LIPIDI
e
METABOLISMO
DEGLI ACIDI GRASSI
Lipidi
• I lipidi sono i principali costituenti delle membrane
biologiche.
• Sono biomolecole insolubile in acqua ma solubili in solventi
organici.
• La loro idrofobicità è dovuta alla presenza degli acidi grassi
nelle molecole di lipidi.
• I lipidi svolgono molteplici ruoli biologici:
•
•
•
•
Molecole combustibili
Molecole segnale
Riserva di energia altamente concentrata
Componenti delle membrane
Gli acidi grassi
Gli acidi grassi sono acidi carbossilici con catene idrocarburiche di
varia lunghezza e grado di insaturazione.
• Gli acidi grassi a 16 e 18 atomi di C sono i più comuni, la catena
idrocarburica è quasi sempre non ramificata e può essere satura o
può contenere uno e più doppi legami. I doppi legami nella maggior
parte degli acidi grassi sono cis. Negli acidi grassi poliinsaturi i doppi
legami sono separati da almeno un gruppo metilenico.
Acidi grassi
Acidi grassi definiti omega ( )
• Acidi grassi essenziali presenti in vari tipi di
alimenti (omega 3: grasso di pesce, salmone,
sgombri, acciughe ecc. nell'olio di pesce;
omega 6: noci e oli di soia, di mais, cibi
proteici, verdure e cereali) e convertiti in vivo
in altri acidi grassi poliinsaturi o altri tipi di
lipidi
Fluidità degli acidi grassi
Più la catena
idrocarburica è corta
e più il grado di
insaturazione è alto,
maggiore è la fluidità
degli acidi grassi e dei
loro derivati
I triacilgliceroli o grassi
• Nel nostro organismo gli acidi grassi sono
immagazzinati negli adipociti (tessuto
adiposo) sotto forma di triacilgliceroli.
•
•
•
•
Svolgono tre specifiche funzioni:
Produzione di energia
Produzione di calore
Isolamento termico
• Sono triesteri degli acidi grassi e del
glicerolo:
H2C-O-CO-R1
H-C-O-CO-R2
H2C-O-CO-R3
SAPONIFICAZIONE:
glicerolo
I LIPIDI DI MEMBRANA
• I lipidi di membrana sono molecole
anfipatiche contenenti una porzione idrofilica
(testa polare) e una porzione idrofobica (coda
apolare).
Testa polare
• I principali lipidi di membrana sono:
Fosfolipidi
Glicolipidi
Colesterolo
Coda idrofobica
I fosfolipidi (fosfogliceridi e sfingolipidi)
•
I fosfolipidi sono la principale classe di lipidi di membrana
•
Sono costituiti da 4 tipi di molecole: acidi grassi, una molecola a cui
sono legati gli acidi grassi, un fosfato a cui è legato ad un alcool.
•
Gli acidi grassi costituiscono la porzione idrofobica del lipide mentre
il resto della molecola ha proprietà idrofiliche per permettere
l’interazione con l’ambiente circostante.
•
La molecola a cui sono legate le varie parti del lipide può essere il
glicerolo (fosfogliceridi o glicerofosfolipidi) o la sfingosina
(sfingolipidi).
•
Gli alcoli presenti nei fosfogliceridi sono: serina, etanolammina,
colina, glicerolo e inositolo.
•
La sfingomielina è uno sfingolipide presente nelle membrane del
tessuto nervoso.
Ac. grasso
glicerolo
Ac. grasso
fosfato
alcool
Principali fosfolipidi
I glicolipidi (glicosfingolipidi e galattolipidi)
•
•
•
•
Sono lipidi che contengono carboidrati.
La struttura contiene sfingosina o glicerolo legata ad acidi grassi ed a
residui di carboidrati.
Il glicolipide più semplice, chiamato cerebroside, lega una sola molecola
di zucchero (glucosio o galattosio)
I galattolipidi contengono un gruppo solfato
• I gangliosidi sono glicolipidi più
complessi che contengono una catena
ramificata di residui saccaridici.
