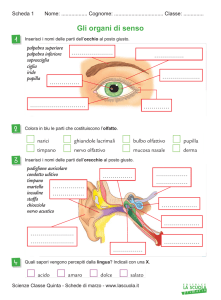
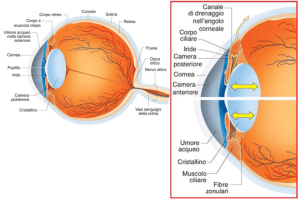
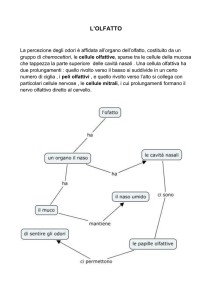
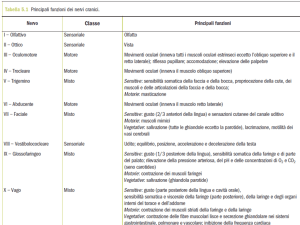
CERVELLO E COMPONENTI OLFATTIVE SECONDARIE NEI BASSI VERTEBRATI.
Biagio D’Aniello,
Aniello, Luciano Luongo,
Luongo, Maria di Meglio & Rakesh K. Rastogi
NEUROANATOMIA
L’evoluzione e la funzione dell’encefalo sono argomenti scientifici
scientifici che offrono difficoltà interpretative per i profondi cambiamenti
cambiamenti a cui l’organo è stato sottoposto dai bassi vertebrati ai mammiferi.
mammiferi. In
più, la rilevante plasmabilità encefalica e il suo rapido adattarsi a condizioni locali ha determinato
determinato diversificazioni sostanziali anche in specie molto affini.
affini. Lo studio della distribuzione di neuropeptidi
è uno strumento molto utile per tentare d’interpretare i meccanismi
meccanismi che sono alla base del motore evolutivo che ha determinato i cambiamenti del cervello; oltre a fornire valide indicazioni a
carattere funzionale. A tale scopo, nel nostro laboratorio siamo da diversi anni impegnati nello studio della distribuzione di alcuni
alcuni neuropeptidi nell’encefalo dei vertebrati al fine di fornire un piccolo
contributo alla comprensione delle controverse problematiche neuroanatomiche.
neuroanatomiche.
GnRH
Fattore di rilascio per le gonadotropine ipofisarie (gonadotropin
(gonadotropin releasing
hormone:
hormone: GnRH).
GnRH). Decapeptide presente, in varie forme molecolari, nel
sistema nervoso centrale e periferico dei vertebrati ed è stato rilevato
anche in alcuni gruppi d'invertebrati. Il suo ruolo, storicamente
storicamente
riconosciuto, è quello di stimolare la secrezione delle gonadotropine
ipofisarie nei vertebrati, ma è anche un neurotrasmettitore.
FMRFammide
Tetrapeptide cardioeccitatorio dei molluschi, isolato da un ganglio di un
mollusco. Presente, in forme varianti, nel sistema nervoso dei vertebrati.
vertebrati.
Queste forme sono riconosciute da con anticorpi diretti contro la
la parte ccterminale ammidata,
ammidata, conservata in tutti i peptidi appartenenti alla
famiglia degli FMRFammidi.
FMRFammidi. Si suppone che questi peptidi svolgano
funzioni neuromodulatorie,
neuromodulatorie, neurotrasmettitorie e neurormonali.
neurormonali.
Nello storione, un osteitto con caratteristicjhe di
primitività e il pesce zebra, un osteitto molto
derivato il pattern di sviluppo del sistema
FMRFammide segue lo stesso schema.
I dati ottenuti mediante cromatografia liquida ad alrta pressione
(HPLC), radioimmunodosaggio (RIA) e immunoistochimica (IHC)
dimostrano che nella pachimedusa (una rana arboricola messicana)
coesistono almeno 3 diverse forme di GnRH.
GnRH.
Il profilo neuroanatomico di
distribuzione del GnRH negli Apodi
presenta un pattern molto primitivo.
