il sistema olfattivo umano
(uno studio teorico a livello di reti neuronali)
by
Giovanni Del Papa
http://www.ing-gdp.com
2
Indice
1.0 I componenti di base del SNC: neuroni , sinapsi, assoni e dendriti. ............................ 3
1.1 I neuroni ............................................................................................................... 3
1.2 Le sinapsi con mediatore chimico ....................................................................... 5
1.3 La generazione del potenziale a punta (spike). .................................................... 7
2.0 Schematizzazione del sistema nervoso. ........................................................................... 9
2.1 Qualche cenno di anatomia. ................................................................................. 9
2.1.1 L'encefalo.
2.1.2 Proencefalo
2.1.3 Corteccia cerebrale e lobo olfattivo
2.1.4 Sistema limbico
2.1.5 Diencefalo
2.1.6 Talamo
2.1.7 Ipotalamo
2.1.8 Epitalamo
2.1.9 Subtalamo
2.1.10 Tronco encefalico (o tronco cerebrale)
2.1.11 Mesencefalo
2.1.12 Ponte
2.1.13 Formazione reticolare.
2.1.14 Mielencefalo
2.1.15 I nervi cranici
2.1.16 il midollo spinale e il Cervelletto
3.0 Il sistema olfattivo nell'uomo .......................................................................................... 16
3.1 Organizzazione generale
3.2 Disegno del sistema olfattivo in generale
4.0 Un modello della codifica degli odori mediante spazi vettoriali. .................................... 26
4.1 Attivita' delle cellule recettrici. ............................................................................ 27
4.1.1 Convergenza recettori-glomeruli.
4.1.2 Proiezione glomeruli-cellule mitrali
4.1.3 Inibizione laterale.
4.2 Una descrizione alternativa: immagini di attivita' .............................................. 36
4.2.1 Immagine della attivita' glomerulare
4.2.2 Ingresso alla cellula mitrale
4.2.3 Immagine dell'uscita della cellula mitrale.
4.2.4 Due tipi di discriminazione nel sistema olfattivo(ordine spaziale e inibizione laterale)
4.2.5 Riconoscimento qualitativo e memoria associativa
4.2.6 Costruzione di un sensore di odori intelligente artificiale.
Bibliografia. ........................................................................................................................... 41
2
10
11
11
12
12
12
13
13
13
13
13
14
14
14
14
14
19
22
30
33
35
36
37
38
38
39
39
3
1.0 I componenti di base del SNC: neuroni ,
dendriti.
sinapsi, assoni e
1.1 I neuroni
Pur essendo di vario tipo [Murray L. Barr. il sistema nervoso umano. Piccin 1993. pp. 9-26]
tutti i neuroni hanno in comune la struttura di base e il loro funzionamento. Il neurone e' la
cellula nervosa elementare. Il nucleo della cellula e' contenuto nel corpo cellulare o soma .
Per quanto riguarda le proprieta' funzionali delle cellule nervose, il nucleo ha una scarsa
importanza. Dal corpo cellulare, da un punto di nome colle, prende origine l'assone .
3
4
L'assone spesso risulta molto lungo; alla fine dell'assone vi puo' essere un' arborescenza, per
connettersi con piu' di una unita'. L'assone, con la sua arborescenza, costituisce l' uscita del
neurone.
Al neurone fanno capo numerosi dendriti. I dendriti insieme al soma formano l'organo di
entrata del neurone. Le sinapsi presiedono al passaggio dei segnali da e per il neurone . Le
sinapsi
sono poste sia vicino alla superficie esterna dei dendriti sia a quella del corpo
cellulare.
Si fa la seguente classificazione dei neuroni:
a)interneurone: entrata neurone - uscita neurone
b)neurone afferente (o sensitivo): entrata recettore - uscita neurone
c)neurone efferente: entrata neurone - uscita effettore
Gli assoni si distinguono in assoni mielinici e assoni amielinici, a seconda che l'assone sia
rivestito o meno da una guaina mielinica. I neuroni possono essere di varie dimensioni. Per
esempio il diametro di un assone non mielinico varia circa da 0,3 micron a 1,3 micron.
Alcuni tipi di neuroni:
a)neurone bipolare o pseudounipolare. e' composto, oltre che dal corpo cellulare, da due
prolungamenti assonici (uno dei quali in effetti di tipo afferente).
b)neurone bipolare. E' composto, oltre che dal soma, da un dendrite e da un assone. A
questo gruppo appartengono per esempio quelli del sistema olfattivo.
c)neuroni multipolari. Hanno sempre ramificazioni dendritiche ed assoniche. Questi ultimi
possono a loro volta suddividersi nei seguenti tipi:
1)Cellule di Purkinje. Hanno una complessa e caratteristica ramificazione dendritica Si
trovano cellule di questo tipo per esempio nel cervelletto.
2)Cellula piramidale . Ha una notevole ramificazione dendritica .
3)Cellula di Golgi del II tipo. E' caratterizzata dall'avere un assone piuttosto corto.
4)Cellula di Golgi del I tipo. Sono simili a quelle del II tipo, ma con un assone molto piu'
lungo.
4
5
5)Cellula di motoneurone. E' caratterizzata dalla presenza di un assone molto lungo, le cui
ramificazioni si innestano nelle placche motrici del muscolo.
Le sinapsi sono le unita' di collegamento attivo di un neurone con:
a)un altro neurone
b)con un recettore
c) con un effettore
Anche le sinapsi sono di vario tipo. Per comprendere il funzionamento di una sinapsi, occorre
distinguere:
la membrana presinaptica e la membrana postsinaptica. La membrana
presinaptica puo' essere considerata come la terminazione della cellula afferente, che si
sviluppa formando i bottoni sinaptici. La membrana presinaptica contiene un certo numero di
vescicole, addensate a grappolo vicino alla membrana stessa. Fra le due membrane vi e' la
fessura sinaptica,
o
delle dimensioni approssimativamente di 200 A . Il diametro delle
vescicole e' invece di
o
300 A . Le membrane che formano il contatto sinaptico sono
o
generalmente piu' spesse (circa 300 A ), rispetto allo spessore esistente nelle altre parti della
Il meccanismo della trasmissione nelle sinapsi e' il seguente: un impulso elettrico che arriva
cellula.
al bottone sinaptico, provoca la trasmissione di un mediatore chimico attraverso le due
membrane costituenti il contatto sinaptico. Quando questo mediatore ha attraversato la
membrana postsinaptica, viene rapidamente distrutto, provocando pero' una variazione dello
stato elettrochimico della membrana postsinaptica. Tenuto conto dell'effetto di tutti i contatti
sinaptici, quando lo stato elettrico della membrana del corpo cellulare ha raggiunto particolari
modificazioni, si ha la generazione al colle dell'assone di un impulso elettrico che si propaga
lungo l'assone, fino ad arrivare alle terminazioni assoniche.
1.2 Le sinapsi con mediatore chimico
5
6
[Sir John Eccles 1965. La sinapsi. Le Scienze Marzo 1965. pp 59-68]
Se si misura la tensione esistente a riposo fra un punto, posto all'interno della cellula, in
vicinanza della membrana postsinaptica e un secondo punto, posto all'esterno della cellula, si
scopre che il potenziale di riposo postsinaptico e' dell'ordine di grandezza di 70mV, con
interno negativo, rispetto all' esterno della cellula. All'arrivo di uno stimolo alla membrana
presinaptica, si ha una variazione del potenziale postsinaptico. A seconda della sinapsi , si
puo' avere un effetto eccitatorio (effetto depolarizzante) oppure un effetto inibitorio (effetto
iperpolarizzante). Gli stimoli relativi alle sinapsi eccitatorie fanno variare il potenziale
postsinaptico, nel senso di rendere l'interno meno negativo rispetto all'esterno. Si ritiene che
le sinapsi inibitorie possano essere di due tipi diversi:
a) sinapsi inibitrici del I tipo che fanno variare il potenziale postsinaptico nel senso di
rendere l'interno piu' negativo rispetto all'esterno, cioe' sinapsi che hanno un effetto analogo a
quelle eccitatrici, ma di segno opposto
b)sinapsi inibitrici del II tipo
che,
indipendentemente dal potenziale postsinaptico
esistente, tendono a riportare questo potenziale al valore del potenziale di riposo.
Considerando il funzionamento del neurone sotto soglia in alcune sinapsi,
si ha poi il
fenomeno della facilitazione in cui, a seguito dell'arrivo di una serie di impulsi eccitatori, le
6
7
variazioni del potenziale postsinaptico via via aumentano. L'effetto opposto e' quello della
defacilitazione
in cui, a seguito dell'arrivo di una serie di impulsi,
le variazioni
postsinaptiche diminuiscono.
Quando la membrana postsinaptica si trova in condizioni diverse da quelle di riposo, in
assenza di stimoli susseguenti,
non considerando ancora il caso che si sia giunti alle
condizioni atte a generare il potenziale a punta (spike),
avviene che il potenziale
postsinaptico tende a ritornare al valore di riposo con una legge di tipo esponenziale. La
costante di tempo di questo esponenziale puo' essere di qualche centinaio di microsecondi ed
anche di qualche millisecondo.