•Nelle membrane i glicolipidi sono
orientati in modo asimmetrico: i
residui di carboidrati sono sempre
esposti sul versante extracellulare
Il colesterolo
• E’ un lipide con un nucleo steroideo a
cui sono legati una catena
idrocarburica e un gruppo ossidrilico.
• Nelle membrane la molecola è
orientata parallelamente alla catena
degli acidi grassi dei fosfolipidi.
• Nei neuroni costituisce il 25% dei
lipidi di membrana.
Principali classi di lipidi
di riserva e di membrana
LE MEMBRANE CELLULARI
•
La formazione delle membrane è una
conseguenza della natura anfipatica
delle molecole lipidiche che sono la
componente maggiore.
•
I lipidi formano un doppio strato di
molecole: le code idrocarburiche si
impaccano nella porzione interna
(interazioni di van der Waals e
idrofobiche), le teste polari
interagiscono con l’ambiente acquoso
cellulare ed extracellulare
(attrazioni elettrostatiche e legami
a idrogeno)
•
% dei diversi tipi di membrana nell’
epatocita di ratto
Membr. Plasm.
RER+REL
Mitocondri
Golgi
Altre
5%
45 %
40 %
5%
5%
Metabolismo dei lipidi:
-ossidazione
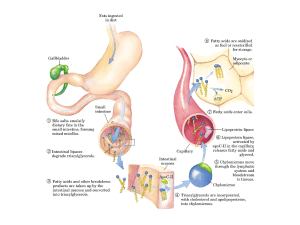
Mobilizzazione dei grassi di riserva
I triacilgliceroli possono essere mobilizzati dall’azione idrolitica di lipasi
soggette a regolazione ormonale.
Azione delle fosfolipasi sui fosfogliceridi
Esistono vari tipi di fosfolipasi (PLA2, PLA1, PLC, PLD) che idrolizzano i legami
esterei che legano i vari acidi grassi al glicerolo nei fosfolipidi. Ciascuna è specifica
per un determinato legame del fosfolipide.
Metabolismo degli
acidi grassi
L’ossidazione degli acidi grassi avviene in
tre fasi:
1) -ossidazione in cui gli acidi grassi vanno
incontro ad un distacco progressivo di unità
bicarboniose nella forma di acetil-CoA.
2) L’unità acetilica dell’acetil-CoA viene
ossidata a CO2 nel ciclo di Krebs
3) I coenzimi ridotti NADH e FADH2
donano i loro elettroni alla catena
respiratoria dei mitocondri, attraverso
questa via gli elettroni arrivano all’ossigeno
con la concomitante fosforilazione di ADP
ad ATP. L’energia rilasciata dall’ossidazione
degli acidi grassi viene quindi conservata
sotto forma di ATP.
Ossidazione degli acidi
grassi a catena pari
La via di
ossidazione degli
acidi grassi si
divide in due fasi:
- Attivazione e
trasporto nei
mitocondri degli
acidi grassi
-ossidazione
degli acil-CoA
Attivazione e trasporto
nei mitocondri degli acidi
grassi
• Il catabolismo degli acidi
grassi inizia nello spazio
intermembrana del
mitocondrio dove gli acidi
grassi sono attivati ad acilCoA utilizzando ATP
(2PAMP).
• La carnitina favorisce il
trasporto degli acil-CoA
nella matrice mitocondriale,
dove vengono ossidati gli
acidi grassi, sotto forma di
acil-carnitina.
Trasporto degli acil-CoA
-ossidazione degli acidi
grassi a catena pari
• Una volta all’interno della
matrice mitocondriale, gli acilCoA vengono degradati
mediante l’ossidazione iniziale
del carbonio in posizione e
attraverso una serie di
passaggi, ciascuno dei quali
rilascia un frammento a due
atomi di carbonio sotto forma
di acetil-CoA dall’acido grasso
che viene ossidato.
• La via metabolica è ciclica
poiché ogni passaggio termina
con la formazione di un acilCoA accorciato di due atomi di
carbonio che viene sottoposto
allo stesso processo nel ciclo
successivo.
