Le molecole biologiche
Le proteine
Le proteine sono macromolecole che costituiscono la
maggior parte delle strutture cellulari ed extra-cellulari
Così come nel caso di lipidi e carboidrati, anch’esse sono
formate dalla polimerizzazione di molecole più semplici
Le proteine sono caratterizzate da una grande
complessità strutturale, che da vita ad una moltitudine di
funzioni.
In base a queste funzioni, le proteine vengono
generalmente classificate
•
Proteine strutturali
Gli enzimi
•
Proteine di membrana
•
•
Fattori di trascrizione
•
Etc…
Le proteine sono polimeri lineari di piccole molecole, chiamate amminoacidi,
legate fra loro da un legame chimico detto legame peptidico
Gli amminoacidi uniti fra loro da tale legame formano la catena polipeptidica,
che diviene proteina solo quando la sua struttura tridimensionale è stabile e
funzionale (un muro NON è una casa!)
Tutte le proteine sono formate dalla diversa combinazione di soli 20
amminoacidi
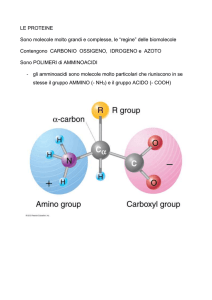
Gli amminoacidi
Molecole anfotere
Gruppo basico – gruppo
amminico
Gruppo acido Carbossile
Carbonio alpha
Ha le sue 4 valenze
legate a 4 atomi/gruppi
di atomi differenti, ciò le
rende asimmetrico e
porta alla formazione di
due isomeri ottici il D ed L
Parte variabile, catena laterale, residuo,
radicale, gruppo R
Il gruppo R differenzia gli amminoacidi e
concede loro le proprietà fisico-chimiche
Spiccata idrofobia, proteine membrana
Spiccata idrofilia
Proteine del nucleo
Istoni, legati al DNA
Alcuni amminoacidi oltre a fungere da precursori delle proteine, sono anche
precursori di alcuni fra i più importanti neurotrasmettitori o sono neurotrasmettitori
loro stessi
In particolare:
Serotonina
Dopamina
Noradrenalina
Adrenalina
GABA
Il legame peptidico
Il legame peptidico è un legame covalente, molto resistente che si origina fra il
gruppo amminico di un amminoacido ed il gruppo carbossilico
dell’amminoacido che lo precede, con l’eliminazione di una molecola
d’acqua.
Nel legame peptidico C-N sono molto più vicini che in una qualsiasi altra
tipologia di legame C-N, ciò rende il legame molto robusto e gli concede un
carattere parziale di doppio legame, eliminando funzioni rotazionali.
Tuttavia un legame così stabile può creare problemi per l’assorbimento dei
singoli amminoacidi, per tale ragione gli organismi sono dotati di proteasi come
la pepsina e la tripsina (proteine enzimatiche) che tagliano il legame peptidico
alle normali condizioni di temperatura
Il legame peptidico forma i polipeptidi
Terminale amminico
INIZIO
Terminale
carbossilico
FINE
Il risultato sarà una lunga catena flessibile il cui scheletro assile è composto
dalla successione di atomi N-C-C-N-C-C-N…etc…
Lo scheletro assile è a sua volta coperto dai vari gruppi R di ogni amminoacido
che sporgono più o meno a seconda della loro lunghezza e che concedono
alla catena le sue proprietà fisico-chimiche
La sequenza degli amminoacidi che compone una catena
polipeptidica è definita come la struttura primaria della proteina.
Le strutture secondarie: alpha-eliche
Anche se i legami peptidici sono non-rotazionali, tutti gli altri
legami presenti nello scheletro assile, lo sono. Ciò consente
alla catena di piegarsi in questi punti sotto la spinta
dell’agitazione termica.
Questo porta alla formazione di una configurazione simile ad
un’elica bloccata da legami idrogeno che si instaurano fra
gli H-N e gli O-C liberi di ogni amminoacido.