1.3 La generazione del potenziale a punta (spike).
[P. Strata Fisiologia del neurone E. M. S. I. Roma 1983 pp. 10-20]
Se si misura la tensione esistente fra un punto interno alla cellula nella zona del colle
dell'assone ed un punto esterno, si trova che, in condizioni di riposo, tale tensione risulta
essere ancora dell'ordine di grandezza di 70mV (negativo l'interno rispetto all'esterno). Per
effetto dei contatti sinaptici, agenti sulla cellula, questa tensione varia nel tempo, risentendo
delle variazioni dei potenziali postsinaptici delle parti della membrana, nelle quali arrivano
gli stimoli. Nel caso che il potenziale al colle dell'assone vari in senso opposto, rispetto alle
condizioni di riposo, fino a che non si raggiunge un particolare livello di soglia, nulla
avviene. Al momento invece nel quale si raggiunge questo livello di soglia (il cui valore
numerico si puo' ritenere che sia dell'ordine di 30 mV, con ancora l'interno negativo rispetto
all'esterno) , l'oscillogramma del potenziale in questo punto risulta modificarsi in modo
radicale e la sua susseguente forma d'onda e' del tutto indipendente, almeno per un certo
intervallo di tempo, dai susseguenti stimoli agenti sul neurone (in generale si dice che il
neurone funziona sotto soglia o sopra soglia, a seconda che non si sia o si sia raggiunto
questo valore).
7
8
A questa forma d'onda si da' il nome di potenziale a punta (spike) od anche potenziale
d'azione. Questo impulso, generato al colle dell'assone, si propaga lungo l'assone. Essendo
questo l'unico tipo di informazione che si trasmette lungo il sistema nervoso,
si puo'
concludere che il sistema nervoso funzioni, almeno nella sua parte trasmissiva, come un
sistema nel quale i segnali sono tutto o niente; cioe' viene adoperata una codificazione delle
informazioni di tipo binario (anche se si ritiene che la frequenza con cui si manifestano questi
impulsi sia il dato veramente significativo [ J. Singh. Linguaggio e cibernetica. EST
Mondadori. 1976. pp152-153] ). Il valore di soglia, per un medesimo neurone, puo' variare
sia al variare delle condizioni esterne di funzionamento sia della presenza o meno di impulsi
precedenti.
A questo riguardo si parla di :
a)periodo di refrattarieta' assoluta: durante tale periodo la soglia ha valore infinito
b) periodo di refrattarieta' relativa: indica uno stato del neurone tale per cui si ha la
generazione dell'impulso soltanto se il potenziale al colle dell'assone raggiunge un valore
maggiore di quello corrispondente alla soglia nominale;
c)periodo di sovraeccitabilita': indica uno stato del neurone al quale corrisponde un valore
di soglia inferiore al valore nominale. Il fenomeno della refrattarieta' mostra che vi e' una
frequenza massima di impulsi trasmettibili dalle singole fibre nervose. Risulta di notevole
importanza la fenomenologia riguardante i seguenti due concetti: sommazione temporale e
sommazione spaziale.
La sommazione temporale non e' altro che l'effetto integrale operato dalla membrana, nel
tempo, vista come un condensatore.
La sommazione spaziale significa che un punto qualsiasi della membrana risente delle
variazioni elettriche prodotte dai vari contatti sinaptici che agiscono sul neurone.
Si potrebbe ora passare alla descrizione del meccanismo con cui si genera il potenziale
d'azione, partendo dalla descrizione della membrana neuronica
per poi passare alla
descrizione dei meccanismi di conduzione , ma si preferisce tralasciare questi argomenti,
8
9
perche' non sono rilevanti dal punto di vista del nostro studio della codifica degli odori nel
sistema olfattivo.
(per un trattazione matematca anche se piuttosto sintetica del neurone si rimanda a :[Tarun
Khanna "fondamenti di reti neurali. Addison Wesley 1991. pp 144-156]
2.0 Schematizzazione del sistema nervoso.
Il Sistema Nervoso centrale (SNC) svolge la funzione fondamentale di elaborazione e
riconoscimento dei segnali che gli giungono dai sensori esterni ed interni al corpo. In
secondo luogo presiede al controllo dei movimenti muscolari ed al sistema ghiandolare
mediante l'invio di opportuni segnali. Il sistema nervoso centrale e' poi la sede delle attivita'
superiori quali la creativita', intelligenza etc.
La trasmissione dei segnali, nel sistema nervoso, viene effettuata dalle fibre nervose
prevalentemente costituite dagli assoni dei neuroni.
Il SNC comunica poi in ingresso ed in uscita mediante unita' periferiche.
In entrata abbiamo i recettori che si dividono in esterocettori (ricevono informazioni
dall'esterno dell'organismo) e in propriocettori che sono responsabili della misura
delle
grandezze (chimiche o fisiche) nell'interno del corpo. In uscita abbiamo gli effettori che
agiscono sia sui muscoli sia sulle ghiandole.Essi si suddividono in :
a)effettori volontari.
b) effettori riflessi (agiscono in modo diretto a stimoli senza intervento del SNC)
c)effettori involontari, che rappresentano le uscite del sistema nervoso autonomo .
2.1 Qualche cenno di anatomia.
fig. tratta da: A. R. Luria L'organizzazione funzionale cerebrale. Le Scienze giugno 1970
9
10
Sulla base della descrizione anatomica del cervello effettuata da [Luigi Cattaneo. Anatomia
del sistema nervoso centrale e periferico dell'uomo. Monduzzi editore 1984. ] diciamo che
L'encefalo e il midollo spinale costituiscono il sistema nervoso centrale.
E' possibile suddividere le varie parti del sistema nervoso ottenendo la seguente suddivisione:
a)Encefalo
b)Nervi cranici
c)midollo spinale
d)nervi spinali
gli elementi costitutivi del sistema nervoso sono i seguenti:
a)neuroni
b)sinapsi
c) trasduttori di entrata
d) trasduttori di uscita.
2.1.1 L'encefalo.
10
11
L'encefalo puo' essere suddiviso in:
proencefalo, mesencefalo, romboencefalo e
mielencefalo.
L' encefalo e' composto di sostanza grigia e sostanza bianca. La sostanza grigia contiene i
corpi cellulari dei neuroni, mentre la sostanza bianca e' costituita da fibre nervose per i
collegamenti fra i vari neuroni. Lo strato piu' esterno dell'encefalo, detto corteccia, e'
composto di sostanza grigia.
La superficie della corteccia e' sede di molte ripiegature (circonvoluzioni) al fine di aumentare
la superficie e poter cosi' contenere un maggior numero di neuroni a parita' di volume. Il SNC
dell'uomo contiene circa 1010 neuroni, il 90% dei quali ha sede nell'encefalo.
Vi sono poi altre zone dell'encefalo costituite da sostanza grigia dette nuclei.
2.1.2 Proencefalo
Si suddivide in: telencefalo e diencefalo. Il telencefalo si suddivide in : corteccia cerebrale
e sistema limbico. Il diencefalo comprende molti nuclei ; il piu' importante e' il talamo, vi
sono poi l'ipotalamo, l'epitalamo ed il subtalamo.
2.1.3 Corteccia cerebrale e lobo olfattivo
La corteccia cerebrale e' divisa in vari settori mediante scissure e solchi. Si individuano
cosi' i due emisferi destro e sinistro. Ciascun emisfero si suddivide in 4 lobi: frontale,
parietale, temporale ed occipitale. Poi c'e' il lobo olfattivo, che puo' essere visto, dal
punto di vista funzionale, appartenente alla corteccia cerebrale; spesso lo si considera
parte del sistema limbico.
Da un punto di vista funzionale, la corteccia cerebrale e' suddivisibile in 4 grandi zone .
area sensitiva alla quale arrivano segnali dai vari sistemi sensoriali. Ogni senso ha
una propria area principale e talora alcune aree secondarie .
11
Nelle aree
12
secondarie spesso si hanno effetti dovuti a stimoli provenienti da piu' organi
sensoriali.
area motoria comanda l'insieme dei muscoli che soprassiedono alla postura e alla
locomozione.
3)Area delle azioni volitive e decisionali.
Guardando in dettaglio le aree sensoriali, si noti che l'area dell'odorato si trova nel lobo
olfattivo; l'area del gusto ha sede nel lobo parietale; le aree tattili si trovano nel lobo
parietale; le aree acustiche si trovano nel lobo temporale; le aree visive sono poste nel
lobo occipitale .
2.1.4 Sistema limbico
E' un sistema subcorticale, sede del sistema olfattivo. Inoltre si ritiene che il sistema limbico
abbia qualche importanza per il comportamento affettivo ed emozionale,
(fame, paura,
aggressivita', attrazione sessuale). Inoltre nel sistema limbico vi sono anche alcuni nuclei, i piu'
importanti dei quali sono il corpo striato ( diviso in nucleo caudato e nucleo lenticolare), il
nucleo amigdale, il nucleo claustro e l'ippocampo. I nuclei del corpo striato fanno parte
delle vie motorie .
