Quando nel XIX secolo gli scienziati rivolsero per la prima
volta la loro attenzione alla nutrizione, in breve tempo
scoprirono che i prodotti naturali contenenti azoto erano
essenziali per la nutrizione degli animali. Nel 1839 il
chimico svedese Jacob Berzelius coniò per questa classe
di composti il termine di PROTEINA (dal greco Proteius
cioè “che occupa la prima posizione).
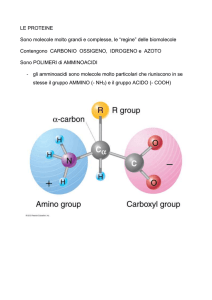
Gruppo R: catena laterale che
differenzia i diversi
amminoacidi. I carboni del
gruppo R sono indicati
attraverso le lettere
dell’alfabeto greco a partire
da β
Gruppo CARBOSSILICO :
gruppo acido
Gruppo AMMINICO: gruppo basico
Gli amminoacidi ( 20) hanno
proprietà strutturali comuni
1
I venti amminoacidi
costituenti le proteine,
variano considerevolmente
per le loro proprietà
chimico-fisiche come la
polarità, l'acidità, la
basicità, l'aromaticità, la
dimensione, la flessibilità
conformazionale, la
capacità di formare legami
crociati e legami idrogeno e
la reattività chimica. Queste
caratteristiche, molte delle
quali sono correlate, sono
in gran parte responsabili
della varietà di strutture e
funzioni delle proteine.
Tutti gli amminoacidi ad eccezione della glicina sono
otticamente attivi, cioè ruotano il piano della luce
polarizzata.
Le molecole
otticamente attive
sono asimmetriche
cioè non
sovrapponibili perché
posseggono un atomo
di carbonio chiralico
I centri chiralici danno origine ad ENANTIOMERI
2
ISOMERIA
Le molecole chirali esistono in due forme isomeriche che
sono speculari e non sovrapponibili, dette
Tutti gli a.a., eccetto uno
(la glicina), contengo un
atomo centrale chirale
(carbonio in α) .
Gli stereoisomeri di tutte
le molecole proteiche
esistenti in natura sono
L-STEREOISOMERI
3
Gli enantiomeri hanno proprietà fisiche identiche ad
eccezione nel modo in cui reagiscono con la
luce piano-polarizzata
La configurazione di un centro di
chiralità viene definita assegnando
le priorità relative dei quattro gruppi.
La molecola a più bassa priorità
viene orientata opposta agli altri tre
a più alta priorità che si dirigono
verso l’osservatore.
Se per passare dal gruppo a più
alta priorità a quello a più bassa,
passando per il gruppo a priorità
intermedio, si percorre una curva in
senso antiorario, lo steroisomero
avrà configurazione S, se in senso
orario avrà configurazione R
FORMULE
GEOMETRICHE
PROIEZIONE DI
FISCHER
4
OGNI AMMINOACIDO DERIVANTE DA UNA PROTEINA E’
NELLA CONFORMAZIONE STECHIOMETRICA “ L”.
Gli a.a. sono classificati in base alla capacità del loro
gruppo R di interagire con l’H2O
Sono presenti 20 catene laterali
diverse che variano per:
Dimensioni
Carica
Capacità di formare legami idrogeno
Idrofobicità
Reattività chimica
Suddivideremo gli amminoacidi in
quattro gruppi in base alle
caratteristiche generali dei loro
gruppi R:
a.a. IDROFOBICI
a.a. POLARI
a.a. AROMATICI
a.a. CARICHI NEGATIVAMENTE
5
6
7
La prolina influenza
profondamente
l’architettura delle
proteine poiché la sua
struttura ad anello
determina più costrizioni
conformazionali nelle
proteine rispetto agli altri
a.a.
8
9
10
Gli a.a. aromatici presentano un picco di
assorbimento alla lunghezza d’onda di
280 nm .
11
Sono a.a. carichi
positivamente a pH neutro
La lisina termina con un
gruppo amminico primario
L’arginina con un gruppo
guanidinico
L’istidina contiene un
gruppo IMIDAZOLICO che
vicino a pH fisiologico
può prendere o
rilasciare protoni
12
Questi a.a. sono chiamati
spesso glutammato e aspartato
per sottolineare che a pH
fisiologicoper sottolineare che
le loro catene laterali sono
deprotonate
13
Le proprietà strutturali e chimiche dei 20 a.a. permettono alle
proteine di svolgere numerosi ruoli funzionali. Esistono altri a.a.
che non fanno parte della struttura delle proteine perché troppo
reattivi vcome l’omocisteina che potrebbe formare strutture
cicliche non adatte.
