Negli organismi unicellulari ogni cellula agisce e si divide
indipendentemente dalle altre. Il più importante fattore
limitante la sua indipendenza è rappresentato dalla
quantità di nutrienti presenti nel mezzo.
Esistono infatti precisi momenti, collegati a carenza di nutrienti, in cui queste
cellule comunicano fra loro e si coordinano. Questo avviene quando, ad
esempio, nel lievito (Saccharomyces cerevisiae), si differenziano le due
tipologie cellulari complementari (mating type) che si accoppieranno.
Quando mancano i nutrienti, l’ameba della specie Dictyostelium
discoideum mette in atto un meccanismo di aggregazione cellulare
con formazione di un organismo pluricellulare semplice, ma capace
di mantenere e proteggere spore.
Finché è presente nutrimento, queste amebe vivono
come organismi unicellulari. Se questo viene a
mancare, le cellule si dirigono verso un centro comune
(a) dove si aggregano e formano una specie di
mucillagine (b) da cui si allunga un peduncolo (c).
(a)
(c)
(b)
Dal momento in cui le cellule si
aggregano e formano il pluricellulare
nelle diverse regioni si
differenziano tessuti
specifici. Nel peduncolo
si forma un tessuto
di sostegno e all’apice si forma
un corpo fruttifero con le spore.
Come è stato dimostrato
sperimentalmente, il segnale
che induce l’aggregazione e
che viene emesso quando
le cellule vengono
affamate è l’AMP ciclico.
Slime Molds Time Lapse - YouTube
“…no more of a bag of amoebae
encased in a thin slime sheath..”
• La pluricellularità implica che le cellule stiano in comunicazione le une con
le altre in un sistema di rete complesso che coordina tutte le attività, dal
differenziamento al metabolismo. Le cellule comunicano con segnali
(molecole segnalanti) che partono da una cellula segnalante ed arrivano
alla cellula target che ha i recettori per ricevere tale segnalazione. Per
segnalare possono essere usate una gran varietà di sostanze come peptidi,
proteine, piccole molecole lipofiliche o derivati da amminoacidi.
• Quando la segnalazione arriva alla superficie della cellula esiste un
complesso meccanismo che converte la segnalazione esterna in una
segnalazione interna. Tale meccanismo si chiama trasduzione del segnale.
FORMAZIONE DEI TESSUTI
Come le cellule si integrano per formare tessuti: molecole di adesione,
giunzioni cellulari e matrice extracellulare
CAM: Cell Adhesion Molecules, molecole di adesione cellulare.
Sono proteine integrali di membrana che permettono alle
cellule di aderire e interagire tra loro o con la matrice
extracellulare.
ECM: Extracellular Cell Matrix, matrice extracellulare. Complessa
rete di proteine e carboidrati secreti dalle cellule aventi un ruolo
strutturale (divisione e movimento) e funzionale.
Giunzioni: raggruppamenti di CAM.
Proteine del citoscheletro: microfilamenti di actina e
filamenti intermedi.
Come si presenta l’interfaccia matrice-cellula al microscopio elettronico
PRINCIPALI FAMIGLIE DI MOLECOLE DI ADESIONE
Le N-CAM (CAM della superfamiglia delle immunoglobuline) e le proteine dimeriche E-caderine di
una cellula formano legami omofilici con molecole presenti sulla membrana della cellula
adiacente. Le selectine, anch'esse proteine dimeriche, formano legami eterofilici con una catena
oligosaccaridica di una glicoproteina presente sulla membrana della cellula adiacente. Anche le
integrine interagiscono con molecole differenti, per esempio le ICAM, mediante adesione
eterofilica.
SUPERFAMIGLIA DELLE IMMUNOGLOBULINE
Dominio immunoglobulinico (Iglike domain): fatto da una coppia
di foglietti b uniti da interazioni
idrofobiche e da un ponte
disolfuro.
LE CADERINE
Tipo di caderina
E-caderine
P-caderine
N-caderine
Distribuzione cellulare
Embrione, tessuti epiteliali
non neuronali
Trofoblasto, placenta
Tessuto nervoso, tessuto muscolare
scheletrico e cardiaco
Attivazione delle caderine per mezzo
del calcio e riconoscimento omofilico
Le caderine servono per far aderire tra loro le cellule appartenenti
allo stesso tipo di tessuto.
