Capitolo 27
METABOLISMO
DEI GRASSI
La maggior parte dell’energia conservata in un organismo si trova nei depositi di
grasso. In questi corpulenti combattenti di sumo la conservazione di energia
sotto forma di grassi è diventata un’arte.
Obiettivi del capitolo
• Essere capace di descrivere il meccanismo dell’ossidazione dei grassi
• Familiarizzare con la quantità di energia prodotta durante l’ossidazione dei grassi
• Essere consapevole del significato del ruolo dei corpi chetonici
• Essere informato su alcune malattie causate dall’alterazione
dei processi di ossidazione degli acidi grassi
• Seguire la via della lipogenesi e della formazione di colesterolo
440
Metabolismo dei grassi
27-1
Livello dei lipidi nel plasma
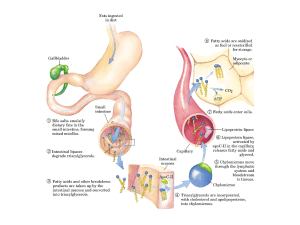
Durante la digestione, grassi e fosfolipidi vengono emulsionati e successivamente idrolizzati in acidi grassi e glicerolo. Essi vengono trasformati in trigliceridi all’interno della mucosa intestinale, passano nel dotto toracico e
quindi nel sangue; tuttavia tali sostanze sono insolubili in acqua e parimenti
nel sangue. Per poter essere trasportati dal sangue i grassi e i fosfolipidi formano un complesso con le proteine (idrosolubili) del plasma. Tali complessi sono chiamati lipoproteine. La Tabella 30.2 riporta i valori dei lipidi nel
sangue.
Anomalie del metabolismo lipidico portano a svariati tipi di ipolipoproteinemia o iperlipoproteinemia. Il tipo di anomalia più comune è il diabete,
in cui un difetto di insulina porta all’ipertriacilglicerolemia.
27-2
Assorbimento dei grassi
La digestione dei grassi ha inizio nell’intestino tenue, mediante idrolisi che
produce acidi grassi e glicerolo. Prima della digestione, i grassi vengono
emulsionati dai sali biliari. I prodotti della digestione dei grassi passano dai
*vasi chiliferi dei *villi ai *vasi linfatici dove essi appaiono come grassi
risintetizzati. Dai vasi linfatici i grassi fluiscono attraverso il dotto toracico
nel sangue e quindi giungono al fegato. Dopo un pasto, il contenuto in grassi del sangue aumenta e rimane per alcune ore ad alti livelli, poi gradualmente decresce fino ai livelli del digiuno.
Nel fegato alcuni dei grassi sono trasformati in fosfolipidi, in questo
modo il sangue, lasciando il fegato, contiene sia grassi che fosfolipidi. I
fosfolipidi, quali le sfingomieline e le lecitine, sono necessari per la formazione del tessuto nervoso e cerebrale. Le lecitine (fosfatidil coline) sono
inoltre coinvolte nel trasporto dei grassi ai tessuti. La cefalina, un altro
fosfolipide, è coinvolta nel normale processo di coagulazione del sangue.
Dal fegato, alcuni grassi si dirigono verso le cellule dove sono ossidati per
fornire calore ed energia. Il grasso in eccesso rispetto al fabbisogno cellulare è immagazzinato come tessuto adiposo.
La lipolisi, l’idrolisi dei triacilgliceroli (trigliceridi) in acidi grassi e glicerolo è controllata principalmente dalla quantità di cAMP presente nei tessuti. Gli ormoni che stimolano la produzione di cAMP e in questo modo incrementano la lipolisi comprendono epinefrina, norepinefrina, glucagone,
ormone adrenocorticotropo (ACTH), ormoni stimolanti i melanociti α e β
(MSH), ormone stimolante la tiroide (TSH), ormone della crescita (GH) e
vasopressina. Al contrario, l’insulina e le prostaglandine (vedi Paragrafo 229) abbassano i livelli del cAMP e in questo modo fanno diminuire la velocità
della lipolisi.
