Famiglia Pinaceae
11 generi, 210 specie; monoiche; squame ovulifere con 2 ovuli, foglie da lineari a
aghiformi singole inserite direttamente sui macroblasti (Abies, Picea, Pseudotsuga,
Tsuga), oppure portate su un brachiblasto a gruppi di 2-5 (Pinus) o numerose (Cedrus,
Larix); semi di norma alati; polline saccato; squama ovulare e copritrice distinguibili
In Italia presenti allo stato spontaneo i generi Abies
(2 specie: A. alba e A. nebrodensis), Larix (L.
decidua) Picea (P. abies), Pinus (con 10 specie)
Famiglia Araucariaceae (sx) 3 generi (Agathis, Araucaria, Wollemia) e 33 specie
monoiche; squame ovulifere con 1 solo ovulo; foglie a lamina relativamente allargata
Wollemia nobilis è un “fossile vivente”, recentemente scoperto in Australia.
Famiglia Podocarpaceae (dx) 18 generi e 130 specie
foglie lineari; macrosporofilli isolati, con 1 solo ovulo, circondati da una squama carnosa
specializzata (epimazio), polline saccato; strobili con microsporofilli che portano 2
sporangi (sacche polliniche) ciascuno
Famiglia Cupressaceae – 30 generi (es. Cupressus, Juniperus, Sequoia, Taxodium,
Thuja etc.) e 133 specie
monoiche; macrosporofilli piani o peltati, opposti o verticillati (galbuli) con squame
ognuna contenente da 1 a 20 ovuli; i galbuli possono essere legnosi o carnosi
(Juniperus); foglie squamose o – raramente – aghiformi disposte su verticilli trimeri;
polline non saccato; squama ovulare e copritrice completamente fuse tra loro
In Italia presenti allo stato spontaneo i generi Cupressus
(con 1 specie) e Juniperus (con 5 specie)
Famiglia Sciadopityaceae
1 sola specie: S. verticillata. Polline non saccato
foglie ridotte a squame, semi alati
Famiglia Taxaceae – 6 generi (es. Taxus, Cephalotaxus) e 30 specie
dioiche; macrosporofilli isolati, con 1 solo ovulo, circondati da un involucro (arillo) –
derivante da una proliferazione dell’asse sotto l’ovulo - che a maturità diviene carnoso e
rosso; foglie da lineari a aghiformi; strobili con microsporofilli che portano 2-9 sporangi
(sacche polliniche) ciascuno
1 sola specie in Italia:
Taxus baccata L.
Ordine Gnetales
3 generi e circa 70 specie viventi. Sifonogamia. Alcune specie producono nettare.
Presentano trachee e foglie retinervie (Gnetum), doppia fecondazione (Ephedra),
proembrione cellulare (Ephedra e Welwitschia), scomparsa dell’archegonio (Gnetum,
Welwitschia) e particolare sviluppo dei tegumenti dell’ovulo a simulare uno “stilo”. Per
questi motivi sono stati da molti e per molti anni considerate filogeneticamente vicine
alle angiosperme.
In realtà non è proprio così: le Gnetales, risultano filogeneticamente vicine (se non
addirittura incluse!) alle Pinales, nonostante l’estremo livello di differenziazione. Le
trachee di Gnetum originano da fibrotracheidi, la doppia fecondazione in Ephedra non
dà origine alla produzione di un endosperma secondario (oltretutto, ciò è stato osservato
in passato anche in alcune Pinaceae e sembra essere un fenomeno strettamente
correlato alla sifonogamia), il proembrione cellulare è noto anche in una Pinales
(Sequoia). Infine lo “stilo” delle Gnetales ha un’origine completamente differente.
numero cromosomico di base x = 7 (Ephedra),
11 (Gnetum), 12 (Welwitschia). Abbastanza
frequenti i poliploidi.
Famiglia Ephedraceae (solo genere Ephedra, con 65 specie, 6 delle quali presenti in
Italia)
Arbusti delle regioni aride del Nordamerica e del bacino mediterraneo, con foglie
squamiformi e coni maschili con elementi sterili.
Famiglia Gnetaceae (solo genere Gnetum, con 30
specie)
Liane o piccoli arbusti dell'Asia sud-orientale, dell'Africa
tropicale e del bacino del Rio delle Amazzoni. Ha foglie
lanceolate simili a quelle delle angiosperme e ovuli non
riuniti in strobili.
Famiglia Welwitschiaceae
Una sola specie (W. mirabilis) endemica del deserto della Namibia. Ha foglie con
meristema basale.
….ma:
Ogni linea evolutiva da noi trattata a livello di ordine elevata a rango di sottoclasse!!!