Ciascun passaggio
comporta 4
reazioni:
1.
Deidrogenazione
con formazione
di un doppio
legame C=C
(deidrogenasi)
1.
Idratazione del
doppio legame
(idratasi)
1.
Deidrogenazione:
il gruppo
ossidrilico viene
ossidato a gruppo
carbonilico
(deidrogenasi)
1.
Scissione tiolica
con formazione
di acetil-CoA e
acil-CoA (tiolasi)
1.
2.
3.
4.
Ossidazione degli acidi grassi a
catena dispari e insaturi
Gli acidi grassi a numero dispari di atomi
di carbonio a seguito dell’ossidazione
danno una molecola di propionil-CoA (3C),
la cui conversione a succinil-CoA
comporta una reazione di carbossiliazione
e una reazione di riarrangiamento
catalizzate da enzimi che utilizzano come
coenzimi vitamine del complesso B.
L’ossidazione degli acidi grassi con doppi
legami richiede prima la conversione del
doppio legame da cis a trans e
successivamente la riduzione del doppio
legame a legame singolo.
Resa energetica dell’ossidazione
degli acidi grassi
• Ossidazione del palmitato (16 C):
Palmitoil-CoA + 7CoASH + 7FAD + 7NAD+ + 7H2O
→ 8 Acetil-CoA + 7FADH2 + 7NADH + 7H+
• Resa energetica in ATP:
Reazione
Attivazione del palmitato a palmitoil-CoA
Ossidazione di 8 molecole di acetil-CoA (ciclo di Krebs)
Resa in ATP
-2
8 x 10 = 80
Ossidazione di 7 molecole di FADH2
7 x 1.5 = 10.5
Ossidazione di 7 molecole di NADH
7 x 2.5 =17.5
Totale
106
Biosintesi
degli acidi grassi
Sintesi del malonil-CoA
• La sintesi inizia con la carbossilazione di acetil-CoA a
malonil-CoA; questa reazione è catalizzata dalla acetilCoA carbossilasi e consuma una molecola di ATP.
• Una molecola di malonil-CoA e una di acetil-CoA sono
ancorati alla proteina trasportatrice di acili (ACP) un
componente dell’enzima della biosintesi
Biosintesi
• Unità bicarboniose (2C) provenienti
dal malonil-CoA (3C) sono
progressivamente aggiunte alla
catena in formazione.
• Es. sette cicli di allungamento
producono il palmitoil-CoA che viene
idrolizzato a palmitato (acido grasso
a 16C) .
• Negli animali gli enzimi che
catalizzano la biosintesi degli acidi
grassi sono organizzati in un
complesso enzimatico
multifunzionale chiamato acido
grasso sintasi, localizzato nel
citosol. Gli intermedi che si formano
durante la sintesi sono ancorati al
complesso per mezzo della ACP.
• Gli acidi grassi insaturi vanno
incontro ad una reazione di
desaturazione per la formazione di
doppi legami.
Sintesi
Acido
grasso
sintasi
Consumo energetico della biosintesi
degli acidi grassi
La reazione complessiva della sintesi del palmitato
da acetil-CoA è:
Acetil-CoA + 7 malonil-CoA + 14 NADPH + 14 H+ →
→ palmitato + 7 CO2 + 8 CoA + 14 NADP+ + 6 H2O
Sintesi malonil-CoA:
Acetil-CoA + HCO3- + ATP → malonil-CoA + ADP + Pi
La sintesi di ciascuna molecola di malonil-CoA richiede una
molecola di ATP e la riduzione dei doppi legami durante la
sintesi dell’acido grasso utilizza NADPH.
Consumo: 7 ATP + 14 NADPH
Regolazione del metabolismo
degli acidi grassi
Regolazione dell’ossidazione degli acidi grassi
• La sintesi e la degradazione degli acidi grassi sono
regolate reciprocamente in modo da non essere
entrambe attivate nello stesso momento.