L’enzima che degrada il cAMP è inibito dalle metilxantine quali la caffeina e la teofillina. Un effetto insolito di questa inibizione è la presenza di un
netto e prolungato incremento dei livelli di acidi grassi liberi nell’organismo
di coloro che bevono grosse quantità di caffè (che contiene caffeina).
27-3
Ossidazione dei grassi
L’ossidazione dei grassi (trigliceridi), in realtà, comporta l’ossidazione di
due prodotti dell’idrolisi - glicerolo e acidi grassi. Gli aspetti generali del
441
Ossidazione dei grassi
Diretta
epinefrina
norepinefrina
ACTH
glucagone
attivazione della
adenilato ciclasi
Indiretta
ormone
della crescita
cortisolo
tiroxina
AMP ciclico
due stadi
attivazione della
trigliceride lipasi
glicerolo + 3 acidi grassi
trigliceride
Figura 27.1
Idrolisi di trigliceridi nel tessuto adiposo.
controllo della scissione dei trigliceridi in glicerolo e acidi gassi sono riassunti nella Figura 27.1. Questa sequenza è bloccata dall’insulina o da alti
livelli di glucosio.
Ossidazione del glicerolo
Il glicerolo che fa parte di un grasso è ossidato a diidrossiaceton fosfato,
come indicato nella sequenza successiva. Il diidrossiacetone fosfato fa parte
della via della glicolisi (vedi Paragrafo 26-6). Questo composto può essere
convertito in glicogeno nel fegato o nel tessuto muscolare o in acido piruvico che entra nel ciclo di Krebs. In questo modo il glicerolo che fa parte di
un grasso è metabolizzato attraverso la via dei carboidrati.
glicerochinasi
glicerofosfato
deidrogenasi
fosfatasi
glicerolo
α-glicerofosfato
diidrossiacetone
fosfato
Ossidazione degli acidi grassi
Vi sono varie teorie sull’ossidazione degli acidi grassi. La prima, proposta
da Knoop nel 1905 ed ancor oggi preferita, è chiamata teoria della β-ossidazione. Questa teoria prevede l’ossidazione del secondo atomo di carbonio
a partire dall’estremità carbossi terminale della molecola di acido grasso,
l’atomo di carbonio β. In questo processo la β-ossidazione rimuove due
atomi di carbonio per volta dalla catena dell’acido grasso. Ossia, un acido
grasso a 18 atomi di carbonio è ossidato ad acido grasso a 16 atomi di carbonio, quindi ad acido grasso a 14 atomi di carbonio e così via, fino a
quando il processo di ossidazione è completato. Una versione semplificata
di questa ossidazione è mostrata nella Figura 27.2.
442
Metabolismo dei grassi
O
||
---CH2—CH2—CH2—C—OH
acidi grassi
tiochinasi
acidi grassi attivi
acil CoA
deidrogenasi
Mg2+
enoil CoA
idratasi
acidi grassi trans-α, β-insaturi
acidi grassi L-β-idrossi
L-β-idrossiacil
acil CoA
deidrogenasi
Mg2+
CoA
deidrogenasi
β-chetotiolasi
CoA
acetil CoA
acidi grassi attivi
di due atomi di carbonio
β-chetoacil CoA
Figura 27.2
Ossidazione degli acidi grassi.
L’acetil CoA così prodotto entra nel ciclo di Krebs e la nuova molecola
di acido grasso attiva passa di nuovo attraverso la stessa sequenza, perdendo ogni volta due atomi di carbonio finché l’intera molecola di acido grasso
non è ossidata. Questa sequenza presuppone la presenza di acidi grassi
contenenti un numero pari di atomi di carbonio, condizione solitamente
riscontrata in natura. Il FADH2 ed il NADH + H+ entrano nella catena respiratoria. Se gli acidi grassi contenenti un numero dispari di atomi di carbonio
sono ossidati, essi seguono lo stesso processo, tranne il fatto che i prodotti
finali sono l’acetil CoA ed il propionil CoA. Il propionil CoA è trasformato
attraverso una serie di processi in succinil CoA che entra nel ciclo di Krebs,
così come l’acetil CoA. Queste reazioni richiedono la presenza di vitamina
B12 ed anche di biotina.