Piante vascolari (tracheofite)
Sottoclasse Rhyniidae (tutte estinte)
Sottoclasse Lycopodiidae
crittogame vascolari afille o con microfilli
Sottoclasse Trimerophytidae (tutte estinte)
Sottoclasse Psilotidae
Sottoclasse Ophioglossidae
crittogame vascolari con megafilli
Sottoclasse Equisetidae
Sottoclasse Marattiidae
Sottoclasse Polypodiidae
“Progimnosperme” (tutte estinte)
“Pteridosperme” (tutte estinte)
Sottoclasse Pinidae
fanerogame o spermatofite
Sottoclasse Magnoliidae
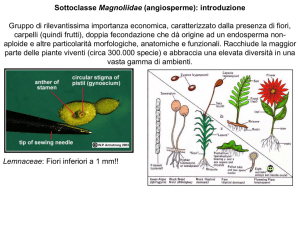
Sottoclasse Magnoliidae (angiosperme): introduzione
Gruppo di rilevantissima importanza economica, caratterizzato dalla presenza di fiori,
carpelli (quindi frutti), doppia fecondazione che dà origine ad un endosperma nonaploide e altre particolarità morfologiche, anatomiche e funzionali. Racchiude la maggior
parte delle piante viventi (circa 300.000 specie) e abbraccia una elevata diversità in una
vasta gamma di ambienti.
Wolffia (Araceae): Fiori inferiori a 1 mm!!
Rafflesia sp. (Rafflesiaceae)
fiore più grande al mondo
parassite (es. Cuscuta sp.,
Convovulaceae) alto a sx
saprofite (es. Monotropa sp.,
Ericaceae) al centro
carnivore (es. Drosera sp.,
Droseraceae) in basso a dx
Amorphophallus
titanum
(Araceae)
infiorescenza più
grande al mondo
alcune specie arboree
raggiungono dimensioni
eccezionali: es. Eucalyptus
jacksonii (Myrtaceae)
angiosperme acquatiche: es. Posidonia
oceanica (Posidoniaceae) una
monocotiledone endemica del Mediterraneo
che forma vaste “praterie” estremamente
ricche in biodiversità
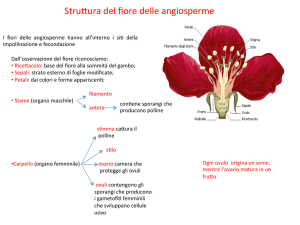
Caratteri distintivi di questa sottoclasse sono:
Legno con trachee; Fiori (equivalenti di rami ad accrescimento definito) con foglie
modificate sterili volte a formare un perianzio (sepali, petali o tepali) e sporofilli
specializzati (stami e carpelli)
Produzione di semi in una struttura (frutto) con funzioni protettive e di facilitazione della
diffusione
Doppia fecondazione e estrema riduzione dei gametofiti: quelli maschili, contenuti nel
granulo pollinico, a maturità sono costituiti da 3 cellule; i femminili, contenuti nell'ovulo,
a maturità sono costituiti generalmente da 7 cellule (8 nuclei).
L'impollinazione è operata in molti casi da animali. La forma del fiore in ogni caso è un
adattamento per favorire l'impollinazione. Tipi:
Fiori zoofili (ca. 88%): fiori di forme e colori variabili, che esprimono spesso il rapporto
di coevoluzione tra piante e animali (v. tabella di riepilogo).
Angraecum sesquipedale (Orchidaceae)
"It is, however, surprising that any insect
should be able to reach the nectar: our English
sphinxes have probosces as long as their
bodies; but in Madagascar there must be
moths with probosces capable of extension to
a length of between ten and eleven inches!"
(Darwin 1862).
Xanthopan morganii subsp. praedicta
Echinocereus sp. (Cactaceae)
Caltha palustris (Ranunculaceae)
Ophrys sp. (Orchidaceae)
Stapelia sp. (Asclepiadaceae)
Rosmarinus officinalis (Lamiaceae)
nettarii
Zigadenus sp.
(Melanthiaceae)
Stenocereus thurberi
(Cactaceae)
Strelitzia reginae (Strelitziaceae)
Serapias (Orchidaceae)
rifugio
Disa ferruginea (Orchidacea) Bellevalia flexuosa (Asparagaceae)
non nettarifera
nettarifera
Mimetismo fiorale
Fiori anemofili: grande quantità di polline, efficienti sistemi di cattura del polline
aerodiffuso (v. noce). È un adattamento tipico delle regioni temperate, insolito nei tropici
noce: Juglans nigra L.
Agropyron sp.
(Poaceae)
Zea mays (infiorescenze maschili e
femminili) - Poaceae
amenti maschili di betulla
(Betulaceae)
Fiori idrofili: in poche specie (circa 150; es. Vallisneria).