Modello di cell sorting. Se due tipi di cellule con caderine
differenti vengono mescolate, le cellule con lo stesso tipo di
caderine si riconoscono e aderiscono selettivamente, formando
due gruppi cellulari separati. Il sorting cellulare è importante
durante i processi di sviluppo.
Modello dell’inibizione da contatto. Se cellule cellule normali contattano
l’una con l’altra durante il loro movimento, si riconoscono
reciprocamente, cessano di muoversi e cambiano direzione. Allo stesso
modo, se il numero di cellule in una coltura aumenta e le cellule
cominciano a contattarsi, si ha un arresto della divisione cellulare. Questi
fenomeni sono promossi anche dalla presenza di caderine.
Le caderine stabilizzano anche le reti del citoscheletro e possono
influenzare l’espressione genica. La porzione intracellulare delle caderine
aggancia l’actina citoscheletrica attraverso proteine (catenine) che fanno da
intermediarie e che possono essere trasduttori del segnale.
catenine
Una volta che le cellule si sono riconosciute come appartenenti allo
stesso tessuto attraverso le caderine, si instaura fra loro un più
stretto rapporto mediante la formazione di GIUNZIONI. Le caderine
formano le GIUNZIONI ADERENTI e i DESMOSOMI. Nelle cellule si
possono poi formare GIUNZIONI GAP e GIUNZIONI TIGHT.
Intermediate
filaments
GIUNZIONI CELLULARI
Le giunzioni cellulari sono strutture che permettono
un’associazione a lungo termine tra cellule adiacenti.
1.
2.
3.
4.
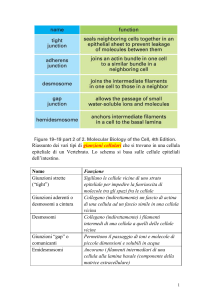
GIUNZIONI STRETTE (Tight junctions)
GIUNZIONI ADERENTI (Adherens junctions)
DESMOSOMI (Desmosomes)
GIUNZIONI GAP (Gap junctions)
SCHEMA DELLE GIUNZIONI CELLULARI NELLE
CELLULE EPITELIALI E LORO FUNZIONE
GIUNZIONI TIGHT
TIGHT JUNCTIONS
(GIUNZIONI STRETTE)
1.
Delimitano il compartimento luminale e
adluminale, impedendo la diffusione
laterale delle proteine e operandone il
loro confinamento nella porzione apicale
o in quella baso-laterale delle membrane.
2.
Sigillano la regione apicale delle cellule interconnesse in modo da impedire il
passaggio di ioni e molecole attraverso gli spazi intercellulari. Il passaggio delle
molecole e degli ioni è perciò sottoposto a selezione molecolare, in quanto
mediato da proteine transmembrana.
La tight junction è costituita da una cintura
che collega apicalmente le membrane
plasmatiche delle cellule.
Le tight junctions sono costituite da creste
continue di proteine idrofobiche giunzionali
transmembrana che si interconnettono
nello spazio intercellulare a formare uno
strato impermeabile. Nelle giunzioni tight
l’adesione tra le cellule adiacenti è rinforzata
nel lato citoplasmatico da legami con i
microfilamenti actinici presenti nella zona
immediatamente sottostante la membrana
delle due cellule, lo strato corticale.
Il legame tra le proteine transmembrana (es.
occludine e claudine) e i filamenti actinici
dello strato corticale è mediato da proteine
periferiche, tra cui le ZO1, ZO2 e ZO3
(Zonula Occludens protein 1, 2 e 3)
GIUNZIONI ADERENTI
(Es. a e b- CATENINE)
La zonula adherens è un sistema di
giunzioni
cellula-cellula
calciodipendente (GIUNZIONI ADERENTI).
Sono localizzate subito sotto le tight
junctions.
DESMOSOMI E EMIDESMOSOMI
DESMOSOMA
EMIDESMOSOMA
I desmosomi sono giunzioni cellula – cellula, calcio
dipendenti, localizzati a macchia di leopardo sulla
membrana cellulare. I desmosomi conferiscono alta
resistenza alla trazione. Sono particolarmente
abbondanti nei tessuti sottoposti a stress meccanici.