L’analisi comparate dei dati sul sistema FMRFammide di una
testuggine e del caimano suggerisce che nel corso dell’evoluzione
dell’evoluzione il
sistema tende ad umentare la sua complessità (teoria della
parcellazione dei nuclei encefalici).
NPY
Neuropeptide Y (NPY), membro della famiglia dei polipeptidi
pancreatici. Presente nel sistema nervoso centrale dei molti vertebrati
vertebrati
in cui è implicato nei meccanismi della memoria, del comportamento,
comportamento,
nella regolazione dei ritmi circadiani,
circadiani, nel controllo della pressione
sanguigna e della temperatura corporea, nella sintesi e rilascio di
alcuni ormoni ipofisari e nel controllo della pigmentazione negli
negli anfibi.
Lo sviluppo e l’organizzazione generale del sistema NPY
presentano rimarchevoli differenze con specie molto affini sotto
l’aspetto evolutivo.
Retina
Mesencefalo
Mesencefalo
Telencefalo
Telencefalo
Diencefalo
Nuclei NPY immunoreattivi nell’encefalo e nella retina
della spigola. Il sistema presenta una notevole
complessità.
Una specie, quattro cervelli! Nei salmoni alcuni nuclei
encefalici presentano due diverse morfologie e dimensioni
nelle femmine e due nei maschi.
NERVO TERMINALE
Il nervo terminale o nervo cranico 0 (NT) fu identificato per la
la prima volta come nervo cranico soprannumerario in un piccolo squalo
squalo (Fritsch
(Fritsch,, 1878). In seguito, un nervo con caratteristiche simili è
stato identificato nel dipnoo Protopterus (Pinkus,
Pinkus, 1894). Più tardi, Locy (1903) ne ha descritto la struttura in una varietà di squali e razze. Esso presenta gangli o singoli neuroni lungo il suo
percorso e si estende generalmente dall’area olfattiva all’encefalo
all’encefalo anteriore (Locy
(Locy,, 1905). L’identificazione del NT nei tetrapodi e nella maggior parte dei pesci ossei è molto difficoltosa poiché,
poiché, in
questi gruppi, esso è strettamente associato ai nervi olfattivo e/o vomeronasale;
vomeronasale; quindi, non appare come una struttura separata. Peraltro, in questi
questi gruppi non sono inequivocabilmente evidenti i
gangli lungo il suo percorso. In questi casi l’immunoistochimica
l’immunoistochimica per alcuni neuropeptidi consente di visualizzare il NT quando è incluso nel nervo olfattivo
olfattivo e nel cervello. Le conoscenze sulla
funzione e sull’evoluzione del NT nei vertebrati sono tuttora piuttosto
piuttosto limitate e le interpretazione neuroanatomiche controverse. Il nostro laboratorio partecipa al dibattito scientifico
scientifico sull’argomento
con ricerche in pesci, anfibi e rettili; in particolare, evidenziando
evidenziando e studiando la neuroanatomia del NT mediante immunoistochimica per il tetrapeptide cardioeccitatorio dei molluschi
(FMRFammide)
FMRFammide) il neuropeptide Y (NPY) e il fattore di rilascio per le gonadotropine ipofisarie (GnRH
(GnRH)) nell’area nasale e nell’encefalo anteriore.
Gambusia affinis
Distribuzione degli elementi FMRFa immunoreattivi nel pesce Moenkhausia sanctaefilomenae
Pangasius hypothalumus
Paracheirodon innesi
Gyrinocheilus aymonieri
A
Qualcosa sugli anfibi: Bufo
B
C
Nella gambusia,
gambusia, che presenta
un bulbo olfattivi sessile, i
neuroni del NT formano un
unico nucleo compatto alla
base del telencefalo anteriore.
A: neuroni bulbari FMRFaFMRFa-ir del NT con proiezioni dirette posteriormente.
Scale bar 200 µm; nell’inserto 1 mm.
B: fibre FMRFaFMRFa-ir del NT a livello ipotalamico.
ipotalamico. Scale bar 100; nell’inserto 1
mm.