Questi legami avvengono con una certa regolarità, i.e. ogni
4 amminoacidi.
Tale conformazione a cilindro, prende il nome di alpha-elica
ed è rivestita dai vari R di ogni amminoacido, in particolare
ogni giro è coperto da 3,6 R
Le strutture secondarie: i foglietti beta-planari
In modo del tutto
analogo i legami
idrogeno possono anche
legare fra loro due
bracci della stessa
catena, o due catane
appaiate e parallele,
dando alla struttura una
forma laminare
Denominata
Foglietti Beta-planari
I bracci che formano i
foglietti possono avere
un andamento parallelo,
anti-parallelo o misto
I «motivi» delle proteine
Combinazioni di alpha-eliche
Elica-ansa-elica
Chiusura lampo di
leucine
I «motivi» delle proteine
Combinazioni di foglietti-beta
Beta-barile
Beta-sandwich
Le strutture terziarie
Insieme degli avvolgimenti
della catena polipeptidica,
stabilizzati dalla formazione
di legami dei gruppi R
E’ stato dimostrato che l’acquisizione delle strutture secondarie e terziarie,
dipendono esclusivamente dalla struttura primaria di una proteina.
La metodologia per giungere a tali conclusioni consiste nella denaturazione di
una proteina nativa (scioglimento di tutti i legami secondari e terziari) e nella
successiva rinaturazione
Le strutture quaternarie
La struttura quaternaria non è presente in tutte le proteine, ma è tipica delle
proteine multimeriche, cioè costituite da più catene peptidiche o subunità, che
possono essere uguali o diverse fra loro.
Un tipico esempio di proteina con struttura quaternaria è l’emoglobina, ovvero la
proteina che si occupa di trasportare ossigeno, contenuta nei nostri globuli rossi.
Essa è costituita da due diversi tipi di catene peptidiche (alpha e beta).
Inoltre ogni catena contiene al suo interno una molecola non-proteica detta gruppo
eme, con un atomo di ferro che è lo specifico legame dell’ossigeno.
La struttura quaternaria rappresenta un importante punto di arrivo del processo
dell’evoluzione molecolare poiché consente un effetto cooperativo fra le diverse unità
Amminoacidi
Catene polipeptidiche, alphaeliche e foglietti-beta
«motivi» : proteine = gotico : architettura
Struttura terziaria
Proteina
Le modificazioni post-traduzionali
Dopo la sintesi e la completa acquisizione dei livelli strutturali secondario e
terziario, la maggior parte delle proteine va incontro a particolari modifiche
strutturali dette modifcazioni post-traduzionali.
Alcune di queste modificazioni sono permanenti e necessarie affinché la
proteina possa acquisire la conformazione tridimensionale finale e quindi le sue
funzionalità biologiche
Altre modificazioni sono reversibili e svolgono funzioni importanti nel controllo
della proteina stessa.