DESMOSOMI
Microscopio ottico ed elettronico:
filamenti di cheratina (rosso) e
desmosomi (giallo-verde)
Le
proteine
transmembrana
appartenenti alla famiglia delle
caderine
(desmogleina
e
desmocollina)
nel
versante
citoplasmatico legano i filamenti
intermedi tramite le proteine della
placca di adesione (tra cui la
desmoplachina e la placoglobina).
GIUNZIONI CELLULA-MATRICE:
EMIDESMOSOMI E ADESIONI FOCALI
1. Entrambi i tipi di giunzione coinvolgono le integrine quali proteine
transmembrana.
2. Le integrine sono connesse alla matrice extracellulare tramite
fibronectina e laminina.
3. Differiscono per gli elementi citoscheletrici citoplasmatici a cui le
integrine sono collegate: i microfilamenti nei contatti focali; i
filamenti intermedi negli emidesmosomi.
EMIDESMOSOMI
Al microscopio elettronico gli emidesmosomi appaiono come metà desmosomi. La
placca di adesione è presente solo nel versante citoplasmatico. Sulla placca
convergono i filamenti intermedi (cheratina).
ADESIONI FOCALI
I contatti o adesioni focali sono giunzioni non ancoranti calcio dipendenti che
legano la membrana plasmatica delle cellule alla matrice extracellulare. Sono
importanti nel movimento cellulare, nella proliferazione e nel
differenziamento.
(a) Cellula statica, attaccata alla matrice extracellulare attraverso delle adesioni focali.
(b) Per iniziare a muoversi la cellula rompe alcuni siti di adesione e forma protrusioni
nella direzione del movimento.
(c) La cellula aderisce al substrato con la protrusione, formando una nuova adesione
focale.
(d) La parte posteriore della cellula si contrae per spingere il citoplasma nella direzione
del movimento.
INTEGRINE
Questo tipo di giunzione mette in comunicazione diretta le cellule. Sono
infatti costituite da gruppi di canali, i connessoni presenti nelle due
cellule adiacenti e in asse fra loro. Ogni connessone è fatto da 6 copie
della proteina connessina Il connessone si giustappone con il
connessone presente sulla membrana della cellula adiacente.
GIUNZIONE GAP
Il canale è regolato (l’apertura è a controllo
di voltaggio; si chiude in presenza di alte
concentrazione di calcio o a pH acido).
SELECTINE mediano l’interazione con
oligosaccaridi nel processo infiammatorio.
C-Type Lectin Domain
P-selectin
EGF-Like Domain
Complement Regulatory Repeat
NH2
Transmembrane Domain
Cytoplasmic Domain
E-selectin
NH2
L-selectin
NH2
Leukocyte
Endothelial Cell
COOH
COOH
COOH
L-selectina: neutrofili, monociti, linfociti B e T.
P-selectina: granuli a delle piastrine non attivate e
corpi di Weibel-Palade delle cellule endoteliali
(espressa costitutivamente nelle piastrine ed
indotta nelle cellule endoteliali).
E-selectina: cellule endoteliali e siero (espressione
indotta da citochine).
Rolling leucocitario nella
risposta infiammatoria
http://bcs.whfreeman.com/lodish5e/pages/bcsmain.asp?v=category&s=00010&n=06000&i=06010.01&o=|00510|00520|00
530|00540|00560|00570|00590|00600|00700|00010|00020|00030|00040|00
050|01000|02000|03000|04000|05000|06000|07000|08000|09000|10000|11
000|1203&n
Lo spazio fra le cellule
Matrice extracellulare
Proteoglicani,
glicoproteine,
acido ialuronico
Fibre collagene
ed elastiche
Proteine multiadesive
COLLAGENE
Il collagene è una proteina fibrosa
fatta di tre catene polipeptidiche
che formano una struttura a tripla
elica simile ad una corda che
fornisce resistenza alla trazione
alla matrice extracellulare (ECM)
Le
catene
di
collagene
contengono sequenze ripetute GX-Y (glicina e frequentemente
prolina e idrossiprolina, ma anche
lisina e idrossilisina).