• L’ossidazione degli acidi grassi è regolata a livello
del processo che trasferisce l’acile nei
mitocondri.
• Il processo è rallentato da elevate concentrazioni
di malonil-CoA, primo intermedio della biosintesi
e indice di elevata disponibilità di glucosio.
• Il glucagone e l’adrenalina stimolano la
degradazione dei triacilgliceroli attivando la
lipasi, l’insulina inibisce la lipolisi.
Regolazione della sintesi degli
acidi grassi
• L’acetil-CoA carbossilasi (prima tappa
della biosintesi degli acidi grassi) è
stimolata dall’insulina e inibita dal
glucagone e dall’adrenalina.
• Il citrato stimola allostericamente la
carbossilasi.
CHETOGENESI:
formazione dei corpi chetonici
• Nello stato di digiuno e nello stato diabetico
l’ossalacetato viene consumato per la sintesi
di glucosio provocando un accumulo di acetilCoA.
• L’accumulo di acetil-CoA ad un livello
superiore a quello che può essere utilizzato
per l’ossidazione nel ciclo di Krebs e per la
sintesi degli acidi grassi, determina la sintesi
dei corpi chetonici:
ACETONE,
ACETOACETATO e D- IDROSSIBUTIRRATO.
• La via di formazione dei corpi chetonici è
chiamata chetogenesi e avviene nel fegato.
Vie biosintetiche dei corpi chetonici
L’acetoacetato si
forma per condensazione di due molecole
di acetil-CoA .
Dall’acetoacetato di
forma il
idrossibutirrato.
L’acetoacetato si
decarbossila
spontaneamente ad
acetone
Ruolo dei corpi chetonici
• I corpi chetonici vengono trasportati dal fegato ai tessuti
periferici.
• L’acetone prodotto in piccole quantità viene eliminato con la
respirazione.
• L’ acetoacetato e il D- -idrossibutirrato sono trasportati dal
sangue ai tessuti extraepatici dove vengono utilizzati per
produrre energia mediante l’ossidazione dell’acetil-CoA, ottenuto
dalla scissione dell’acetoacetato.
• Durante il digiuno o in condizioni patologiche (es. diabete), che
determinano una carenza di glucosio, alcuni tessuti (t. nervoso,
muscolare, corteccia surrenale..) utilizzano i corpi chetonici come
principale fonte di unità acetiliche.
• Se la quantità di corpi chetonici prodotta è superiore alla capacità
degli organi extraepatici di utilizzarli (es. digiuno prolungato o
diabete non controllato), l’aumento eccessivo di corpi chetonici nel
sangue (chetosi) provoca un abbassamento del pH generando una
condizione chiamata acidosi che può portare a coma e morte.
Metabolismo dei lipidi
Relazione tra sintesi degli acidi grassi e
metabolismo dei carboidrati
• L’acetil-CoA è un
intermedio chiave
tra il metabolismo
lipidico e il
metabolismo dei
carboidrati.
• I carboidrati
vengono facilmente
convertiti in grassi,
ma gli animali non
possono effettuare
la conversione degli
acidi grassi in
glucosio.
Quesiti
1)
2)
3)
4)
5)
6)
I Lipidi: definizione, classificazione e ruoli biologici
Quali sono i principali lipidi di membrana
Struttura e funzione dei trigliceridi
Attraverso quali fasi avviene l’ossidazione completa degli acidi grassi.
Quale è il ruolo della carnitina nel processo di ossidazione degli acidi grassi
Quali sono i meccanismi di regolazione del metabolismo degli acidi grassi, sia in
termini di ossidazione che di biosintesi.
7) In quali condizioni si può avere la formazione dei Corpi chetonici e dove avviene
8) Quali sono i corpi chetonici e quale è il prodotto di partenza per la loro formazione?
9) Quali tessuti/organi possono usare i corpi chetonici per produrre energia?
10) E’ possibile produrre acidi grassi a partire da glucosio? Può avvenire anche
l’opposto, ovvero è possibile convertire gli acidi grassi in glucosio: Motivare la
risposta.