E' stata anche avanzata l'ipotesi [Olton David S. La memoria spaziale in Le scienze 110 ,
ottobre 1977] che l'ippocampo possa risultare importante per la memorizzazione delle
informazioni circa la dislocazione spaziale del soggetto rispetto all'ambiente.
2.1.5 Diencefalo
Il Diencefalo comprende molti nuclei. I piu' importanti sono: talamo, ipotalamo, epitalamo,
subtalamo.
2.1.6 Talamo
12
13
Passano attraverso questo nucleo tutti i segnali dalle vie sensoriali verso la corteccia
2.1.7 Ipotalamo
L'ipotalamo svolge la funzione di regolatore di funzioni di tipo vegetativo, quali per esempio
regolazione della temperatura e del metabolismo idrico.
2.1.8 Epitalamo
E' una parte del sistema olfattivo.
2.1.9 Subtalamo
Il subtalamo che serve da collegamento fra la corteccia cerebrale motoria ed il tronco
cerebrale, fa parte delle vie motorie cosiddette extrapiramidali.
2.1.10 Tronco encefalico (o tronco cerebrale)
Attraverso il tronco encefalico passano quasi tutte le vie sensitive e motorie. E' suddiviso
nelle tre parti:
mesencefalo, romboencefalo e midollo allungato (o bulbo). Il
romboencefalo a sua volta, e' divisibile in due parti: il ponte ed il cervelletto. Fa parte del
tronco encefalico la formazione reticolare, che ha sede principalmente nel mesencefalo e nel
bulbo.
2.1.11 Mesencefalo
Fanno parte del mesencefalo: i nuclei del collicolo superiore e del collicolo inferiore, i
nuclei dei nervi oculomotori . Questi nuclei delle vie sensitive spesso sono denominati come
nuclei del corpo (o lamina) quadrigemino.
Fa parte del mesencefalo anche il nucleo rosso.
13
14
2.1.12 Ponte
Fanno parte dell ponte: il nucleo lemnisco laterale (con il fascio di collegamenti denominato
appunto lemnisco laterale); il nucleo trapezoideo ( con il fascio di collegamenti denominato
corpo trapezoideo).
Il ponte e' un nucleo . E' collegato anche al cervelletto, alla corteccia cerebrale, al bulbo e al
midollo spinale.
2.1.13 Formazione reticolare.
E' una massa connettiva, posta alla base dell'encefalo, si ritiene che la formazione reticolare
eserciti una notevole azione di integrazione tra le varie vie sensitive e motorie. La formazione
reticolare e' importante anche per determinare lo stato di attenzione (vedi ART), di veglia o
di sonno.
2.1.14 Mielencefalo
Il mielencefalo (midollo allungato) e' il prolungamento del midollo spinale. Fanno parte del
midollo allungato i nuclei olivari .
2.1.15 I nervi cranici
Dalla parte inferiore dell'encefalo si dipartono 12 paia di nervi, chiamati nervi cranici.
2.1.16 il midollo spinale e il Cervelletto
Non hanno importanza nel sistema olfattivo, quindi rinunceremo alla loro descrizione.
Stessa cosa dicasi per i nervi spinali.
Dopo questa visione di insieme andiamo ad analizzare in dettaglio il sistema olfattivo.
14
15
15
16
3.0 Il sistema olfattivo nell'uomo
I recettori olfattivi posseggono siti ricettivi specializzati per certi odori. John Amoore e coll.
(Amoore, J. . E. , Johnson, W. , Jr, and Rubin, M. 1964. The stereochemical theory of
odor. Sci. Am. 210(2): 42-49)
hanno esaminato piu' di cento sostanze odorifere ed hanno supposto che esistano almeno sette
odori primari: l'odore di canfora, di muschio, di fiori, di menta, l'odore etereo, pungente e
l'odore putrido. Le sostanze appartenenti a una certa categoria hanno configurazioni
molecolari simili, tuttavia, sebbene molti odori ricadano in queste sette categorie, vi sono
molte eccezioni. Odori simili non sono sempre prodotti da molecole con strutture analoghe.
Gli uomini possono distinguere fra migliaia di specie odorifere. Noi siamo in grado di
rilevare la presenza anche di solo 108 molecole di una certa sostanza odorifera sparsa in un
ambiente di centinaia di m3. La soglia che permette di apprezzare la presenza di un odore e'
inferiore a quella che consente il riconoscimento specifico. Se le concentrazioni delle sostanze
odorifere sono molto basse, la sensazione e' aspecifica; solo a concentrazioni maggiori e'
possibile identificare con esattezza il tipo di odore.
(Shepherd, 1972, Synaptic organization of mammalian olfactory bulb. Physiol. Rev. 52864-917)Il senso dell'olfatto e' dovuto a recettori olfattivi situati nella profondita' delle cavita'
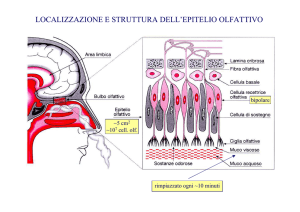
nasali. Nella specie umana i recettori sono limitati a un tratto di epitelio specializzato, di
color bruno giallastro, che costituisce l'epitelio olfattivo e che copre all'incirca 5 cm2 del
recesso dorsale della cavita' nasale. Quest' epitelio contiene tre tipi di cellule: i recettori, le
cellule di sostegno e le cellule basali. I recettori sono cellule nervose bipolari che hanno un
tratto periferico breve e un lungo tratto centrale. Il tratto periferico breve si estende verso la
superficie della mucosa, dove si espande in un piccolo corpicciolo che da' origine a parecchie
ciglia. Queste ciglia formano un groviglio denso sulla superficie della mucosa e si ritiene che
16
17
interagiscano con molecole odorifere. Il tratto centrale lungo e' costituito da un assone
amielinico che va dalla cavita' nasale al bulbo olfattivo. Da 10 a 100 di questi assoni si
associano in fascetti circondati da cellule di Schwann. L'insieme di questi fascetti di assoni
viene detto nervo olfattivo, che e' il primo paio di nervi cranici. Prima di raggiungere il bulbo
olfattivo, gli assoni devono passare attraverso la lamina cribrosa dell'osso etmoide.
figure tratte da: [ Kandel, E. R. & Schwartz, J. H. (1982) Principles of Neural Science.
New York: Elsevier/North Holland Capitolo 32 I sensi chimici: gusto e olfatto].
Il bulbo olfattivo e' una parte del sistema nervoso centrale che prende origine dal telencefalo.
Esistono due bulbi olfattivi simmetrici deputati alla mucosa olfattiva delle due cavita' nasali,
percio', a differenza delle altre modalita' sensoriali, la via olfattiva ha le prime sinapsi nel
telencefalo.
17
18
Le cellule recettrici olfattive sono piu' di 108. I recettori olfattivi
si ricambiano
completamente ogni 60 giorni a partire dalle cellule basali che ne sono i precursori. I recettori
olfattivi costituiscono gli unici neuroni del sistema nervoso dell'adulto, in grado di andare
incontro a mitosi cellulare. (Da questo fatto prendono spunto alcuni modelli di tipo neuronale
impiegati in sistemi artificiali per la discriminazione di odori, in cui i neuroni possono
aumentare di numero dinamicamente[Evor L. Hines, Claire C. Gianna and J. W Gardner.
Neural network based electronic nose using constructive algorithms.
Techniques and
application of neural networks. 1993 Ellis Horwod Workshop. pp135-154] ). Le cellule che si
sviluppano devono mandare il dendrite neoformato verso la superficie della mucosa e l'assone
nella direzione opposta per stabilire sinapsi nel bulbo olfattivo.
Le molecole odorifere stimolano le cellule olfattive, in quanto vengono adsorbite in primo
luogo nello strato mucoso. [S. G. Shirley and Krishna C. Persaud. The biochemistry of
vertebrate olfaction and taste. The neurosciences. Vol II 1990: pp 59-68]Le molecole
diffondono quindi verso le ciglia dei neuroni recettori e si legano a siti specifici che si
occupano del riconoscimento molecolare . Si ritiene che il legame di una specifica sostanza
odorifera a un sito della cellula recettrice, deputato al riconoscimento molecolare, determini
l'apertura di canali ionici specifici per gli ioni Na+ e i K+, [Eleonora P. Giorgi. Il senso
dell'olfatto. Le Scienze num. 101. Gennaio 1977] generando in tal modo un potenziale di
recettore depolarizzante. Le cellule recettrici posseggono un'attivita' spontanea. Il potenziale
18
19
di recettore, determinato dai diversi tipi di stimolazione olfattiva, provoca un aumento della
frequenza di scarica di tipo graduato; quanto maggiore e' la stimolazione olfattiva e quanto
piu' ampio il potenziale di recettore, tanto maggiore e' la frequenza di scarica. Anche le
cellule di sostegno, che circondano i recettori, rispondono alla stimolazione odorifera con
piccole depolarizzazioni o iperpolarizzazioni, ma non generano potenziali d'azione. Studi
elettrofisiologici, fatti da Gordon Sheperd e Thomas Getchell, a Yale, [Getchell, T. V. , and
Sheperd, G. M. 1978. Responses of olfactory receptor cells to step pulses of odour at
different concentrations in the salamander. J. Physiol. (Lond. ) 282: 521-540. ] mediante
l'impiego di quantita' controllate di sostanze odorifere, dimostrano che i singoli recettori sono
particolarmente sensibili per una o due diverse categorie di odori. Mediante l'uso di 2deossiglucosio marcato per lo studio dell'attivita' metabolica dei diversi neuroni, Sheperd,
William Stewart e John Kauer [Stewart, W. B. , Kauer, J. S. , and Shepherd, G, M 1979
Functional organization of rat olfactory bulb analysed by the 2-deoxyglucose method. J.