Gli acidi grassi insaturi sono metabolizzati lentamente. Devono prima
essere ridotti da alcune delle deidrogenasi presenti nelle cellule. Essi non
possono seguire per l’ossidazione il ciclo degli acidi grassi.
Energia prodotta dall’ossidazione degli acidi grassi
L’ossidazione di 1 g di grassi produce più del doppio dell’energia prodotta
dall’ossidazione di 1 g di carboidrati. Vediamo come ciò accade.
L’ossidazione dell’acetil CoA attraverso il ciclo di Krebs produce 12
legami fosforici ad elevata energia (ATP) per ogni molecola di acetil CoA.
Se si considera l’ossidazione dell’acido palmitico, un acido grasso a 16
443
Corpi chetonici (acetone)
atomi di carbonio, verranno formate durante il ciclo della β-ossidazione otto
unità a 2 atomi di carbonio. Queste 8 unità a 2 atomi di carbonio produrranno 8 × 12 = 96 ATP. Tuttavia 2 ATP sono consumati nell’attivazione iniziale dell’acido grasso. Inoltre, è stato calcolato che l’acido palmitico produce 35 ATP mentre passa attraverso il ciclo degli acidi grassi (7 FADH2, ciascuno equivalente a 2 ATP, e 7 NADH, ciascuno equivalente a 3 ATP).
Pertanto il numero netto di molecole di ATP prodotte sarà 96 – 2 + 35 =
129 (vedi Tabella 27.1).
Considerando che ciascuna mole di ATP richiede 7,6 kcal per la sua formazione, saranno necessarie 129 × 7,6 kcal, ossia 980 kcal. La produzione
teorica a partire da 1 mole di acido palmitico è di 2340 kcal, cosicché l’efficienza di conversione è pari a 980/2340, ossia 42%, mentre la rimanente
energia sarà prodotta sotto forma di calore. (Altri acidi grassi e il glicerolo
sono inoltre ossidati, cosicché il risultato netto è che i grassi producono
molta più energia rispetto ai carboidrati).
Tabella
ATP FORMATA DALL’OSSIDAZIONE DI UN ACIDO GRASSO CARBONIO-16
27.1
Fonte
Numero di molecole di ATP
formato per molecola di C16
7 FADH2
7 NADH
Attivazione iniziale di un grasso acido
8 acetil CoA
14
21
–2
96
129
27-4
Corpi chetonici (acetone)
In un paziente diabetico o in qualsiasi altra situazione in cui il metabolismo
dei carboidrati è limitato, l’organismo utilizza ossalacetato per produrre glucosio per il cervello e per i muscoli. Ciò riduce la quantità di ossalacetato
disponibile per il ciclo di Krebs e l’acetil CoA non può essere opportunamente metabolizzato. Quando ciò accade l’acetil CoA viene trasformato in
acetoacetil CoA, che viene a sua volta trasformato in acido acetoacetico nel
fegato dall’enzima deacilasi. L’acido acetoacetico può essere trasformato in
acetone ed in acido β-idrossibutirrico, come è mostrato nella Figura 27.3.
Queste tre sostanze - acido acetoacetico, acido β-idrossibutirrico e acetone - sono comunemente chiamate corpi acetonici o corpi chetonici. Essi
sono trasportati mediante il sangue ai muscoli ed ai tessuti, dove sono nuo-
tiolasi
acetil CoA
acetoacetil CoA
deacilasi
(nel fegato)
acido acetoacetico
β-idrossibutirrico
deidrogenasi
acido β-idrossibutirrico
Figura 27.3
Formazione dei corpi chetonici (acetone).
decarbossilazione
acetone
444
Metabolismo dei grassi
vamente trasformati in acetoacetil CoA e successivamente ossidati normalmente. Tuttavia nel diabete la produzione di queste sostanze da parte del
fegato eccede la capacità del muscolo e dei tessuti di ossidarle, per cui esse
si accumulano nel sangue.