Anche la dispersione di semi o frutti può essere legata al vento (anemocoria), all’acqua
(idrocoria) o ad animali (zoocoria)
Harpagophytum
(Pedaliaceae)
Xanthium
(Asteraceae)
Talora i fiori sono riuniti in infiorescenze che possono essere definite (dette anche
cimose) o indefinite (dette anche racemose)
Ciclo di una angiosperma: Soia (Glycine max)
Gli adattamenti per il trasporto di polline da una pianta all'altra sono volti a favorire
l'impollinazione incrociata, che si basa su diverse strategie:
separazione di fiori staminiferi e fiori pistilliferi su piante diverse (piante dioiche: es.
Laurus nobilis, alloro)
produzione sulla stessa pianta di fiori unisessuali (piante monoiche: es. generi
Quercus, Betula, Alnus), spesso a maturazione differenziata.
Vi sono anche condizioni miste: (andromonoiche, ginomonoiche, androdioiche e
ginodioiche)
separazione temporale della produzione di stami e pistilli nei fiori ermafroditi
(dicogamia)
separazione spaziale di stami e stimmi in fiori ermafroditi (ercogamia/eterostilia)
autoincompatibilità genetica gametofitica o sporofitica.
Sistemi sessuali presenti nelle piante e loro effetti
sui tassi di autogamia in piante autocompatibili
Ermafroditismo (79%)
Monoicismo (5%)
1%
10%
Andromonoicismo
Androdioicismo
Ginomonoicismo
Ginodioicismo
Trioicismo
Dioicismo (5%)
(Autogamia elevata)
Autogamia
modulata a
seconda
dell’abbondanza
dei diversi tipi
sessuali nelle
popolazioni
(Autogamia nulla)
Il Dioicismo nelle
angiosperme si sarebbe
originato più volte e
indipendentemente
La maggior parte delle
angiosperme (ca. 80%)
sono ermafrodite ma nel
75% delle famiglie sono
presenti specie monoiche
e/o dioiche
L’evoluzione di cromosomi
sessuali nelle piante è
recente (ca.10-20 milioni
anni) rispetto ai
mammiferi (300 milioni
anni) in cui è molto più
frequente trovare
eteromorfismo
Lo studio dei cromosomi sessuali…
•
•
•
H. v. Henking (1891, scoperta dell’ereditarietà del sesso su base cromosomica)
McClung e Sutton (1901) individuarono il “cromosoma accessorio” (X)
E. Baur (1912) descrisse un mutante a foglie strette di Silene alba, la cui morfologia
era legata al sesso.
Negli anni successivi (anni ’40-’90) studi di citogenetica hanno permesso di evidenziare
la presenza ed eventuale eteromorfismo dei cromosomi sessuali (es. Polygonaceae,
Cannabidaceae)
Solo negli ultimi 20 anni con tecniche di genomica e biologia molecolare identificazione
dei geni “sessuali”, al loro sequenziamento e a quello dei cromosomi:
• Zea mays 1993, clonazione dei geni
• Silene 1999
• Asparagus 2000, mappatura cromosomi sessuali
• Humulus lupulus 2000
• Carica papaya 2004
S. latifolia
R.acetosa
Westergaard nel 1958, raggruppò i cromosomi sessuali delle piante in 3 tipi che illustrano
3 successivi stadi evolutivi:
• Vitalità genotipo YY (l’Y differisce dall’X solo per i geni “sessuali”) es. Ecballium,
Asparagus
• Il genotipo YY non è vitale ma il cromosoma Y ha un ruolo decisivo per la
determinazione del sesso. Es. Carica papaya, Silene latifolia
• Il cromosoma Y diventa irrilevante per la determinazione del sesso che è determinato
dal bilanciamento X:Autosomi. (>1 femmina, <0.5 maschio)
Es. Rumex acetosa, Humulus japonicus
Le tappe principali verso la formazione dei cromosomi sessuali:
1. Comparsa mutazioni maschio/femmina sterilità su autosomi e soppressione
della ricombinazione nella regione dei loci e immediate vicinanze
Su f =suppressor female
M = anther maturation, stamen promoting
La presenza di più geni e vicini tra loro serve a prevenire la ricombinazione tra X e Y
che potrebbe portare a individui “neutri”
La vicinanza di tali geni sarebbe frutto di inversioni e/o traslocazioni
Quando la degenerazione dell’Y è allo stadio iniziale, il genotipo YY è vitale (Active-Y
chromosome System)
Es. Asparagus officinalis
2n=20
Le 10 coppie sono ben distinguibili per taglia
e misura
La coppia 5 è stata identificata come quella
dei cromosomi sessuali, omomorfici con
basso contenuto di eterocromatina
costitutiva.