Comp. Neurol. 185: 715-734] hanno cercato di determinare quali cellule del bulbo olfattivo
rispondono a sostanze odoranti specifiche e hanno dimostrato l'esistenza di particolari linee
private. Questi autori hanno osservato che i diversi tipi di odori evocano attivita' di tipo
diverso nelle diverse zone del bulbo olfattivo, percio', analogamante a quanto accade negli
altri sistemi sensoriali, la codificazione ha luogo in particolari linee private modificate. Nel
caso del nervo olfattivo le diverse linee private distribuiscono la propria attivita' a zone
diverse del bulbo olfattivo .
A differenza degli altri sistemi sensoriali, nei quali le localizzazioni centrali contengono una
rappresentazione topografica in termini di coordinate spaziali del mondo fisico, il bulbo
olfattivo ed i centri olfattivi superiori impiegano il proprio spazio neurale non per
rappresentare lo spazio fisico del mondo esterno, ma per dare una rappresentazione distinta
dei diversi odori fondamenteli ossia codificano uno spazio degli odori .
3.1 Organizzazione generale
19
20
figura tratta da [ Kandel, E. R. & Schwartz, J. H. (1982) Principles of Neural Science.
New York: Elsevier/North Holland Capitolo 32 I sensi chimici: gusto e olfatto].
I piccoli assoni amielinici dei recettori olfattivi terminano nel bulbo olfattivo, che e' il primo
centro di ritrasmissione del sistema olfattivo . All'interno del bulbo olfattivo, gli assoni dei
recettori olfattivi fanno sinapsi (in formazioni sferiche specializzate dette glomeruli) con
diversi tipi di cellule e in particolare con grandi cellule mitrali e piccole cellule a
pennacchio, che sono i principali neuroni efferenti . Gli assoni delle cellule mitrali e delle
cellule a pennacchio si immettono nel tratto olfattivo e si distribuiscono ai centri olfattivi
secondari che vengono anche chiamati corteccia olfattiva.
Questo tipo di corteccia puo' venir suddivisa in cinque parti: (1) il nucleo olfattivo anteriore
(che connette i due bulbi olfattivi attraverso una parte della commessura anteriore; (2) il
20
21
tubercolo olfattivo, (3) la corteccia piriforme ( che e' il principale centro di discriminazione
olfattiva), (4) il nucleo corticale dell'amigdala e (5) l'area entorinale, che proietta a sua volta
nell'ippocampo . Tutte queste aree corticali fanno parte della paleocorteccia (paleocortex). E'
chiaro tuttavia che l'analisi olfattiva non ha termine nel paleocortex. Come per tutti gli altri
sistemi sensoriali, le informazioni olfattive vengono alla fine ritrasmesse al talamo e al
neocortex. Il tubercolo olfattivo proietta al nucleo dorso-mediale del talamo. Questo nucleo
talamico a sua volta proietta alla corteccia orbito frontale, che si ritiene essere la parte di
corteccia implicata nella percezione cosciente degli odori. Percio' il sistema olfattivo ha due
diversi tipi di proiezioni, uno verso il sistema limbico (l'amigdala e l'ippocampo) e l'altra
verso il talamo ed il neocortex (integrandosi con il sistema gustativo). I nuclei dell'amigdala
sono anche centri di ritrasmissione che connettono la corteccia olfattiva all'ipotalamo e al
tegmento del mesencefalo. Si ritiene che questa via limbica sia deputata a mediare le
componenti affettive degli stimoli odoriferi. Al contrario, la proiezione talamo-neocortex e'
deputata alla percezione cosciente dell'olfatto, percio' il sistema olfattivo rappresenta un
esempio di sistema in cui viene impiegata un'analisi in parallelo.
Detlev Schild. 1988. Principles of odor coding and a neural network for odor discrimination
. Biophysical Society. Vol 54 DEc 1988 : 1001-1011
21
22
3.2 Disegno del sistema olfattivo in generale
22
23
Dopo aver descritto la struttura del sistema olfattivo nell'uomo, vediamo gli elementi che sono
comuni ai sistemi olfattivi delle varie specie animali e che utilizzeremo nella modellizzazione
analitica. Useremo in realta' una rappresentazione semplificata , per poter costruire in seguito
un modello che potra' accordarsi con una realizzazione in termini di reti neuronali.
La struttura dettagliata del sistema olfattivo differisce da specie a specie. Comunque , quando
sono considerati solo i primi stadi
dell'elaborazione dell'informazione,
tutti i sistemi
mostrano delle caratteristiche comuni. Queste caratteristiche sono di seguito esposte.
1)Il primo stadio e' composto di cellule recettrici che possono essere inserite in una mucosa,
come avviene nei vertebrati o che possono essere situate in diverse parti del corpo, come
antenne o zampe. Le cellule recettrici olfattive sono numerose in tutte le specie. (Per un
confronto si veda il riferimento[Chase, R. 1986. Lessons from snails tentacles. Chem. Sens.
11: 411-426] ). 107 cellule e' il numero medio che si trova nei vertebrati. Una tipica cellula
recettrice di vertebrato non e' molto selettiva; risultati sperimentali indicano che una cellula
e' capace di rispondere ad un grande numero di stimoli, benche' con differenti livelli di
attivita' [Revial, M. F. G. Sicard, A. Duchamp, and A. Holey. 1982. New Studies on odour
23
24
discrimination in the frog's olfactory receptor cells. I. Experimental results. Chem. Sens 7:
175-190]. A parte le collocazioni delle cellule recettrici che differiscono nelle diverse specie,
le loro connessioni nello strato successivo del sistema sono piutttosto simili.
Gli assoni delle cellule recettrici, che lasciano la mucosa, formano il nervo olfattivo che entra
nel bulbo olfattivo. Qua gli assoni terminano nei cosiddetti glomeruli che sono dei
conglomerati di forma sferica di sinapsi, poste tra gli assoni dei recettori e le cellulle mitrali a
pennacchio, i neuroni di uscita del bulbo olfattivo. L'attivazione dei glomeruli sembra che sia
omogenea, quando il sistema viene stimolato, come indicato dagli studi 2-DOG [Lancet, D.
, C. A. Greer, J. S. Kauer, and G. M. Shepherd. 1982Mapping of odor-related neuronal
activity in the olfactory bulb by 2DOG autoradiography. Proc. Natl. Acad. Sci. USA. 79: 670166] . Le cellule mitrali ricevono i loro segnali di ingresso dai glomeruli, posti nelle loro
vicinanze o per mezzo di pochi dendriti che entrano nei glomeruli adiacenti o per mezzo di un
dendrite principale che, a sua volta, deriva dal glomerulo in cui esso entra. Le cellule mitrali
possono anche ricevere input dai glomeruli limitrofi attraverso interneuroni eccitatori (come
nei vertebrati superiori ). Ortogonalmente a questa direzione del flusso di informazione dai
recettori al SNC, vi sono dei circuiti locali che incorporano cellule periglomerulari, cellule
con un corto assone e cellule granulari apportatrici di segnali inibitori ed eccitatori tra le
cellule mitrali a pennacchio. Gli interneuroni inibitori sono sotto il controllo efferente. Le
cellule a granulo, stimolate dalle cellule mitrali a pennacchio, inibiscono queste ultime nelle
loro vicinanze, cioe' esse inibiscono le cellule mitrali a pennacchio dalle quali sono state
precedentemente eccitate (facendo pensare ad una sorta di meccanismo di controllo in
retroazione negativa vedi ART). Vediamo ora cosa succede al livello di cellule recettrici. Le
cellule recettrici generano potenziali di azione ad un tasso relativamente lento, circa 0-15
spike/s. Il tasso spontaneo e' ben al disotto di 1spike/s . La risposta ad un impulso di odore a
gradino e' chiaramente uno stato dinamico, che comincia con un salto brusco, seguito da un'
attivita' approssimativamente costante . La distribuzione degli intervalli tra gli spikes e' (
eccetto che per un breve intervallo di tempo) approssimativamente esponenziale . Il verificarsi
degli spike e' causale ( ossia si ha dopo il verificarsi dell'evento gradino di odore) ed i treni di
24
25
spike, prodotti dalle cellule recettrici, sono realizzazioni di un processo di Poisson (vedi
Appendice). Gli assoni dei recettori convergono sulle cellule mitrali con all'incirca 1000
assoni per cellula mitrale, comunque, poiche' vi sono circa 25 volte piu' cellule mitrali che
glomeruli, il fattore di convergenza di un recettore sui glomeruli e' di circa 25000. Questi
numeri dimostrano chiaramente un' enorme convergenza di cellule recettrici sui glomeruli ed
una divergenza dai glomeruli verso le cellule mitrali. I numeri precisi non sono fondamentali
in questo contesto. Si considera che circa 1000 treni di spike con un tasso medio di circa uno
spike/s formano l'ingresso di una cellula mitrale. L'ingresso complessivo alla cellula mitrale e'
di circa 1000 spikes al secondo, corrispondenti ad un intervallo medio tra gli spikes di 1 ms.