CHETOSI
L’accumulo eccessivo di corpi chetonici è detto chetonemia. L’accumulo eccessivo di corpi chetonici nelle urine è detto chetonuria. L’accumulo complessivo di corpi chetonici sia nel sangue che nelle urine viene detto chetosi.
Durante la chetosi, l’acetone può essere rivelato nel respiro del paziente, poichè l’acetone è un composto volatile ed è facilmente escreto attraverso i polmoni. La chetosi può presentarsi nel diabete mellito, nel digiuno o in epatopatie gravi o in corso di diete ricche di grassi e povere in carboidrati.
Nel diabete mellito, l’organismo è incapace di ossidare i carboidrati e al
contrario, ossida i grassi; ciò comporta un accumulo di corpi chetonici nel
sangue e nelle urine. Questi corpi chetonici sono acidi e tendono ad abbassare il valore del pH del sangue. L’abbassamento del pH del sangue è detto
acidosi e può portare ad un coma fatale. Durante l’acidosi è necessaria una
maggiore quantità di acqua per eliminare i prodotti del metabolismo. A
meno che l’assunzione di acqua da parte del diabetico non venga aumentata, si può verificare disidratazione. La disidratazione dei diabetici può
anche essere causata da poliuria dovuta ad un aumento della quantità di
glucosio nelle urine.
Analogamente, durante il digiuno prolungato o durante una dieta ad alto
contenuto in grassi e basso contenuto in carboidrati, l’organismo tende a
bruciare grassi invece di carboidrati, con conseguente chetosi ed acidosi.
In caso di grave danno epatico, il fegato non può immagazzinare glicogeno nella quantità necessaria. La risultante carenza di carboidrati necessari per la normale ossidazione dei grassi conduce alla chetosi.
27-5
Disturbi metabolici causati da alterate ossidazioni degli acidi grassi
La malattia giamaicana del vomito è causata dall’ingestione di frutti
acerbi dell’albero akee. Questi frutti acerbi contengono ipoglicina, una
sostanza che inattiva l’acil-CoA-deidrogenasi, inibendo così la β-ossidazione e causando quindi ipoglicemia.
La malattia di Refsum è una rara malattia genetica causata dall’accumulo di acido fitanico. L’acido fitanico, a sua volta, blocca la β-ossidazione.
Il deficit di carnitina palmitoiltransferasi epatica comporta ipoglicemia
e un basso livello di corpi chetonici nel plasma.
Il deficit di carnitina palmitoiltransferasi muscolare comporta una difettosa ossidazione degli acidi grassi, e ciò causa debolezza muscolare.
27-6
Immagazzinamento dei grassi
I grassi in eccesso rispetto a quelli necessari per i normali processi ossidativi dell’organismo sono immagazzinati come tessuto adiposo sotto la cute e
intorno agli organi interni. Questo grasso immagazzinato serve per numerosi importanti scopi.
445
Immagazzinamento dei grassi
1.
2.
3.
4.
Riserva di cibo.
Sostegno per gli organi interni.
Ammortizzatore per gli organi interni.
Isolamento dell’interno dell’organismo da improvvisi cambiamenti
esterni della temperatura.
Il grasso immagazzinato nell’organismo è in equilibrio con quello nel
sangue. In altre parole, i grassi immagazzinati nel tessuto adiposo non
rimangono semplicemente lì come composti inerti fino a quando non sono
necessari. Essi sono continuamente usati e rimpiazzati e vi è sempre un trasporto dinamico di grassi tra il sangue ed i tessuti di deposito.
L’obesità (20% o più al disopra del peso normale) è una condizione in
cui un eccesso di grassi è depositato sotto forma di tessuto adiposo. Una
persona obesa mangia più cibo di quanto il suo organismo possa bruciare,
e l’eccesso è convertito in grasso e immagazzinato come tessuto adiposo.
Per ogni 9 kcal di cibo ingerito in eccesso rispetto al fabbisogno dell’organismo, si deposita 1 g di grasso.