Il cromosomi sessuali contiengono 2 geni
vicini tra loro: M (male acivator) e F (female
suppressor)
Il sesso maschile è in eterozigosi
2. Degenerazione dell’ Y
La soppressione della ricombinazione si estende e porta alla degenerazione di una piccola
porzione di cromosoma e alla formazione di una MSY (Male Specific-Y region)
Le prime cause di degenerazione:
Muller’s ratchet, accumulo di mutazioni deleterie in modo irreversibile
Hitch-hiking, fissazione di mutazioni alleliche deleterie in quanto unite a forme alleliche
vantaggiose
Background selection, cioè selezione contro le mutazioni deleterie attraverso un tasso di
mutazioni ricorrente (ostacola la fissazione delle mutazioni alleliche mediamente
vantaggiose)
La regione MSY, contentente geni essenziali, può essere protetta dalla degenerazione con
la duplicazione (es. la MSY dell’uomo ha 9 strutture palindrome derivate dalla
duplicazione)
La perdita di contenuto genico dovuta alla degenerazione è sufficiente per causare la
letalità del genotipo YY
Es. Carica papaya
Trioica
Nel 1938 si pensava che il sesso fosse
determinato da un unico gene con 3 alleli
Nel 1953, Storey propose un gruppo di geni vicini
tra loro
Poiché i genotipi omozigoti dominanti non sono
vitali, Horovitz e Jimenez (1967) intuirono che la
determinazione del sesso fosse del tipo XX-XY
La porzione non ricombinante occupa il 10-15% del cromosoma e tra X e Y il tasso di
ricombinazione è diverso
Dal sequenziamento di cloni batterici: MSY ha minore densità di geni e maggiore di
elementi trasponibili
L’accumulo di elementi trasponibili e le duplicazioni nella regione MSY causano
aumento della quantità di DNA nell’Y
La regione degenerata si estende e X e Y diventano eteromorfici, l’Y è più grande
(es. Silene)
3.
4.
La forte degenerazione dell’Y causa la perdita di funzione di molti geni e la successiva
eliminazione di queste porzioni: l’Y si riduce di taglia (es. mammiferi)
5. L’Y viene perso e il sesso è determinato dalla proporzione X:Autosomi
(es.Rumex, Humulus). La soppressione della ricombinazione si estende all’intero Y,
perdita anche della regione pseudo-autosomica
Vitalità YY
XX-XY
X:A ratio
I casi studio più importanti…
Es. gen. Silene
Tutte le specie 2n=24
La maggior parte sono ginodioiche (S.noctiflora, S.vulgaris)
Poche sono ermafrodite (S.conica, S.gallica) o dioiche (S.latifolia, S.dioica)
S.dioica
S.gallica
N° corretto di cromosomi - Strasburger 1910
Presenza di cromosomi sessuali Blackburn
(1923) e Winge (1923)
Solo nel 1948 Westergaard capì la funzione
dell’Y
E solo 50 anni dopo che l’X è indispensabile per
l’embriogenesi (Veuskens, 1992) e lo sviluppo
del gametofito femminile (Janousek, 1998)
S.noctiflora
Es. gen. Rumex
R. acetosella
Presenta una certa variabilità nel N° cromosomico
Comprende specie ermafrodite, poligame (trioiche),
ginodioiche e dioiche
Il dioecismo si è originato una sola volta nel genere ed
esistono due lineages monofiletici:
• XX/XY (R.acetosella, tetraploide; R.graminifolius e
R.suffruticosus-endemico delle montagne centro
settentrionali della penisola iberica)
•XX/XY1Y2 e X:A (R.acetosa, diploide; R.papillaris o
intermedius)
I due Y sono quasi eterocromatici con 2 famiglie di DNAsatelliti (RAE180 e RAYSI)
R. acetosa
R. suffruticosus e R. acetosella hanno cromosomi sessuali meno
differenziati di quelli di R. acetosa, e che sono più simili allo stadio
evolutivo di quelli del genere Silene
Sebbene il dioicismo sia comparso in tempi simili nei generi Rumex e
Silene (circa 20 milioni anni fa), questo ha accumulato una quantità
molto inferiore di sequenze di DNA ripetitive nell’Y.
Altri meccanismi fiorali che influenzano i
livelli di autogamia nelle piante
Dicogamia (proterandria/proteroginia)
Ercogamia
Eterostilia
Autoincompatibilità
Incapacità fisiologica nei confronti
dell’autofecondazione: il polline di un individuo è
riconosciuto dagli stimmi dei fiori portati dallo stesso
individuo e gli viene impedito di germinare o,
comunque, di fecondare la cellula uovo.