Data la lunga costante di tempo eccittatoria di una cellula mitrale (circa >100 ms) , e' ovvio
che l'ingresso globale ad una cellula mitrale e' una funzione monotona che segue il corso
dinamico temporale dell' attivita' della cellula recettrice. Questi fenomeni sono stati registrati
nelle scariche delle cellule mitrali del pesce dorato da Schild stesso. [Schild, DS. 1986.System
analysis of the goldfish olfactory bulb: spatio-temporal transfer properties of the mitral cell
granule cell complex. Biol. Cybern. 54:9-19]. E' stato registrato un brusco salto dinamico,
seguito da una attivita' statica. Il brusco salto e' talora positivo come nelle cellule recettrici,
talora negativo e seguito da una depressione dell' attivita' . Questo e' esattamente cio' che ci si
potrebbe aspettare da semplici considerazioni concernenti la rete ad inibizione laterale (vedi
le mappe di Kohonen nel seguito).
Vi e' anche un altro, piu' significativo, aspetto della convergenza dalle cellule recettrici alle
cellule mitrali. Come menzionato, i treni di spike delle cellule recettrici sono in buona
approssimazione dei processi di Poisson. Poiche ' la somma di piu' processi di Poisson e'
ancora un processo di Poisson, l'ingresso totale ad una cellula mitrale e' ancora un processo di
qesto tipo. L'attivita' relativa ai treni di spike sommati e' approssimativamente proporzionale
al numero N di treni sommati. La deviazione standard del processo somma aumenta,
comunque, solo proporzionalmente a
N . In altre parole, l'ampiezza del rumore relativo
(r.m.s.) diminuisce come N-1/2.
25
26
E' chiaro che questa diminuizione del rumore incrementa la capacita' di elaborazione dell'
informazione da parte del canale " recettori --> cellule mitrali".
La descrizione precedente mette in luce caratteristiche che sono comuni ai livelli periferici di
tutti i sistemi olfattivi. Solo queste caratteristiche generali sono necessarie per la formulazione
del seguente concetto di codifica olfattiva.
Nella figura sotto riportata e' rappresentata sinteticamente la struttura olfattiva.
Fig tratta da: J. Kauer Contributions of topography and parallel processing to odor coding
in the vertebrate olfactory pathway . Trends in neuroscience, Vol 14, No. 2, 1991. pp 79-85
4.0 Un modello della codifica degli odori mediante spazi vettoriali.
26
27
4.1 Attivita' delle cellule recettrici.
La proiezione degli odori Sk, k=1, 2, . . . , K sulla attivita' di tutti i recettori puo' essere
descritta dai vettori r(Sk),
f1 : S k → r(S k ) ∈ R = R L
ove R ha una base ortornormale r1, r2, . . . rL. Qua , gli indici indicanti gli stimoli sono
scritti
come apici e quelli indicanti i componenti vettoriali
come pedici. La l-esima
componente rl di r(Sk) e' l'insieme che indica la risposta all'odore S k da parte della l-esima
classe di cellule recettrici. Una classe di cellule recettrici e' rappresentata come un insieme di
recettori che rispondono tutti quanti nello stesso modo ad un qualsiasi insieme di stimoli
successivamente applicati L 0 .
Nel caso piu' semplice che non vi siano due cellule recettrici che rispondono nella stessa
maniera agli stimoli, il numero L di classi di cellule recettrici eguaglia il numero di cellule
recettrici stesse. Il numero L e' attualmente sconosciuto, comunque esistono esperimenti che
indicano la probabile esistenza, a livello biochimico, di differenti classi di cellule recettrici .
27
28
In seguito,
i componenti vettoriali verranno trattati come variabili stazionarie; si deve
comunque tenere presente che essi sono in realta' funzioni del tempo; per l'esattezza l'insieme
rappresenta risposte di attivita' delle classi di recettori.
La dissimilarita' fisiologica D di due odori S k e S i puo' essere descritta in R come la
lunghezza D (k , i ) del vettore differenza tra rk e ri:
D = r k − ri .
Una definizione alternativa della somiglianza tra stimoli a livello di recettori e' il coefficiente
di
correlazione
C r (k , i )
tra
i
componenti
vettoriali
rlk , l = 1, 2, . . . , L
ed
rli , l = 1, 2, . . . , L dei due stimoli k e i .
L
(
)
C r (k , i )= ∑ (rlk rli ) / L = r k • r i / L
l=1
E' interessante come vengano rappresentati a livello di recettori stimoli dovuti a sostanze
aventi stessa struttura molecolare e composizione, ma differenti concentrazioni . Uno studio
indica che si verifica un reclutamento di cellule recettrici, se la concentrazione cresce. Come
conseguenza , un incremento nella concentrazione portera' sia ad un aumento nella lunghezza
sia ad un cambio di direzione di r. La concentrazione non viene cosi' solo codificata da un
incremento della frequenza con cui una singola cellula emette impulsi, ma anche dal pattern
tra le fibre delle classi di recettori attivi. I grafici indicanti come la concentrazione influenzi
le attivita' delle cellule recettrici sono mostrati nella figura sottostante.
28
29
Il vettore r , visto come una funzione della concentrazione , da' una curva r(c) nello spazio
di attivita' dei recettori R (per il caso bidimensionale, si veda la figura c). Due odori S k e S i
sono fisiologicamente distinguibili a livello dei recettori, se le curve rk (c ) e ri (c ) non si
sovrappongono ; se le curve r(c) di due odori coincidono ad una certa concentrazione, questi
odori sono codificati in modo identico a questa concentrazione. Sorprendentemente ,
comunque, questa ambiguita' a livello dei recettori puo' essere rimossa in un successivo
stadio del sistema, come vedremo in seguito. Occupiamoci ora del problema dell'invarianza
della percezione olfattiva. Nell'olfatto, c'e' un fenomeno di questo tipo. Si immagini uno
sperimentattore che misuri la risposta a due stimoli applicati in successione, diciamo A e B,
che vengono registrati simultaneamente da 4 cellule recettrici. Supponiamo che egli non
conosca ne' le qualita' ne' le concentrazioni degli stimoli, ne' da quali classi di recettori stia
registrando. Se i due stimoli risultano nella misura come due pattern che si sovrappongono
cosi' come quelli di c1 e c5 in fig. c), allora non esiste alcun modo con il quale si possa
decidere se gli stimoli abbiano o non abbiano la stessa qualita'. In altre parole,
la
concentrazione e la codifica della qualita' non sono ben separate a livello dei recettori. Stimoli
della stessa qualita' , ma di differenti concentrazioni non vengono proiettate sugli stessi
insiemi di classi di recettori. L'invarianza nella percezione di qualita' (che si mantiene per la
maggior parte degli stimoli) significa che una curva rk (c) e' vista come un tutt'uno in uno
29
30
stadio successivo nella elaborazione dell'informazione. Invece la sovrapposizione
di rk (c)
con un'altra curva in un solo punto e' solo di importanza secondaria.
4.1.1 Convergenza recettori-glomeruli.
La convergenza delle fibre delle cellule dei recettori sui glomeruli puo' venire adeguatamente
descritta dai coefficienti cln . cln indica un numero di fibre recettrici di classe l che entra
nell'n-esimo glomerulo. La proiezione f2 delle attivita' della classe dei recettori sull' attivita'
in ingresso dei glomeruli,
f2 : R → G = R N
e' data da
L
g n = ∑ c lnrl
(1)
l =1
l'ingresso a tutti i glomeruli e' allora:
g = (g1 , g2 , g3, . . . , gN )
Al livello di glomeruli, gli stimoli S k , k = 1, 2,..., K sono cosi' rappresentati in uno spazio Ndimensionale. Il numero N di glomeruli puo' andare da un minimo di 20 , come nel tentacolo
della chiocciola , a 3000 nei vertebrati superiori.