L’omocisteina può
ciclizzare e formare un
anello stabile a cinque
termini, che
potenzialmente
potrebbe scindere il
legame peptidico.
La ciclizzazione della
serina è invece
impedita perché
l’anello a 4 termini è
instabile
Altri a.a., diversi dai venti elencati, possono far parte di alcune
strutture proteiche ma generalmente derivano da modificazioni
specifiche di un residuo a.acidico standard dopo la sintesi della
catena polipeptidica su cui è inserito
14
Gli amminoacidi in soluzione a pH neutro esistono in
forma di ioni dipolari chiamati ZWITTERIONI
Quando un amminoacido che non possiede un gruppo R ionizzabile
viene posto in acqua a pH neutro, si trova in soluzione sottoforma di
ione bipolare, o ZWITTERIONE. Possiede uno ione basico ( gruppo
amminico) ed uno acido (groppo carbossilico) perciò può
comportarsi come acido o come base.
In soluzione acquosa gli a.a. si comportano cioè
come basi ed acidi deboli
15
Curva di titolazione della GLICINA
La curva di titolazione
mostra due fasi distinte,
corrispondenti ai due
differenti gruppi titolabili
della glicina.
A pH bassi prevale la forma
completamente protonata
(pK1) a pH alti quella
deprotonata (pK2). A metà
tra i due pK troviamo il
PUNTO ISOELETTRICO (pI)
il pH in cui la glicina si
trova nella sua forma
bipolare zwitterionica e
dove la sua carica netta è
zero.
LO STATO DI IONIZZAZIONE DIPENDE DAL PH DELLA SOLUZIONE
16
Gli amminoacidi con un gruppo R ionizzabile hanno curve di
titolazione più complesse, con tre fasi corrispondenti alle tre possibili
tappe di ionizzazione.
17
Nel 1953 Frederick Sanger determinò la sequenza degli
amminoacidi dell’insulina, un ormone proteico. Il suo lavoro è
stato una pietra miliare nello sviluppo della biochimica perché
ha dimostrato per la prima volta che le proteine hanno
sequenze amminoacidiche definite con grande precisione,
costituite da L-amminoacidi legati con legame peptidico.
18
Due molecole di amminoacidi possono unirsi
covalentemente mediante un legame ammidico, chiamato
LEGAME PEPTIDICO, formando un DIPEPTIDE
Questo tipo di legame avviene tra il gruppo carbossilico
di un amminoacido ed il gruppo amminico del secondo
amminoacido per eliminazione di una molecola di acqua
19
Il legame che deriva dalla
condensazione di αamminoacidi (con eliminazione
di una molecola di H2O) viene
chiamato legame peptidico:
Struttura rigida planare per le interazioni
di risonanza che forniscono al legame
peptidico circa il 40% di carattere di
doppio legame
Caratteristiche del legame peptidico
I legami peptidici non si
rompono nelle condizioni che
denaturano le proteine:
riscaldamento o
concentrazioni elevate di
urea. Per idrolizzare questi
legami, non
enzimaticamente, occorrono
esposizioni a acidi o basi
forti con temperature elevate
20
Nomenclatura dei polimeri amminoacidici
I polimeri (di tipo poliammiacidico) contenenti due, tre,
pochi (da tre a dieci) oppure molti residui amminoacidici
sono detti rispettivamente dipeptidi, tripeptidi, oligopeptidi
oppure polipeptidi. I polipeptidi sono polimeri lineari che
possono contenere da circa 40 fino a più 4000 residui
amminoacidici e, poiché la massa media degli amminoacidi
è di 110 Da, essi hanno masse molecolari che variano da
circa 4 a 440 kDa.
Nel peptide, il residuo amminoacidico con cui termina la
catena polipeptidica, viene chiamato residuo
AMMINOTERMINALE (n-terminale), ha il gruppo
amminico libero; il residuo all’altra estremintà ha un
gruppo α-carbossilico libero e viene chiamato residuo
CARBOSSITERMINALE (C-terminale).
Quando si riporta la fotografia di un polipeptide
l’estremità amminoterminale viene posta a sinistra
quella carbossiterminale a destra
21
La struttura delle proteine viene suddivisa in quattro livelli di
organizzazione:
La funzione delle proteine può essere compresa analizzando la loro
struttura. La struttura delle proteine viene suddivisa in quattro livelli
di organizzazione:
1) La struttura primaria è la sequenza amminoacidica di una catena
polipeptidica;
22
Struttura delle proteine
23