La maggior parte delle persone ha una tendenza ad acquistare sovrappeso man mano che diventa più vecchia. Ciò è dovuto al fatto che esse
richiedono meno cibo per il mantenimento del loro organismo e fanno
meno esercizio fisico rispetto alle persone più giovani.
In genere l’obesità comporta una minore aspettativa di vita, come indicato nella Figura 27.4. Una persona in sovrappeso corre un rischio più elevato di sviluppare una malattia cardiovascolare, diabete o malattie epatiche. Un peso superiore del 10% a quello considerato normale per una persona di una certa età ed una data altezza è causa di problemi medici. La
risposta all’obesità consiste in una dieta appropriata sotto la supervisione di
un medico, perché il metabolismo dell’organismo è un meccanismo estremamente complicato che può essere disturbato molto facilmente.
Un eccessivo accumulo di trigliceridi nel fegato causa cirrosi ed alterata
funzione epatica. Questo accumulo può essere dovuto sia ad aumentati
livelli di acidi grassi liberi nel plasma, che a un blocco nella produzione di
Eccesso di mortalità*
UOMINI
DONNE
10%
sovrappeso
20%
sovrappeso
30%
sovrappeso
*confrontato con la mortalità per rischi standard (100%)
Figura 27.4
Tabella di mortalità. Le percentuali rappresentano l’eccesso di mortalità rispetto
alla mortalità per rischi standard (mortalità per rischio standard = 100%). (Per
gent. conc. di Metropolitan Life Insurance Co., New York.)
446
Metabolismo dei grassi
lipoproteine plasmatiche causato dagli acidi grassi liberi.
Anche l’alcolismo cronico può causare iperlipidemia e condurre infine a
cirrosi.
27-7
Lipogenesi
La lipogenesi - la conversione di glucosio in grassi - ha luogo nel fegato e nel
tessuto adiposo, con predominanza di quest’ultimo sito. L’insulina è necessaria per la lipogenesi sia nel fegato che nel tessuto adiposo. Il principale fattore che controlla l’andamento della lipogenesi è lo stato nutrizionale
dell’organismo. Se un individuo è a dieta ad alto contenuto in carboidrati, la
lipogenesi aumenta. Se un individuo è a dieta ristretta o ricca di grassi o
quando vi è un deficit di insulina come nel diabete mellito, la lipogenesi
diminuisce. Vi è anche un aumento della concentrazione degli acidi grassi
liberi nel plasma associato ad una diminuzione della velocità di lipogenesi.
La sintesi degli acidi grassi avviene nei mitocondri e nel citoplasma della
cellula, soprattutto in quest’ultimo compartimento. Nei mitocondri questo
processo consiste nell’allungamento delle catene di acidi grassi di moderata
lunghezza, mentre i processi citoplasmatici comportano la sintesi di acidi
grassi a partire da acetil CoA.
Le varie tappe nella sintesi degli acidi grassi a partire dall’acetil CoA
sono:
Tappa 1 L’acetil CoA è trasformato in malonil CoA.
O
O
||
||
Mn2+
CH3—C—S—CoA ⎯⎯⎯⎯→ CH2—C—S—CoA
ATP, biotina
|
COOH
acetil CoA
malonil CoA
Nota che è stato aggiunto un atomo di carbonio alla catena.
Tappa 2 Il malonil CoA reagisce con un’altra molecola di acetil CoA per
formare un complesso acetoacetilico.
O
O
O
O
||
||
||
||
CH2—C—S—CoA + CH3—C—S—CoA ⎯→ CH3—C—CH2—C—complesso + CO2 + H2O
|
COOH
malonil CoA
acetil CoA
complesso acetoacetil
Osservare che l’atomo di carbonio aggiunto nell’equazione precedente è stato rimosso. Esso era stato utilizzato principalmente per
attivare il carbonio α dell’acetil CoA in modo tale che la reazione
di condensazione potesse aver luogo. Il complesso enzimatico
richiesto in questa tappa è chiamato acido grasso-sintetasi ed è
composto da sette enzimi.
Tappa 3 Il gruppo chetonico del complesso acetoacetilico è ridotto al corrispondente alcool dal NADPH.