Alcuni cenni storici…
C. Darwin (1877)
“The Different Forms of Flowers on Plants of the same Species”
Lloyd (1975) “The manteinance of gynodioecy and androdioecy in
angiosperms”
Charlesworth (1984) “Androdioecy and evolution of dioecy” - Rewiev
• osservazione patterns di allocazione delle risorse
• applicazione modelli matematici
(J.R. Pannell, 2002)
La variabilità nei sistemi sessuali
presenza di uno o più fiori per individuo
“sesso” morfologico e/o funzionale
ercogamia (separazione fisica delle
strutture sessuali nel fiore)
dicogamia (separazione temporale delle
funzioni sessuali: proteroginia e proterandria)
difasia (alternanza dell’espressione
sessuale dell’individuo nella stagione)
Cosa determina il sesso nelle piante?
• espressione genica
• self-crossing / out-crossing
• allocazione ottimale delle risorse verso le funzioni maschile e femminile
(S. C. H. Barret, 2002)
Verso il polimorfismo sessuale
In una popolazione ermafrodita…
Le strategie di ogni “sesso” sono caratterizzate dal relativo contributo alla fitness che
risulta come investimento paterno e/o materno per le generazioni successive.
Verso il polimorfismo sessuale
Buona impollinazione
Favorisce la diffusione di Femmine
Disponibilità di risorse
(Ginodioicismo/monoicismo)
Scarsa impollinazione
Favorisce la diffusione di Maschi
Risorse limitate
(Androdioicismo/monoicismo)
Limiti per la fitness:
• Maschile
disponibilità di ovuli, dispersione polline
• Femminile
risorse energetiche e ambientali
La mancanza delle strutture femminili consente una completa
allocazione delle risorse verso quelle maschili (meno costose)
I maschi possono raddoppiare la loro fitness:
quantità polline
vitalità polline
(J. R. Pannell, 2002)
Androdioicismo, fenomeno raro
maschi
Ginodioicismo: risposta
alla depressione da
inbreeding
Androdioicismo: nelle
popolazioni out-crossing e
solo dove le condizioni
ecologiche permettono un
grande aumento della
fitness maschile
femmine
(J. R. Pannell, 2002)
Casi particolari:
Dioicismo criptico (Cryptic dioecy)
Piante con sistemi riproduttivi apparentemente misti, si rivelano funzionalmente dioiche
ad una più attenta analisi
Difasia di genere (Gender diphasy)
Piante che variano la loro espressione sessuale a seconda dell’età e/o della taglia degli
individui
Alti tassi autogamici sono vantaggiosi
soprattutto
in
popolazioni
piccole
e
frammentate poiché facilitano la produzione di
semi e quindi il recupero di nuovi individui.
In popolazioni di grandi dimensioni, che hanno
buone chances di riproduzione incrociata,
l’autogamia diventa svantaggiosa in virtù
dell’inbreeding depression da essa indotta
Effetto Allee
Aumento dell’efficienza riproduttiva registrato in
piccole popolazioni animali al crescere del numero
di individui in esse presenti (Allee et al., 1949;
Allee, 1951).
L’effetto Allee è stato rinvenuto anche nelle piante:
-in quelle entomofile una elevata densità di
individui favorisce maggiore abbondanza di pronubi
e, quindi, una più elevata produzione di semi.
-in quelle anemofile, le probabilità di impollinazione
crescono in popolazioni molto ricche.
La legge di Baker
Sebbene l’incrocio e l’alta variabilità genetica da esso
indotta nella progenie siano spesso adattativamente
favorevoli, in alcune circostanze l’affidabile e costante
produzione di semi, e l’omogeneità genetica determinate
dall’autogamia possono risultare più appropriate al
contesto ecologico in cui le piante vivono (Baker, 1966)
In condizioni ambientali assai sfavorevoli ad una efficiente
impollinazione, la selezione naturale tende a favorire
caratteri fiorali che le chances di autofecondazione,
soprattutto in piante annue
Sistemi riproduttivi misti
Tra gli estremi rappresentati dalle piante abitualmente incapaci di
autofecondazione, e quelle che si riproducono principalmente per
autofecondazione, può essere posizionata la maggior parte delle angiosperme,
in cui sono presenti vari tassi di incrocio/inincrocio (tradotto da Baker, 1959)
La riproduzione mista
Selfers
obbligati
Out-breeders
obbligati
Piante con sistemi
misti
Piante principalmente
autogame
Piante con sistema misto
(da Vogler & Kalisz, 2001)
20%
33%
Ruolo della riproduzione mista nelle piante
L’autofecondazione permette la riproduzione
quando le possibilità d’incrocio sono nulle
(Darwin, 1876; Baker, 1955)
Tassi d’impollinazione variabili durante una
