può avere cinque
destini metabolici che dipendono dalle necessità
dell'organismo.
1) In caso di glicemia bassa, il glucosio 6-P viene
defosforilato dalla fosfatasi per esportare glucosio libero
nel flusso sanguigno.
2)
Nel caso di bassa attività fisica e di normale
concentrazione ematica di glucosio, se le riserve epatiche di
glicogeno lo consentono, il glucosio 6-P viene immagazzinato
come glicogeno attraverso la glicogenosintesi.
3) In caso di richiesta energetica il glucosio-6-P prende la
via glicolitica e le successive deidrogenazioni per produrre
ATP nella catena respiratoria.
4) Se l'acetil-CoA è in eccesso esso servirà come
precursore per la sintesi di acidi grassi che saranno poi
incorporati nei fosfolipidi o nei trigliceridi.
5) Se si devono produrre coenzimi ridotti (NADPH) per
qualche sintesi oppure la cellula necessita di ribosio per la
biosintesi degli acidi nucleici, il glucosio 6-P è il precursore
della via del pentosio-fosfato.
44
Passiamo ora all'azione del fegato sugli aminoacidi [seguire lo schema]
Gli aminoacidi che entrano nelle cellule epatiche hanno diversi destini. Vediamoli.
1) sono i precursori per la sintesi delle
proteine epatiche o 2) per quelle tissutali.
3) alcuni aminoacidi sono i precursori di
nucleotidi, ormoni e neurotrasmettitori.
4) Gli aminoacidi in eccedenza per le biosintesi
dei punti precedenti vengono deaminati e
degradati ad acetil-CoA ed intermedi del ciclo
di Krebs che 4a) in caso di sufficiente energia
e/o di lavoro muscolare basso, possono essere
convertiti in glucosio attraverso la
gluconeogenesi.
L'acetil-CoA oltre a seguire la via del
metabolismo energetico, può essere utilizzato,
se in eccesso, 4c)per la sintesi degli acidi
grassi.
4d) L'ammoniaca rilasciata nella deaminazione
viene convertita in urea nel ciclo dell'urea.
5) Durante un digiuno prolungato, il livello di
glucosio ematico tende a scendere sotto i
livelli normali. Allora alcune proteine del
tessuto muscolare vengono degradate per
liberare aminoacidi liberi. Questi aminoacidi
per transaminazione cedono il gruppo
amminico al piruvato formando alanina che
viene trasportata al fegato che, a sua volta, la
deamina a piruvato.
Il piruvato, come sappiamo può percorrere la
via gluconeogenetica e produrre glucosio da
immettere nel sangue per ripristinare l'omeostasi glucidica. Questo ciclo, chiamato glucosio-alanina, è
quello che regola la glicemia tra un pasto e l'altro.
Per ultimo vediamo l'utilizzazione degli acidi grassi da parte del fegato.
Gli acidi grassi, nell'epatociti possono essere convertiti in lipidi, fosfolipidi e triacilgliceroli che, legati
alle lipoproteine plasmatiche, vengono esportati al tessuto adiposo.
Nella maggior parte dei casi gli acidi grassi sono il combustibile principale per il fegato e quindi
attraverso l'attivazione e la β-ossidazione vengono convertiti in acetil-CoA e NADH che poi produrrà
ATP nella catena respiratoria tramite la fosforilazione ossidativa. L'eccesso di acetil-CoA prodotto
viene convertito in corpi chetonici che rilasciati nel sangue vengono utilizzati dai tessuti extraepatici
per essere ossidati e produrre energia.
45
Il tessuto adiposo
E' il tessuto di deposito dei grassi provenienti dall'intestino, dopo l'assorbimento e quelli sintetizzati,
per la maggior parte, nel fegato ed esportati al tessuto adiposo tramite le VLDL. [vedi]
Quando l'organismo avverte la necessità di energia e lo zucchero ematico è in bassa concentrazione,
l'azione dell'ormone adrenalina attiva la lipasi contenuta nel tessuto adiposo che idrolizza i
triacilgliceroli in acidi grassi e glicerolo. Gli acidi grassi così liberati vengono trasportati nei tessuti
muscolo scheletrici e al cuore che li ossidano per estrarre ATP.
I muscoli scheletrici
I muscoli scheletrici, deputati a compiere un lavoro meccanico, sono la sede del maggior consumo
d'energia. Essi possono utilizzare acidi grassi, corpi chetonici o glucosio a seconda del grado di attività
muscolare. Nel muscolo a riposo i principali nutriente sono gli acidi grassi forniti dal tessuto adiposo. In
caso di moderata attività il muscolo può utilizzare invariabilmente corpi chetonici epatici, acidi grassi o
glucosio. Quando il muscolo esegue un'attività massima, l'apporto di ossigeno e di sostanze nutrienti
provenienti dal sangue, è insufficiente ai bisogni immediati. In tal caso il muscolo dopo aver demolito
glicogeno endogeno, demolisce il glucosio a lattato nella via anaerobica. Questo sforzo massimo può
continuare, dipendendo dall'allenamento, solo per poco tempo provocando accumulo di acido lattico che
deve essere smaltito. Dopo lo sforzo, il soggetto deve continuare in una respirazione accelerata per un
po' di tempo. La maggior parte dell'ossigeno così assunto viene utilizzato principalmente dal fegato per
produrre ATP da destinare alla sintesi conversione del lattato, giunto dal muscolo, in glucosio. (ciclo di
Cori).
Durante il periodo di intensa attività il muscolo scheletrico, che contiene fosfocreatina, può produrre
ATP nella reazione catalizzata dalla creatina chinasi:
fosfocreatina + ADP —→ creatina + ATP
ma durante la fase di recupero la fosfocreatina viene rigenerata tramite la reazione inversa:
creatina + ATP —→ fosfocreatina + ADP
Il cervello
Il cervello dell'adulto usa solo glucosio proveniente dal flusso sanguigno. La esochinasi, con una bassa
Km può utilizzarlo anche in concentrazioni basse che però non devono scendere sotto livelli critici che
comporterebbero gravi danni funzionali profondi ed irreversibili.
Durante i periodi di digiuno, il cervello può utilizzare anche il β-idrossibutirrato, un corpo chetonico
proveniente dal fegato.
L'energia dell'ATP è necessaria per creare e mantenere il potenziale d'azione generato dall'antiporto
della pompa sodio-potassio, ATPasi dipendente.