Nei dipnoi il nervo terminale presenta
due vie separate. In questo lavoro è
precisata l’origine neuroanatomica
delle due radici da un unico ganglio ed
è proposta un’ipotesi innovativa
sull’evoluzione del NT.
Nello squaletto,
squaletto, con bulbi
olfattivi fortemente peduncolati,
i neuroni del NT sono dispersi
lungo il nervo olfattivo, il
peduncolo olfattivo e la regione
basale det telencefalo, ma non
formano aggregati consistenti.
Nel neon, che ha bulbi olfattivi
sessili vi sono due aggregati di
neuroni del NT. Il primo nel
bulbo olfattivo anteriore,
proprio all’ingresso del nervo
olfattivo; il secondo e nel bulbo
olfattivo posteriore.
In Gyrinocheilus,
Gyrinocheilus, con bulbi
olfattivi peduncolati, i neuroni
del NT tendono ad essere
molto anteriori, ma qualche
neurone è anche presente a
livello del telencefalo basale.
Nei teleostei il nervo terminale è anche indicato come sistema olfattoretinale,
olfattoretinale, perché oltre a proiettare nella regione
nasale raggiunge anche la retina. I corpi cellulari dei neuroni del nervo terminale sono distribuiti tra il nervo olfattivo, i
bulbi e il telencefalo basale. Il nucleo del telencefalo basale che non è presente in tutte le specie è detto nucleo
olfattoretinale.
olfattoretinale. La distribuzione dei neuroni del NT è condizionata dal tipo di bulbi olfattivi: se i bulbi olfattivi sono sessili i
neuroni tenderanno ad essere più anteriori.
C: fibre FMRFaFMRFa-ir a livello del nervo olfattivo. Scale bar 100 µm; nell’inserto
1 mm.
Il corpo cellulare dei neuroni del NT di questo pesce possono raggiungere
raggiungere i
100 µm e sono i più grandi mai osservati nel telencefalo dei vertebrati.
vertebrati.
Negli anfibi anuri adulti il nervo terminale è poco sviluppato,
mentre nei girini è piuttosto consistente, in quanto funge da
substrato migratorio per molti neuroni neuropeptidergici che
si originano nel placode olfattivo e migrano nell’encefalo,
appunto attraverso in nervo terminale. Nelle immagini in A un
girino di rospo placodectomizzato precocemente. Si osservi
come il lato sinistro in cui è stato rimosso il placode olfattivo
chirurgicamente sono assenti neuroni FMRFammide.
FMRFammide. In B
neuroni FMRFammide del NT nella mucosa olfattiva in via di
differenziamento e nel nervo olfattivo. In C neuroni del NT
lunbgo il bulbo olfattivo. D un placodectomizzato con
assenza del NT nel lato operato.
La presenza del NT viene riportata in un rettile
squamato solo durante lo sviluppo ontogenetico. Si
conferma quindi in questo lavoro l’unico dato esistente
nei rettili in letteratura sulla presenta transitoria del NT
in una tartaruga, pubblicato 100 anni prima.
SISTEMA DI FIBRE OLFATTIVE EXTRABULBARI
Nel 1991 Szabo e collaboratori hanno segnalato che alcune fibre del sistema olfattivo
olfattivo principale, anziché terminare, come di consueto, nei bulbi
bulbi olfattivi, li attraversano per invadere profondamente
il cervello. Originariamente essi indicarono queste fibre come “projection
“projection olfactive primaire extrabulbaire”
extrabulbaire” (extrabulbar
(extrabulbar olfactory projections = EBOP), più tardi rinominato “extrabulbar
“extrabulbar olfactory
system” (EBOS; Schober et al., 1994). Cadeva quindi l’idea che il bulbo olfattivo fosse l’unico
’unico
bersaglio
delle
fibre
originate
dai
recettori
primari
della
l
della mucosa olfattiva (Ramon y Cajal,
Cajal, 1911). La
presenza di un sistema con queste caratteristiche è stato dimostrata
dimostrata almeno durante lo sviluppo embrionale in tutti i vertebrati eccetto i rettili. E’ assente negli adulti amnioti.
amnioti. Le nostre ricerche si
sono articolate sull’utilizzo di marker neuronali capaci di migrare dal luogo di applicazione lungo le fibre nervose
nervose per cercare di studiare i dettagli della neuroanatomia dell’EBOS negli anfibi.