stessa stagione di fioritura, o tra anni
differenti, favoriscono anch’essi una certa
frequenza di autofecondazione, conducendo
ad una commistione di riproduzione incrociata
ed autogama
Al diminuire della frequenza delle visite degli
impollinatori
aumenta
il
contributo
dell’autofecondazione alla produzione di semi
della popolazione
Meccanismi d’autoimpollinazione (1)
Cleistogamia (es. in Viola): vantaggi energici legati al risparmio sulla
fase maschile ed impollinazione
Geitonogamia: fonde le proprietà ecologiche dell’impollinazione
incrociata e le conseguenze genetiche dell’auto-fecondazione
(da Lloyd & Schoen, 1992)
Meccanismi d’autoimpollinazione (2)
Autoimpollinazione
facilitata
(il
movimento
dell’impollinatore nel fiore determina il trasferimento
di polline dalle antere agli stimmi): per eliminarla
sono necessari arrangiamenti delle strutture fiorali
che fanno si che il contatto con lo stimma preceda
quello con le antere (es. Cypripedium)
Autoimpollinazione autonoma: non
intervento di vettori pollinici o visitatori
(da Lloyd & Schoen, 1992)
richiede
Autoimpollinazione autonoma
Anticipata: avviene prima che si creino condizioni
adatte all’impollinazione incrociata (per es. fiori che
diventano ercogami in un secondo tempo, sono
necessari ulteriori studi)
Competitiva: avviene in un contesto che è
potenzialmente idoneo anche all’impollinazione
incrociata (caratterizza fiori ermafroditi privi di
meccanismi di separazione spazio-temporali tra
organi maschili e femminili)
Ritardata: si verifica successivamente ad un
periodo in cui è favorita l’impollinazione incrociata
(fiori dicogami o solo inizialmente ercogami)
(da Lloyd & Schoen, 1992)
Il meccanismo migliore per ottimizzare i vantaggi offerti dal mescolare impollinazione
incrociata ed autoimpollinazione è rappresentato dall’autoimpollinazione ritardata;
questa dando precedenza temporale all’incrocio consente infatti di:
minimizzare il dispendio di ovuli e polline resi indisponibili alla riproduzione
incrociata
assorbire i massimi livelli di inbreeding depression
Per contrasto, molte angiosperme delle regioni temperate si moltiplicano per
propagazione vegetativa o apomissia. Ciò accade di norma nelle specie poliploidi.
Sottoclasse Magnoliidae (angiosperme): origine
Nel 1879, in una lettera al grande botanico Joseph Hooker, Charles Darwin definì
l’evoluzione del fiore, e quindi l’origine delle angiosperme, come un “abominable
mystery”
Nel tempo, svariate teorie sono state proposte per spiegare l’”improvvisa” comparsa,
alla fine del Mesozoico (nel Cretaceo, fra i 120 ed i 95 milioni di anni fa), di questo
gruppo. Ne ricordiamo qui alcune:
- Teoria pseudàntica (Eichler, 1876 – Engler 1897): le angiosperme sarebbero derivate
dalle conifere (Pinales) o dalle Gnetales. Questi autori ritenevano fondamentalmente
omologhi i coni riproduttivi di questi gruppi ed i fiori delle angiosperme. Una modifica di
questa teoria (da molti sostenuta ancora negli anni ’90) prevedeva che i progenitori delle
angiosperme fossero le Gnetales, o che almeno queste condividessero un antenato
comune. Le Casuarinaceae erano viste come la famiglia più primitiva tra le
angiosperme viventi in virtù delle loro infiorescenze femminili organizzate in
“coni” – In realtà si tratta di angiosperme evolute (Fagales)
- Teoria euàntica (Delpino, 1890 – Bessey 1893): le angiosperme sarebbero derivate
da un particolare gruppo estinto di gimnosperme (o da un progenitore comune con), le
Cycadeoidales, caratterizzate da particolari strutture riproduttive che potevano ricordare
i fiori delle angiosperme. Le Magnoliales ed altri gruppi affini erano viste come i
gruppi più primitivi esistenti.
altri studiosi, nel tempo, hanno sostenuto una origine (anche polifiletica!) delle
angiosperme da più gruppi di gimnosperme o di pteridosperme come le paleozoiche
Glossopteridales e le mesozoiche Caytoniales.
Più recentemente, Stuessy (2004), tenendo conto di tutte le informazioni disponibili
provenienti sia dalla sistematica molecolare che dalla paleontologia, ha elaborato una
teoria (definita “Teoria della transizione-combinazione”) che sembra in effetti essere
quella più fondata e sostenuta dai dati:
Le angiosperme si sarebbero evolute lentamente tra quelle pteridosperme del
Giurassico (ca. 150 milioni di anni fa) che avevano evoluto dei carpelli.