La glicina è piccola ed è l’unico
amminoacido che entra nella
regione interna della tripla
elica. L’idrossiprolina, grazie al
gruppo –OH, forma legami
idrogeno che stabilizzano la
tripla elica.
L’idrossiprolina e l’idrossilisina, entrambe
frequenti nel collagene, sono formate a
partire da pro e lys dopo la traduzione,
grazie all’azione di idrossilasi (la vitamina C
è un loro cofattore).
RAPPRESENTAZIONE SCHEMATICA DI UNA
MOLECOLA DI TROPOCOLLAGENE DI TIPO I
Due catene polipeptidiche α1 e una catena α2).
Ogni molecola è lunga 280 nm e un giro
completo dell'elica è pari a 8,6 nm.
Molecola di procollagene
Molecola di tropocollagene (280 nm)
Fibrilla di collagene (formazione di
legami covalenti).
Le fibre di collagene (collagene di tipo 1, 2, 3) si formano per interazioni laterali delle
molecole di collagene stabilizzate da legami covalenti fra il C-terminus di una molecola
e l’N-terminus di quella adiacente. Gli amminoacidi coinvolti nel legame sono lisina e
idrossilisina. L’elicatura è possibile perché esistono delle forti ripetizioni di Gly-Pro-X.
MICROFIBRILLA DI COLLAGENE
(schema e microfotografia ottenuta
dopo colorazione negativa). Le
zone
chiare
(zona
di
sovrapposizione) appaiono tali in
quanto il colorante non penetra in
esse; quelle scure (zona vuota) si
presentano tali in quanto il
colorante si deposita negli intervalli
tra l'allineamento testa-coda di una
molecola di tropocollagene con
un'altra. In due filamenti affiancati
le molecole di tropocollagene sono
sfasate di circa un quarto della loro
lunghezza (64-70 nm). Da questa
organizzazione deriverebbe la
struttura
periodica
delle
microfibrille elementari. Il periodo
è dato dalla somma di un
segmento chiaro e uno scuro.
×175.000.
Ogni molecola di precursore del collagene viene prodotto dalle cellule del connettivo
come una proteina da esportazione glicosilata. Tre molecole poi si elicano e formano il
tropocollagene che ha, alle estremità, dei segmenti peptidici non elicati che verranno
rimossi. L’allineamento laterale avviene nello spazio extracellulare.
Esistono diversi tipi di collagene. L’allineamento
delle molecole di tropocollagene a formare fibrille
avviene, ad esempio, per il collagene di tipo I
presente nei connettivi o nei tendini, mentre nella
regione adiacente alle cellule degli epiteli il
collagene che è di tipo IV, assume una forma a rete
e va a costituire la lamina basale.
L’altra proteina fibrosa della matrice extracellulare è l’elastina. Le
molecole di elastina sono legate insieme da legami covalenti che
costituiscono una rete. In questo modello, ciascuna molecola di
elastina può espandersi e contrarsi e indurre un allungamento o
una contrazione nell’intero complesso assemblato.
ELASTINA
GLICOPROTEINE E PROTEOGLICANI
Nella matrice sono presenti anche i PROTEOGLICANI,
peptidi a cui sono covalentemente legati dei polimeri disaccaridici detti
glicosamminoglicani (GAG).
I proteoglicani possono essere legati alle cellule o possono essere liberi nella
matrice.
GLICOSAMMINOGLICANI
Sono prodotti nell’apparato di Golgi.
Molte unità disaccaridiche sono solfatate.
Aggancio del GAG al peptide.
IALURONANO (ACIDO IALURONICO).
E’ un GAG non solfatato dato dalla ripetizione
di acido glucuronico e N-acetil-glucosammina.
Forma gel viscosi e idratati che resistono alle
compressioni.
AGGRECANO
Uno dei più importanti proteoglicani è l’aggrecano, il proteoglicano più
presente nella cartilagine.
Proteine adattatrici
Molecola di aggrecano
Più molecole di aggrecano possono poi
legarsi allo ialuronano o acido
ialuronico (100:1) attraverso proteine
adattatrici a formare aggregati di
proteoglicani-ialuronano che possono
essere grandi come batteri (5µm di
lunghezza).