Le modificazioni post-traduzionali
Le modificazioni permanenti si possono classificare in:
•
•
Tagli proteolitici (eseguiti da enzimi specifici detti proteasi)
• Distruggere proteine difettose
•
Eliminare porzioni della catena polipeptidiche non più necessarie (rimuovere porzioni della
catena che inizialmente possono essere necessari per il corretto avvolgimento della
proteina, ma che in seguito possono rivelarsi superflui) – L’esempio dell’INSULINA
•
Proteggere se stesse dalla attività potenzialmente pericolosa della proteina matura (ad
esempio nel caso delle cellule che creano gli enzimi in grado di scindere le proteine assunte
col cibo e che devono proteggersi dagli effetti di proteasi dei loro stessi prodotti) – proenzimi inattivi
•
Tagliare una sola macro-catena polipeptidica in vari frammenti, ognuno dei quali
costituisce una proteina avente una sua propria funzione – L’esempio del pro-opiomelanocortina (POMC)
L’aggiunta di porzioni non proteiche
• Oligosaccaridi - Glicoproteine
• Lipidi - Lipoproteine
• Gruppo prostetico
•
Eliminare porzioni della catena polipeptidiche non più necessarie (rimuovere porzioni della catena che
inizialmente possono essere necessari per il corretto avvolgimento della proteina, ma che in seguito
possono rivelarsi superflui) – L’esempio dell’INSULINA
Le modificazioni post-traduzionali
Le modificazioni permanenti si possono classificare in:
•
•
Tagli proteolitici (eseguiti da enzimi specifici detti proteasi)
• Distruggere proteine difettose
•
Eliminare porzioni della catena polipeptidiche non più necessarie (rimuovere porzioni della
catena che inizialmente possono essere necessari per il corretto avvolgimento della
proteina, ma che in seguito possono rivelarsi superflui) – L’esempio dell’INSULINA
•
Proteggere se stesse dalla attività potenzialmente pericolosa della proteina matura (ad
esempio nel caso delle cellule che creano gli enzimi in grado di scindere le proteine assunte
col cibo e che devono proteggersi dagli effetti di proteasi dei loro stessi prodotti) – proenzimi inattivi
•
Tagliare una sola macro-catena polipeptidica in vari frammenti, ognuno dei quali
costituisce una proteina avente una sua propria funzione – L’esempio del pro-opiomelanocortina (POMC)
L’aggiunta di porzioni non proteiche
• Oligosaccaridi - Glicoproteine
• Lipidi - Lipoproteine
• Gruppo prostetico
•
Tagliare una sola macro-catena polipeptidica in vari frammenti, ognuno dei quali costituisce una
proteina avente una sua propria funzione – L’esempio del pro-opiomelano-cortina (POMC), prodotto
da molti distretti corporei ma dei quali il più importante è senza dubbio l’ipofisi
Corticotropina, stimola la
produzione dei
corticosteroidi nella
ghiandola surrenale
Regolazione del
comportamento
Grooming nei topi
Ulteriori tagli che producono corte
molecole di 5 amminoacidi dette
encefaline
Oppioidi endogeni
Le modificazioni post-traduzionali
Le modificazioni permanenti si possono classificare in:
•
•
Tagli proteolitici (eseguiti da enzimi specifici detti proteasi)
• Distruggere proteine difettose
•
Eliminare porzioni della catena polipeptidiche non più necessarie (rimuovere porzioni della
catena che inizialmente possono essere necessari per il corretto avvolgimento della
proteina, ma che in seguito possono rivelarsi superflui) – L’esempio dell’INSULINA
•
Proteggere se stesse dalla attività potenzialmente pericolosa della proteina matura (ad
esempio nel caso delle cellule che creano gli enzimi in grado di scindere le proteine assunte
col cibo e che devono proteggersi dagli effetti di proteasi dei loro stessi prodotti) – proenzimi inattivi
•
Tagliare una sola macro-catena polipeptidica in vari frammenti, ognuno dei quali
costituisce una proteina avente una sua propria funzione – L’esempio del pro-opiomelanocortina (POMC)
L’aggiunta di porzioni non proteiche
• Oligosaccaridi - Glicoproteine
• Lipidi - Lipoproteine
• Gruppo prostetico – gruppo eme
Le modificazioni post-traduzionali
Le modificazioni reversibili constano nei processi di fosforilazione e defosforilazione, aggiunta o rimozione di un gruppo fosfato al gruppo R di uno o più
amminoacidi alcolici (serina, treonina e tirosina)
Proteinchinasi
I complessi sopramolecolari
Spesso accade che le proteine non svolgono le loro funzioni singolarmente ma in unione
con altre proteine uguali o diverse.
L’insieme di queste proteine formano i complessi sopramolecolari.
Questi sono riconoscibili per la presenza di numerosi «motivi» e sono tenute insieme da
interazioni deboli o legami covalenti.
Ogni proteina di un complesso sopramolecolare, viene definita sub-unità.
Ciascuna sub-unità può svolgere compiti diversi.
Vedere Fig. 2.35