Successivamente, in questa linea filogenetica sarebbe comparsa la doppia
fecondazione così come appare oggi nelle angiosperme. Infine, nell’ambito di questa
nuovo gruppo, sarebbero comparsi i primi fiori (con perianzio). Queste 3 transizioni
fondamentali non si sarebbero combinate assieme per almeno 100 milioni di anni.
Quando ciò è avvenuto, si è avuta la grossa radiazione evolutiva delle angiosperme, in
risposta soprattutto alla selezione dovuta alla coevoluzione con gli insetti ed altri animali
(sia predatori che agenti dispersori dei semi).
Questa teoria spiegherebbe anche la discrepanza tra l’età dell’origine indicata dai dati
molecolari (200 milioni di anni fa e anche più), che testimonierebbe l’origine del gruppo
pteridospermico carpellato (forse Caytoniales?) che ha poi evoluto la doppia
fecondazione e succesivamente la struttura fiorale completa, e l’età indicata dai primi
reperti fossili (Cretaceo, 100-130 milioni di anni fa).
Da ciò discende che le gimnosperme esistenti non avrebbero connessioni filogenetiche
dirette con le angiosperme, e ciò e confermato da recenti studi molecolari che vedono le
esistenti Pinopsida come sister group delle Magnoliopsida, evolutosi in parallelo ad
esse.
Archaefructus (125 milioni di anni
fa)
Interpretato come
protoangiosperma con “fiori”
pluricarpellari, ma senza perianzio.
Secondo alcuni autori si tratterebbe
di infiorescenze femminili con ogni
fiore ridotto ad un singolo carpello,
che avrebbero perso
secondariamente il perianzio (come
accade nelle viventi
Zannichelliaceae).
I primi granuli pollinici di
angiosperme (Clavipollinites),
monoaperturati, risalgono più o
meno allo stesso periodo.
Archaeanthus (100-95 milioni di
anni fa)
una delle più antiche angiosperme
note allo stato fossile, simile alle
attuali magnolie.
Dello stesso periodo sono noti
anche molti fossili di “paleoerbe”,
alcune delle quali addirittura
ascrivibili a famiglie tuttora esistenti
(es. Chloranthaceae, Piperaceae,
Nymphaeaceae etc.)
Leefructus (120-125 milioni di anni
fa)
una delle più antiche angiosperme
note allo stato fossile… attribuita
dagli autori alle eudicots
(Ranunculales)!!
Rapporti filogenetici tra le angiosperme
viventi
All’inizio degli anni 90’ si è formato un gruppo
di studiosi con lo scopo di chiarire con
moderne tecniche di sistematica molecolare, i
rapporti filogenetici tra le angiosperme:
l’Angiosperm Phylogeny Group (APG).
Questo gruppo ha pubblicato i primi risultati
nel 1998 (APG: 462 famiglie, raggruppate in
40 ordini), aggiornati poi nel 2003 a 457
famiglie riconosciute e 45 ordini (APG II) e nel
2009 a 413 famiglie e 62 ordini (APG III).
ANGIOSPERM PHYLOGENY WEBSITE
http://www.mobot.org/MOBOT/research/AP
web/welcome.html
La tradizionale suddivisione delle angiosperme in monocotiledoni e dicotiledoni non è
sostenuta da tali analisi
le dicotiledoni appaiono infatti un gruppo parafiletico con caratteri plesiomorfici quali i
due cotiledoni, la radichetta persistente, fusti con fasci vascolari ad anello,
accrescimento secondario e foglie retinervie
l'origine monofiletica delle monocotiledoni (“monocots”, per noi superordine Liliianae) è
indicata da caratteri sinapomorfici: foglie parallelinervie, embrione con un solo
cotiledone, fusti con fasci vascolari sparsi (atactostele) e sistema radicale avventizio
tra le dicotiledoni, un notevole numero di specie costituiscono un gruppo monofiletico
(“eudicots”, svariati superordini di “vere dicotiledoni”), con sinapomorfie che includono
polline triaperturato (tutte le altre angiosperme, monocotiledoni incluse, hanno polline
mono-aperturato), fiori stereotipati su base 4-5 (le dicotiledoni che fanno eccezione sono
quasi tutte al di fuori di questo clado) alcune particolari sequenze nucleotidiche
Superordine Lilianae
fitomelano nei semi;
tegumento seminale
esterno collassato,
nettarii nei setti
dell’ovario
no fitomelano;
tegumento seminale
esterno cellulare;
nettarii alla base dei
tepali
Acorus
Alisma
Tamus
Commelina
Chamaerops
Eudicots
Ceratophyllum
Superordine Rosanae
clado
corrispondente
grossomodo alle
“dicotiledoni
gamopetale”
Superordine Asteranae
polline tri-aperturato
Tipi di aperture (zone dove la
sporopollenina che riveste il granulo
pollinico è più sottile per facilitare
l’emissione del tubetto pollinico):
-porato: con delle aperture rotonde
-colpato: con delle aperture
longitudinali
-colporato: che presenta entrambe le
caratteristiche
ciò vale per tutti i tipi di polline
le specie rimanenti di dicotiledoni – con polline monoaperturato – sono un gruppo
estremamente eterogeneo e parafiletico. Molte hanno olii eterei e elementi vascolari
allungati con placche inclinate e perforazioni scalariformi; fiori attinomorfi, con carpelli e
stami prevalentemente liberi; semi con embrione piccolo e endosperma abbondante
i componenti legnosi di questo gruppo sono indicati come magnoliide, quelli erbacei
come paleoerbe
le magnoliide (Amborellales, Austrobaileyales, Canellales, Magnoliales, Laurales) sono
piante a portamento arboreo/arbustivo con foglie alterne o opposte, coriacee, con
nervatura pennata e stomi paracitici (si distinguono cellule ausiliarie). Fiori composti da
parti separate tra loro, pezzi del perianzio disposti a spirale o in verticilli di tre e stami
spesso laminari.