Pesce rosso: Carassius auratus
Pesce zebra: Danio rerio
Branzino: Dicentrarcus labrax
Santa filomena: Moenkhausia
sanctaefilomenae
Axolotl: Ambystoma mexicanum
Taricha granulosa
Rana temporaria
Rana esculenta klepton
Lucertola campestre: Podarcis sicula
28
37-46.
OB
AC
mb
Il tracciante posto nella mucosa
olfattiva della lucertola termina la
sua diffusione nel bulbo olfattivo
principale (OB) e in quello
accessorio (AOB), senza fibre
extrabulbari. Nella lucertola, così
come negli altri amnioti studiati
adulti non è presente un EBOS.
A
a
AC
Il sistema di fibre olfattive extrabulbari (EBOS) nelle quattro specie di teleostei mostrate segue lo stesso
pattern. Un tracciante neuronale (DiI)
DiI) è stato posto nella mucosa olfattiva (OM) in campioni fissati in
paraformaldeide. Il tracciante diffonde nelle membrane grazie alle
alle sue proprietà lipofiliche e raggiunge il
bulbo olfattivo (OB) migrando lungo il nervo olfattivo (ON). La maggior parte delle fibre olfattive primarie
terminano nel bulbo, ma alcune proseguono lungo il telencefalo medio
medio basale (TELmb
(TELmb)) fino a raggiungere
la commmessura anteriore (AC) dove si verifica una consistente decussazione.
decussazione. Nei teleostei il sistema di
fibre olfattive extrabulbari è molto più sviluppato nelle specie più primitive, nelle quali raggiunge il
mesencefalo. Nelle specie più derivate, come i teleostei in figura il sistema limita la sua estensione al
diencefalo. Le funzioni di questo sistema non sono ancora state chiarite.
OB
L’EBOS negli urodeli (sopra) evidenziato con il tracciante HrP posto nella
camera nasale dell’animale in vivo (sopra) raggiunge i bulbi olfattivi
olfattivi
principali (OB) e accessori (AOB); li attraversa e percorre il telecefalo
ventrale (TELv
), decussa consistentemente a livello della commessura
(TELv),
anteriore (AC), raggiunge il nucleo preottico (NPO) e si estende fino al
tegmento del mesencefalo (TEG). Stessa tecnica in Rana temporaria (a
destra); mentre in Rana esculenta è usato il DiI postmortem.
postmortem. Nei girini degli
anuri l’EBOS (a destra) risulta più sviluppato rispetto agli anuri
anuri adulti. In
pratica il sistema subisce una regressione evolutiva riprodotta durante lo
sviluppo embrionale. Con tutte le precauzioni del caso “l’ontogenesi
ricapitola la filogenesi”…
filogenesi”…
AC
TEG
La presenza di un EBOS nelle
specie acquatiche depone a favore
dell’ipotesi di una sua funzionalità
legata alla percezione delle
sostanze disciolte nell’acqua.
Tuttavia, noi mettiamo in dubbio
questa ipotesi, perchè la
regressione del sistema inizia già
nella linea evolutiva dei pesci,
quindi non legata alla transizione
dall’ambiente acquatico a quello
terrestre. Inoltre, l’EBOS è presente
anche in anamni che usano l’olfatto
in ambiente aereo.
Secondo alcuni autori L’EBOS è da
considerarsi una componente funzionale del
nervo terminale. In questo lavoro mostriamo
che i due sistemi hanno
diverse proprietà chimiche,
odologia e morfologia; di
conseguenza propendiamo
verso la loro separazione
funzionale.
A sinistra traccianti DiO
(verde) e DiI (rosso) posti
nel
cervello
marcano
alcuni neuroni recettoriali
olfattivi primari.