STRUTTURA SCHEMATICA DI UNA MOLECOLA DI AGGRECANO
ASSOCIATA ALL’ACIDO IALURONICO
PROTEINE MULTIADESIVE
FIBRONECTINA
Un componente essenziale della matrice extracellulare (ECM),
facente parte delle proteine multiadesive, è la proteina
fibronectina, che si assembla in fibrille che permettono alle
cellule di agganciarsi alla matrice.
Le integrine legano ed esercitano forze sulla fibronectina e accoppiano
meccanicamente il citoscheletro di actina alla ECM attraverso un
elaborato complesso di adesione (adesioni focali). Il legame tra integrine
e fibronectina è fondamentale nelle cellule migranti.
LAMININA
Questa proteina multiadesiva fa da ponte fra le integrine delle cellule
epiteliali, non migranti, e la matrice. La si ritrova nella lamina basale.
Gli epiteli poggiano su una lamina basale sotto la
quale troviamo il connettivo.
La lamina basale è composta da collagene di tipo
IV, da proteoglicani e da laminina, una
proteina multiadesiva che interagisce anche con
le integrine
Collagene IV
Proteoglicani
Laminina
Integrina
Tutte le integrine sono proteine di adesione transmembrana
eterodimeriche. Mostrano un effetto velcro: sono forti messe
insieme, ma deboli prese singolarmente. Sono attive in presenza di
Ca2+ o Mg2+ e collegano la matrice extracellulare al citoscheletro.
Ci sono 24 tipi di catene α e 9 di catene β che si combinano a
formare integrine diverse (famiglie di integrine) aventi specificità
cellulari e ruoli diversi.
α5β1 binds fibronectin.
α6β1 binds laminin.
α7β1 binds laminin in muscle.
Le integrine legano le cellule alla lamina basale da sole o
attraverso l’interazione con l’actina (adesioni focali) o i filamenti
intermedi (emidesmosomi). Anche i proteoglicani possono
legare la cellula alla lamina basale.
INTEGRINE E TRASDUZIONE DEL SEGNALE
Il legame delle integrine alla membrana basale porta anche alla formazione delle
adesioni focali (focal adhesions, FA). FAK (Focal Adhesion Kinase) è una chinasi
coinvolta nel controllo del comportamento della cellula mediato da integrine.
Quando FAK viene autofosforilata in seguito ad adesione, FAK diviene un
regolatore di migrazione cellulare.
ALCUNE FUNZIONI DELLA LAMINA BASALE
Determina la polarità della cellula
Organizza e lega le cellule
Permette la migrazione cellulare
Una forma specializzata di lamina basale si trova nel
glomerulo renale, dove assolve il compito di filtrare il
sangue che arriva al rene.
IL DIFFERENZIAMENTO CELLULARE
TOTIPOTENZA
Massa cellulare
interna
PLURIPOTENZA
Da blastocisti a gastrula
Meiosi: un tipo speciale di divisione cellulare che richiede proteine speciali
Perché la fecondazione?
La vita di una cellula all'interno dell'organismo è sempre
in bilico. Privata dei contatti con il proprio microambiente (tessuto) o delle vie di comunicazione
(connessioni nervose) la cellula va in contro a morte
cellulare programmata. E' solo la continua verifica di
"appartenenza" all'ambiente che la circonda che
impedisce alla cellula di suicidarsi e le consente, per il
momento la vita.
Un breve riferimento
all’APOPTOSI…
MEMBRANA
PLASMATICA
MEMBRANA
MITOCONDRIALE
ESTERNA
http://bcs.whfreeman.com/lodish5e/pages/bcsmain.asp?v=category&s=00010&n=22000&i=22010.01&o=|00510|00520|00530|00540|0
0560|00570|00590|00600|00700|00010|00020|00030|00040|00050|01000|02000|03000|0
4000|05000|06000|07000|08000|09000|10000|11000|120
APOPTOSI: morte cellulare programmata
Frammentazione della cromatina
Frammentazione del nucleo
Esposizione della fosfatidilserina nel foglietto esterno
del doppio strato lipidico
http://sciencestage.com/v/308/biology-cell-embryo-ovaryegg-fallopian-fetus-fetal-blastocyst-uterus-placentapregnant-pregnancy-fertilize.html