Quando i coefficienti c ln vengono scritti in forma di matrice C = (c ln ) , la relazione lineare
tra r e g e'
g = C r
o quando lo stimolo e' indicato,
30
31
g k = C rk
Ove i coefficienti c ln sono incogniti. Questo fatto comunque non ci impedisce alcuni
commenti sul loro significato:
(a) Tutti i coefficienti c ln sono positivi o zero. La trasformazione non e' percio' una rotazione
del sistema di coordinate, poiche' per questo caso sarebbero necessari anche coefficienti
negativi.
(b) I coefficienti c ln dell' ennesimo glomerulo compongono il vettore
c n = (c1n , c 2n , . . . , c Ln ) ,
e l'ingresso complessivo g n all'ennesimo glomerulo (Eq. 1) puo' essere scritto come il
prodotto scalare:
gn = cn • r
Da cui emerge che l'ingresso complessivo glomerulare e' massimo se c n ed r sono paralleli.
La convergenza recettori-glomeruli puo' cosi' essere ben visualizzata come segue: si immagini
per ciascun glomerulo un vettore di forma predeterminata c n , in modo che la forma di
convergenza possa essere visualizzata come un numero di vettori c n nello spazio delle classi
di recettori. Quando uno stimolo rappresentato da rk viene applicato, le proiezioni di rk su
tutti i vettori c n rappresentano il pattern di ingresso ai glomeruli.
Un altro modo di vedere questo fenomeno e' che la somma in eq. 1 non e' niente altro che il
coefficiente di correlazione incrociata tra le componenti dei vettori c n ed r. Con r parallelo a
c n , la correlazione e' massima e l' n-esimo glomerulo riceve l'input massimo. Altri glomeruli,
31
32
i cui vettori di convergenza non sono ortogonali a cn , ricevono input da questo stimolo rk .
Fino a questo momento, i vettori di convergenza sono stati considerati come fissi, laddove
gli stimoli erano variabili. Anche l'opposto potrebbe essere importante, data la plasticita' del
sistema. Se vi fosse uno stimolo predominante ( rk ) ed una convergenza recettore glomerulo
che possa variare nel tempo, allora l'ingresso ad un determinato glomerulo potrebbe diventare
massimo, se c ln cambiasse in modo tale che cn terminasse parallelo ad rk. In questo caso, il
vettore dell'attivita' dei recettori verrebbe mappato ottimamente su di un glomerulo ed il
glomerulo correlatore sarebbe un filtro ottimo adattato ed un riconoscitore auto-organizzato di
caratteristiche, come indicato da Kohonen [Kohonen,
T.
1987 Self-Ogranization and
Associative Memory. Springer-Verlag, Berlin]. Una analogo fenomeno di convergenza
auto-organizzata puo' essere ipotizzata, considerando piu' di un glomerulo .
(c) I numeri L e N, ed in particolare il loro rapporto L/N influenzano considerevolmente il
processo di elaborazione dell'informazione. N puo', almeno in linea di principio, venire
misurato. L e' attualmente sconosciuto, cosicche' occorre discutere tre casi: L<N, L=N, e
L>N. Nel primo caso la matrice C ha piu' righe che colonne, ed N differenti odori potrebbero
essere mappati in modo ottimo sui glomeruli. Per tutti gli altri stimoli, il vettore rk non
coincide con alcuno dei vettori di convergenza gn, e la forma in ingresso glomerulare di
questi stimoli e' una combinazione lineare degli stimoli mappati in modo ottimo. La
rappresentazione glomerulare di uno stimolo rk e' poi tanto piu' ridondante quanto piu' e'
grande il rapporto N/L. Questa ridondanza garantisce una trasmissione molto sicura
dell'informazione. Se , d'altra parte, gli spazi R e G hanno lo stesso numero di dimensioni
(L=N), i vettori rk vengono semplicemente trasformati in un altro sistema di coordinate; la
mappatura puo' venir descritta come sopra "mappatura ottimale ", ma vi sono anche altre
ragionevoli ipotesi. Se le connessioni tra i glomeruli e gli strati seguenti del sistema operano
come una memoria associativa [si veda le reti neurali BAM], il che sembra la cosa piu'
plausibile, l'intera informazione odorifica potrebbe venire recuperata da solo una parte del
32
33
sistema anche nel caso di L=N. Esperimenti condotti da Schild stesso non sembrano fornire
nessuna indicazione circa la dimensione di L.
Se e' L>N, e' importante conoscere il numero L 0 di vettori rk linearmente indipendenti. E'
possibile che vi siano meno ( L 0 <L ) vettori di attivita' dei recettori indipendenti, di quanto
non vi siano classi di recettori ( un semplice esempio di un tale caso sarebbe geometricamente
rappresentato da un certo numero di vettori su di un piano di orientazione arbitraria in un
sistema di coordinate tridimensionali). Se L0>N, allora
l'informazione andrebbe persa. In
questo caso differenti odori potrebbero avere rappresentazioni identiche. Nel caso opposto
(L0 < N), la trasformazione C potrebbe ridurre la dimensione dello spazio in cui gli odori
sono rappresentati.
(d) Una leggera variazione di uno stimolo (r-->r+dr) risulterebbe in una proporzionale
variazione del vettore g a livello glomerulare. In altre parole, quali che siano i coefficienti
della matrice C, stimoli simili sono mappati su forme di attivita' similari in ingresso ai
glomeruli . Vedremo meglio in seguito cosa questo fatto significhi.
4.1.2 Proiezione glomeruli-cellule mitrali
La mappatura dai glomeruli agli ingressi delle cellule mitrali e' data da
f3 : G → M = R J
con mkj = ∑n=1 p g kn
nj
N
o con la matrice scritta come P = (pnj ),
mk = P g k
Poiche vi sono circa 25 volte piu' cellule mitrali che glomeruli, J ≈ 25N . La matrice P ha
proprieta' speciali: essa e' una matrice di proiezione con elementi positivi che non tendono ad
33
34
annullarsi su e nell'intorno della diagonale. Questa proprieta' corrisponde ad una divergenza
localmente ristretta dai glomeruli alle cellule mitrali. L'informazione di S k ,
rk e g k
e'
decomposta e proiettata su differenti sottospazi di M corrispondenti a differenti aree dello
strato di cellule mitrali . Poiche' gli odori sono rappresentati in al massimo N dimensioni (la
massima dimensione dello spazio G), non possono esservi piu' di N vettori linearmente
indipendenti in ingresso alle cellule mitrali . Assumendo 25 volte piu' cellule mitrali che
glomeruli, questo significa che 24*N (ossia 25*N - N) cellule mitrali, cioe' la maggior parte
di esse, hanno vettori in ingresso che sono linearmente dipendenti e principalmente non
necessari per la convergenza dell'informazione di uno specifico odore. Comunque ,
la
combinazione delle cellule coinvolte nella trasmissione delle informazioni cambia da stimolo
a stimolo. Una particolare cellula mitrale, per esempio , la j-esima riceve i suoi ingressi
secondo il vettore:
pj = (p1j , p2 j , . . . pnj )
ove pnj
si annulla al di fuori di un certo intorno della j-esima cellula mitrale,
pj = (0, . . . , 0, pn − 1, j , pn, j , . . . pn + 1, j , 0, . . . , 0).
Una cellula mitrale adiacente riceve il suo ingresso da (quasi) gli stessi glomeruli, benche'
con differenti pesi
pj + 1 = (0, . . . , 0, pn − 1, j + 1 , pn, j + 1 , . . . pn + 1, j + 1 , 0, . . . , 0).
Le attivita' di ingresso di queste cellule sono mj = pj • g e mj + 1 = pj + 1 • g . La cellula
con il vettore proiezione p che corrisponde meglio a g riceve l'input maggiore. Estendendo
questo ragionamento a tutte le cellule mitrali, vediamo che vi sono insiemi di cellule mitrali
34
35
che vengono attivate in modo ottimo, un insieme per ciascuna forma di attivazione dei
glomeruli.
4.1.3 Inibizione laterale.
Nel bulbo olfattivo , l'inibizione laterale e' presente principalmente tra i glomeruli adiacenti e
tra le cellule mitrali. Le esatte forme di connessione cosi' come le intensita' sinaptiche sono,
comunque, non note in dettaglio. Secondo Schild e' attualmente impossibile giustificare con
precisione gli effetti di questi circuiti con inibizione laterale (con i modelli di reti neuronali da
noi impiegati tenteremo di dare una possibile spiegazione.A tale proposito si veda la CPN,
ART e le reti Hamming nel seguito della tesi). Schild ipotizza che la inibizione laterale agisca
come un filtro spaziale passa alto, che incrementi il contrasto di una forma di attivita' spaziale
. Altri possibili effetti come il guadagno dipendente dalla attivita' (vedi ART) non vengono
considerati nel modello di Schild. Quest'ultimo suppone che l'input delle cellule mitrali sia
trasformato in uscita della cellula mitrale mediante la relazione:
f4 : M → M
con,
J
bki = ∑ s jimkj
o
j=1
bk = Sm k ,
(2)
ove la (soppressione) matrice S ha valori negativi in prossimita' della diagonale e valori che
tendono ad annullarsi altrove. Le attivita' negative bki vengono scartate come non significative
per la generazione dello spike:
35
36
aki = max(0, bki ) .