le paleoerbe (Nymphaeales, Piperales, Chloranthales, Ceratophyllales) sono erbe di
piccole dimensioni, con foglie alterne, sottili, a nervatura più o meno palmata e stomi
anomocitici (non si distinguono cellule ausiliarie). Fiori composti da numerose o poche
parti, quelle dell'androceo e del perianzio generalmente in verticilli di tre (tutti caratteri
che le avvicinamo molto alle monocotiledoni, anch’esse considerabili paleoerbe).
polline mono-aperturato
adesso tutte
riunite
nell’Ordine
Austrobaileyales
Nymphaea sp.
A. trichopoda: piccolo suffrutice dioico con fiori piccoli,
endemico della Nuova Caledonia (Australia)
“ANITA” grade
Friedman dopo aver scoperto
nel 2002 (NATURE) che nelle
Austrobaileyales e
Nymphaeales (Illicium,
Nuphar etc.) il gametofito è 4nucleato (2 sole divisioni
della somatogenesi: tipo
Schisandra) e quindi
l’endosperma secondario che
si forma in queste piante in
seguito alla doppia
fecondazione è diploide,
ipotizza nel 2004 questa
teoria modulare dello
sviluppo del gametofito
femminile nelle angiosperme
Ciò può valere per le
angiosperme evolutesi
successivamente, ma non
per Amborella trichopoda!
Infatti questa pianta è stata
recentemente re-investigata
dallo stesso Friedman.....
che ha scoperto (pubblicato
su NATURE nel Maggio
2006) che A. trichopoda non
presenta affatto un normale
sviluppo di tipo Polygonum
come riportato in letteratura,
ma a questo si aggiunge,
dopo la formazione del
gametofito, 1 divisione della
gametogenesi che va a
generare l’oosfera da una
delle tre cellule somatiche
micropilari!
ANELLO DI
CONGIUNZIONE CON LE
GIMNOSPERME
Probabilmente, questa è la condizione di sviluppo più primitiva, alla quale sarebbe
seguita una drastica riduzione del gametofito femminile a 4 cellule (ancora presente
nelle Austrobaileyales), che nelle angiosperme originatesi successivamente si sarebbe
“raddoppiato” nel modo visto in precedenza.
Gametofito femminile di
tipo Schisandra
Formazione di
perisperma (=
endosperma primario)
PRIMA della
fertilizzazione!!
Altre Nymphaeales (ed
Austrobaileyales)
producono comunque
perisperma, ma dopo la
fertilizzazione.
Magnolia
Laurus
Chloranthus
Canella
Superordine Magnolianae
Aristolochia
Piper
fiore di Magnolia grandiflora
strutture che, nelle piante moderne, possono essere ritenute plesiomorfiche:
- granulo pollinico monoaperturato
- perianzio con scarsa differenziazione tra calice e corolla
- sepali e petali liberi
- carpelli non specializzati, in particolare privi di superfici stimmatiche, e separati. In
alcuni gruppi basali i carpelli non sono completamente chiusi
4 tendenze evolutive nei fiori:
- da fiori con pezzi in numero indefinito a fiori con pochi pezzi in numero definito
- riduzione del numero di verticilli fiorali da 4 a 3, 2 o 1, con estremo accorciamento
dell'asse fiorale
- ovario da supero a infero
- da simmetria radiale a simmetria bilaterale
di gran
lunga il più
diffuso
CASI ESEMPLIFICATIVI DELLA
VARIABILITA’ DI COSTITUZIONE DEL
GAMETOFITO FEMMINILE DALLE
GIMNOSPERME ALLE
ANGIOSPERME