(3)
Il vettore ak fornisce una approssimazione grossolana delle attivita' delle cellule mitrali
dovute agli stimoli S k . Una forte soppressione laterale puo' inibire tutte le attivita', eccetto
una entro un certo sottospazio; in questo modo,
un vettore, che prima dell'azione
dell'inibizione laterale aveva diverse componenti positive in un sottospazio, viene trasformato
in un vettore che ha solo una componente positiva. Questo effetto di ortogonalizzazione e'
riportato dalla equazione (2)
e dalla trasformazione non lineare (eq.
3). Inoltre tale
trasformazione agisce separatamente su molte aree del bulbo olfattivo piuttosto che su tutte le
cellule mitrali assieme.
4.2 Una descrizione alternativa: immagini di attivita'
Schild propone anche un modello alternativo della codifica degli odori. Nel seguito verra'
brevemente descritta una rappresentazione equivalente alternativa che tiene esplicitamente in
conto la posizione geometrica dei glomeruli e delle cellule mitrali nei rispettivi strati. La
posizione specifica di una cellula su di uno strato viene opportunamente data in coordinate
sferiche (scritte come α , β o φ , δ ) prendendo come origine il punto di mezzo del bulbo.
4.2.1 Immagine della attivita' glomerulare
Si consideri un campo vettoriale discreto in modo che per ciascun glomerulo sia assegnato un
vettore (il vettore "convergenza") c( α , β ) . La l-esima componente c l (α, β) di tale vettore
rappresenta il numero di fibre dell'l-esima classe di recettori che entra nel glomerulo alle
coordinate ( α, β ). Allora prendendo lo stimolo S k , rk si proietta sui glomeruli secondo la
relazione :
36
37
g(α, β) = c(α, β) • rk
valida per ciascuna coppia α, β . La principale differenza dalla precedente descrizione e' che
g(α, β) dipende da due coordinate geometriche, invece di essere un vettore componente. Le
componenti di c(α, β) e c n sono identiche per lo stesso glomerulo .
rk viene mappato su di un array bidimensionale g(α, β) , che puo' venire interpretato come
un' immagine spaziale dell'odore, costitutita da punti (i glomeruli), a ciascuno dei quali e'
assegnato un valore, (corrispondente ad un livello di grigio in una immagine classica)
ottenendo una descrizione simile a quella che puo' essere fatta per le immagini retiniche ( vedi
ART, BAM, HAMMING etc.
e per un paragone neurofisiologico [G.
M.
Sheperd I
microcircuiti del sistema nervoso Le Scienze Aprile 1978 No. 116 pp. 71-78]). La succesiva
elaborazione di questa immagine puo' cosi' venire compresa in termini di elaborazione digitale
di immagine e di filtraggio digitale.
4.2.2 Ingresso alla cellula mitrale
La proiezione glomerulo cellula mitrale e' ristretta ad una certa zona ed e' data da :
m(φ, δ) = ∑ pα,β (φ, δ)g(α, β)
α, β
con pα,β (φ, δ ) = 0 per α − φ ≥ Δδ e β − δ ≥ Δδ e pα,β (φ, δ ) ≥ 0 altrimenti.
I valori Δφ e Δδ sono determinati dal range spaziale di ingresso di una cellula mitrale (
comprendendo gli interneuroni eccitatori). I coefficienti del filtro pα,β (φ, δ ) determinano il
tipo di filtro: essi sono zero per ogni φ, δ eccettuata in una piccola regione del piano α, β .
I filtri cosi' descritti sono filtri spaziali passa basso.
Se i coefficienti dei filtri fossero spazio-invarianti
l'equazione (3) diverrebbe
37
cioe' , pα,β (φ, δ ) = p(α − φ, β − δ ),
38
m(φ , δ ) = ∑ p(α − φ , β − δ )g (α , β )
α ,β
ove la somma e' effettuata solo per piccoli intervalli di α − φ e β − δ .
Questa espressione ha la forma di una convoluzione, ed il carattere passa basso e' meno
ovvio. Comunque potrebbe essere che per ciascuna cellula mitrale i coefficienti del filtro
siano differrenti, cioe' i coefficienti non sono spazio-invarianti. In ogni caso, ogni cellula
mitrale agisce come un filtro passa basso sulla immagine glomerulare, campionando ed
elaborando solo una piccola parte di esso. L'immagine glomerulare viene cosi' filtrata da un
filtro passa basso composto da un numero di cellule mitrali 25 volte piu' grande di quanto non
siano i glomeruli. Cellule mitrali adiacenti,
che ricevono ingresso da (quasi) lo stesso
insieme di glomeruli, hanno ingressi simili che differiscono solo per il fatto di essere
differenti combinazioni lineari dei coefficienti del filtro in ingresso.
4.2.3 Immagine dell'uscita della cellula mitrale.
La funzione in ingresso alla cellula mitrale mk viene trasformata in una funzione in uscita
dalla cellula mitrale bk mediante la relazione:
bk (φ, δ) = ∑ sα,β (φ, δ)mk (α, β)
α, β
Di nuovo, i valori negativi sono scartati : ak (φ, δ) = max 0, bk (φ, δ) .
ak e' una funzione che assegna alla cellula mitrale di coordinate (φ, δ ) l'attivita' ak (φ, δ ) .
ak e' cosi' la forma di attivita' spaziale delle cellule mitrali nello strato bidimensionale delle
cellule mitrali stesse.
4.2.4 Due tipi di discriminazione nel sistema olfattivo(ordine spaziale e inibizione
laterale)
38
39
(vedi CPN)
Dalle considerazioni fatte nei paragrafi precedenti e' chiaro che odori che sono simili nello
spazio di attivita' delle cellule recettrici vengono mappati su pattern spazialmente adiacenti di
glomeruli e sulle cellule mitrali con attivita' similari . Percio' e' stata introdotta una relazione
di ordine spaziale (vedi mappe di Kohonen) tra gli odori . L'inibizione laterale a livello di
cellula mitrale ha cosi' l'effetto di introdurre un meccanismo di competizione che ha lo scopo
di incrementare il contrasto tra le rappresentazioni neurali degli odori. Odori simili possono
cosi' essere ben discriminati a livello di cellula mitrale.
4.2.5 Riconoscimento qualitativo e memoria associativa
Non esiste alcuna prova sperimentale che possa spiegare come il sistema olfattivo possa
associare la stessa qualita' con i differenti insiemi di cellule mitrali attivate da diverse
concentrazioni. Anche da osservazioni tratte dalle reti neuronali, si puo' ipotizzare che la
percezione invariante della qualita' di un odore potrebbe essere dovuta ad un meccanismo di
memoria associativa (tipo quello alla base delle reti BAM piu' avanti descritte) che si
manifesterebbe entro il bulbo olfattivo. Come in tutti i casi di memoria associativa, il
manifestarsi di una informazione parziale in ingresso (uno stimolo dovuto ad un odore a
concentrazione specifica ) provoca un recupero totale dell'informazione (la forma di qualita'
associata con tutti i valori di concentrazione.
4.2.6 Costruzione di un sensore di odori intelligente artificiale.
La costruzione di un sensore di odori intelligente artificiale, corrispondente ad un sistema
biologico, pone un problema pratico: come dovrebbero essere realizzate artificialmente le
connessioni tra i tre principali tipi di elementi (cellule recettrici, glomeruli e cellule mitrali) ?
Attualmente queste connessioni non sono note nel dettaglio; e' percio' impossibile copiarle.
Mediante le reti neuronali, pero', si puo' tentare di riprodurre i meccanismi che portano ai
39
40
comportamenti esaminati nel sistema olfattivo biologico (riconoscimento e codifica degli
odori) e, in seguito, a partire dal modello cosi' costruito, andare a verificarlo mediante un
confronto parallelo tra il sistema artificiale e quello neurofisiologico. Questo e' proprio cio'
che tenteremo di fare nella nostra tesi.
40
41
Bibliografia.
[1] Murray L.
1993. pp. 9-26
Barr.
Il sistema nervoso umano. Piccin
[2] Sir John Eccles 1965. La sinapsi. Le Scienze Marzo
1965. pp 59-68
[3] P. Strata, Fisiologia del neurone E. M. S. I.
1983 pp. 10-20
Roma
[4]
J. Singh. Linguaggio e cibernetica. EST Mondadori.
1976. pp152-153
[5] Tarun Khanna "fondamenti
Wesley 1991. pp 144c-156
di
reti
neurali.
Addison
[6] A. R. Luria. L'organizzazione funzionale cerebrale.
Le Scienze giugno 1970
[7] Luigi Cattaneo. Anatomia del sistema nervoso centrale
e periferico dell'uomo. Monduzzi editore 1984.
[8] Olton David S.
ottobre 1977
La memoria spaziale . Le scienze 110 ,
[9] Amoore, J. . E. , Johnson, W. , Jr, and Rubin, M.
1964.
The stereochemical theory of odor.
Sci.
Am.
210(2): 42-49
[10] Shepherd,
olfactory bulb.
1972, Synaptic organization of mammalian
Physiol. Rev. 52-864-917
[11]
Kandel, E.
R.
& Schwartz,
J.
H.
(1982)
Principles of Neural Science.
New York:
Elsevier/North
Holland Capitolo 32 I sensi chimici:
gusto e olfatto.
[12] Evor L. Hines,
Claire C. Gianna and J. W Gardner.
Neural network based electronic nose using constructive
algorithms.
Techniques
and
application
of
neural
networks. 1993 Ellis Horwod Workshop. pp. 135-154
[13] S. G. Shirley and Krishna C.
Persaud.
biochemistry of vertebrate olfaction and taste.
neurosciences. Vol II 1990: pp 59-68
[14] Eleonora P. Giorgi.
Il
Scienze num. 101. Gennaio 1977
41
senso
dell'olfatto.
The
The
Le
42
[15] Getchell, T. V. ,
and Sheperd, G. M.
1978.
Responses of olfactory receptor cells to step pulses of
odour at different concentrations in the salamander.
J.
Physiol. (Lond. ) 282: 521-540.
[16] Stewart, W. B. , Kauer, J. S. , and Shepherd, G,
M 1979 Functional organization of rat olfactory bulb
analysed by the 2-deoxyglucose method. J. Comp.
Neurol.
185: 715-734
[17]
Kandel, E.
R.
& Schwartz,
J.
H.
(1982)
Principles of Neural Science.
New York:
Elsevier/North
Holland Capitolo 32 I sensi chimici:
gusto e olfatto.
[18] Detlev Schild. 1988. Principles of odor coding and a
neural network for odor discrimination . Biophysical
Society. Vol 54 DEc 1988 : 1001-1011
[19] Chase,
R. 1986.
Chem. Sens. 11: 411-426
Lessons
from
snails
tentacles.
[20] Revial,
M.
F.
G. Sicard,
A. Duchamp,
and A.
Holey. 1982. New Studies on odour discrimination in the
frog's olfactory receptor cells. I. Experimental results.
Chem. Sens 7: 175-190.
[21] Lancet, D. , C. A. Greer, J. S. Kauer, and G.
M.
Shepherd. 1982 Mapping of odor-related neuronal
activity in the olfactory bulb by 2-DOG autoradiography.
Proc. Natl. Acad. Sci. USA. 79: 670-166
[22] Schild, DS. 1986. System analysis of the goldfish
olfactory bulb: spatio-temporal transfer properties of the
mitral cell granule cell complex. Biol. Cybern. 54:9-19.
[23] J.
Kauer. Contributions of topography and parallel
processing to odor coding in the vertebrate olfactory
pathway . Trends in neuroscience, Vol 14, No. 2, 1991.
pp 79-85
[24] Kohonen, T. 1987 Self-Ogranization and Associative
Memory. Springer-Verlag, Berlin.
[25] G. M. Sheperd. I microcircuiti del sistema nervoso
Le Scienze Aprile 1978 No. 116 pp. 71-78
[26] J. W. Gardner & P. N. Bartlett .Pattern Recognition
in odour sensing.Academic Press.1992. pp. 161-179
[27] Richard P.
aprile 1987
Lippmann IEEE ASSP magazine. Vol 4 No.
42
2
43
[28]
T.
Kohonen
.
"Self
organized
formation
of
topologically correct feature maps" Biological Cybernetics
43: 59-69 1982 Springer-Verlag.
[29] James M.
Bower.
Relations between the dynamical
properties of single cells and their networks in piriform
(olfactory) cortex
[30] E. I.
Knudsen,
S.
du Lac,
and S. D.
Esterly
(1987) "Computational maps in the brain", Annual Review of
Neuroscience, 1987 10: 41-65.
[31] Yves Burnod.
An Adaptive neural network:
cerebral cortex 1990, Prentice Hall London
the
[32]
Mountcastle,
1957
Modality
and
topographic
properties of single neurons of cat's somatic sensory
cortex. J. Neurophys. 20, 408-434;
[33] Towe, A. Notes on the Hypothesis of columnar
organization in somatosensory cerebral cortex.
Brain
Behav. Evol. 11, 16-47 , 1975.
[34] Hebb,
Wiley 1949.
D. :
Organization of behavior. New York:
[35] Singer ,
W.
Rauschecker, J. ,
Werth,
R. : The
effect of monocular exposure to temporal contrast on
ocular dominance in kittens.
Brain Res.
134,
568-572
. 1977.
[36] R. R Llinas.
La corteccia
Scienze No. 81 Maggio 1975
del
cervelletto.
[37] Robert Hecht-Nielsen, 1 Dec 1987 /Vol.
Applied Optics p. 4979-4984
Le
26, No. 23 /
[38] Carpenter, G.A. e Grossberg, S., Neural dynamics of
category
learning
and
recognition:Attention,
memory
consolidatio, and amnesia. J.Davis, (Eds) AAAS Symposium
series, in press, 1987
[39] Robert Hcht-Nielsen Hecht. Nielsen Neurocomputer
Corporation Dec. 1987 Applications of Counterpropagation
Networks Neural Networks Vol I pp. 131-139, 1988.
[40] Sklansky ,J., .Pattern recognition: introduction and
foundations, Electrical engineering and computer science,
Dowden, Hutchinson & Ross, Inc.
43
44
[41] Gail A. Carpenter and Stephen Grossberg. A massively
parallel architecture for a self-organizing neural pattern
recognition machine. Computer Vision, Graphics and image
processing, 1987,37, 54-115
[42] S. Grossberg. How does a brain build a cognitive
code? Psychological Review 87: 1-51
[43] Freeman W. J. Mass action in the nervous system New
York : Academic Press 1975
[44] Grossberg,
S. Adaptive Pattern Classification and
universal recording II Feedback expectation,
olfaction
and illusions.Biological Cybernetics 1976, 23,121-134
[45] S.Grossberg Communication, memory, and development
.In Rosen & F. Snell, Progress in theretical biology
(vol.5).New York:Academic Press, 1978
[46] Stent, G.S., A Physiological mechanism for hebbs
postulate of learning, Procedings of the National Academy
of Sciences, U.S.A., 70, 997-1001 (1973)
[47] Bart Kosko. Contructing an Associative Memory Byte
September 1987 pp. 137-144.
[48] Grossberg, S The Adaptive Brain,
North-Holland, 1987.
I & II. Amsterdam:
[49] Kosko, B. "Bidirectional Associative Memories" IEEE
Transactions on System,
Man,
and Cybernetics,
Fall
1987.
[50] J. W. Gardner and P.N. Bartlett. A brief history of
electronic noses. Sensors and Actuators B, 18-19 (1994)
211-220.
[51] Kuniko Fukushima, "A neural network for visual
pattern recognition" Computer, March 1988,pp. 65-74
[52] Robert Hecht -Nielsen. "Neurocomputing:picking
Human brain", IEEE Spectrum, March 1988, pp. 36-41
the
[53] R. Walker, F. Favata. "Neuroinformatica", Informatica
Oggi, n.45, 1988, pp. 66-83
[54]
T.
Kohonen
typewriter",Computer ,
44
,
"
The
neural
March 1988, pp. 11-25.
phonetic
45
[55] Gail. A. Carpenter and Stephen Grossberg, "The art of
adaptive pattern recognition by a self-organizing neural
network" Computer March 1988, pp. 77-91
[56] B. Widrow, R. Winter. Neural Network for adaptive
filtering and adaptive pattern recognition, Computer March
1988. pp. 25-39
[57] R. Linsker, Self-organization in
network, Computer, March 1988, pp. 105-117
a
perceptual
[58] D. A. Turner. Conductance transient onto dendritic
Spines in a segmental cable model of hippocampal neurons,
Biophys J. Biophysical society, Vol. 46, July 1984, pp.
85-96
[59]
S. T. Welstead "Neural network and Fuzzy logic
application in c/c++, Wiley and sons , 1994, pp. 344-391
[60] C. C. Casey Klimasauskas, An introduction to neural
computing, Manuale del programma neural-works professional
II, 1988
[61] D. W. Tank, "Circuiti elettronici basati su modelli
biologici, Le Scienze n. 234 , Febbraio 1988
[62] D. L. Alkon. " I meccanismi molecolari
memoria", Le scienze n. 253, Settembre 1989
della
[64] G. E. Hinton, D. C. Plaut e Tim Shallice " La
simulazione dei danni cerebrali" Le scienze
n. 304,
Dicembre 1993
[65] W. S. McCulloch, " Embodiments of
Press,Cambridge Massachusetts, 1965,1989.
MInd"
MIT
[66] F. Crick " Function of Thalamic reticular comlplex:
the serchlight hypothesys", Procedings of national academy
of sciences 81:4586-4590.
[67] A. G. Knapp and James Anderson, "Theory of
categorization based on distributed memory storage, "
Journal of experimental psychology:learning, memory and
cognition, 10:616-637.
45