Il potenziale di membrana
La differenza di concentrazione di tutti gli ioni tra l'interno e l'esterno della cellula crea una differenza di
potenziale elettrico tra l'interno e l'esterno. Questa tensione elettrica è detta potenziale di membrana e
nei diversi tipi di cellule umane va dai -15mV ai -100mV. In particolare il segno meno indica che l'interno
ha potenziale negativo rispetto all'esterno. Nelle cellule nervose il potenziale è normalmente di -70mV
ma poichè esso può variare, il livello di -70mv viene detto potenziale di riposo.
-70 mV
Il potenziale d’azione
Se si stimola una cellula nervosa questa apre alcuni suoi canali ionici e un certo numero di ioni passano
attraverso la membrana riducendone il potenziale o addirittura invertendolo. Questa variazione di
tensione elettrica (o voltaggio) tra interno ed esterno della cellula viene detta depolarizzazione e cessa
spontaneamente entro tempi di millisecondi o decine di millisecondi riportando il potenziale di
membrana a livello del potenziale di riposo.
Alcuni anestetici locali agiscono
bloccando i canali ionici della
membrana delle cellule nervose
impedendo che la cellula si depolarizzi
ovvero impedendo che essa trasmetta
verso il cervello l'informazione del
dolore generato in periferia.
L’anestetico locale entra nella cellula
dove blocca temporaneamente il
canale voltaggio dipendente per il Na+
Vengono generalmente somministrati
assieme a vasocostrittori (adrenalina),
per prolungare la presenza
dell'anestetico sul sito d'azione evitando
la sua diffusione a livello sistemico.
Anche per ridurre il sanguinamento nel
caso di piccoli interventi cutanei e avere
un campo operatorio più pulito.
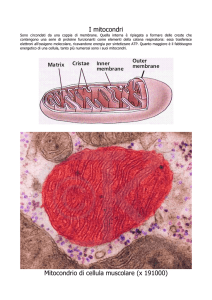
I mitocondri sono organuli citoplasmatici il cui interno è costituito da molte pieghe membranose a
doppio strato dette creste, che costituiscono una superficie estesa a cui sono ancorati i componenti per
la respirazione aerobica e per la produzione di ATP.
Il mitocondrio contiene enzimi, ribosomi e DNA.
Il mitocondrio è la struttura che utilizza ossigeno per estrarre energia.
Gli organismi anaerobi non possono utilizzare l'ossigeno e questo limita molto la quantità di energia che
essi possono ricavare dal nutrimento. In altre parole essi scartano sostanze ancora ricche di energia e
quindi non completamente sfruttate. Gli organismi aerobi (quali l'uomo e la maggior parte degli organismi
procarioti ed eucarioti) utilizzano l'ossigeno per metabolizzare completamente prodotti (es. l'acido lattico)
fino a ridurli a CO2 e H2O.
Vediamo brevemente come avviene il primo passo della produzione di energia in una cellula eucariotica. Il
glucosio entrato nel citoplasma viene metabolizzato dagli enzimi della glicolisi i quali producono piruvato,
NADH e ATP.
Si tratta di processi di deidrogenazione in cui viene rimosso idrogeno dal substrato (il substrato viene
ossidato) e l'accettore di idrogeno (ossidante) non è l'ossigeno, bensi molecole complesse (es. NAD e FAD).
diidrossiacetone
fosfato (DHAP)
gliceraldeide-3-fosfato
il diidrossiacetone fosfato viene convertito in un'altra molecola di gliceraldeide 3-fosfato
Fin qui vengono
prodotte solo due
molecole di ATP per
ogni molecola di
glucosio e viene
scartato il piruvato che
è ancora molto ricco di
energia.
A questo punto se non
vi è ossigeno il
piruvato è ridotto a
lattato ed esce dalla
cellula.
Dopo la glicolisi, negli organismi aerobi segue la respirazione. Questa è suddivisa in ciclo dell'acido citrico
o di Krebs, che si realizza nella matrice del mitocondrio, e nella catena di trasporto degli elettroni, che è
localizzata sulle creste mitocondriali.
Da notare che nel ciclo di Krebs entrano anche polisaccaridi, grassi e metaboliti delle proteine e da tutti
questi composti viene estratta energia.
La contrazione muscolare richiede energia e questa viene fornita dall'ATP. A seguito di uno sforzo intenso,
le riserve di ATP nel muscolo si esauriscono velocemente. A questo punto viene utilizzato il creatinfosfato
presente nelle cellule muscolari.
Dopo meno di un minuto anche le riserve di creatinfosfato si esauriscono e altra energia viene ottenuta
dal metabolismo ossidativo.
Le fibre muscolari veloci hanno
pochissimi mitocondri per cui
possono prodursi pochissimo ATP
per respirazione aerobica.
Essi producono ATP
anaerobicamente dalla glicolisi,
che pur producendo poco ATP
ha il vantaggio di essere un
processo veloce. Il problema è
che questo processo consuma
velocemente il glucosio e produce
acido lattico. L'acido lattico abbassa
il pH muscolare, provoca dolori e
crampi. L'acido lattico viene trasportato
dal sangue al fegato che lo trasforma in glucosio.
Le fibre lente contengono cellule con molti mitocondri e lavorano negli esercizi aerobici. Un esercizio
aerobico (andare in bicicletta o camminare a un buon ritmo) può durare molto più a lungo senza avvertire
la fatica poichè si produce ATP in aerobiosi e quindi senza la produzione di acido lattico.
Dopo aver esaurito il glucosio vengono
utilizzati gli acidi grassi. Questi contengono
molti atomi di H per cui la loro
beta-ossidazione libera molta più energia
rispetto a un peso equivalente di molecole
glucidiche.
Il cuore produce ATP soltanto attraverso
il metabolismo aerobico.
E’ possibile allenarsi per aumentare la soglia aerobica
Interazioni tra cellule
Le cellule di uno stesso organo devono interagire tra loro per realizzare un’azione coordinata, come ad
es. crescita, migrazione, differenziamento, mantenere la forma macroscopica di un organo.
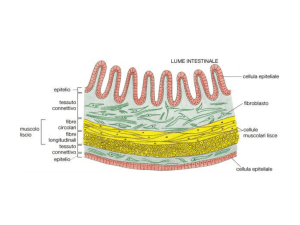
Lo strato esterno
della pelle,
costituito da tessuto
epiteliale, è
composto di cellule
legate tra loro
tramite una sorta di
piccole giunzioni o
piccoli ponti che
permettono sia di
mantenere le cellule
tra loro adese sia di
permetterne la
comunicazione
Lo strato più profondo della pelle ha un
tessuto di tipo connettivo, che è costituito
principalmente da materiale extracellulare e
le cellule si trovano intersperse ma non a
contatto le une con le altre. Queste cellule
comunicano tra loro attraverso recettori.
Una proteina della matrice extracellulare è il
collagene, una glicoproteina che
ha elevata resistenza alla trazione.
Esso tiene insieme il tessuto
connettivo.
Il collagene è prodotto principalmente dai fibroblasti
e secondariamente dalle cellule muscolari lisce ed
epiteliali. Il collagene è formato da più polipeptidi
legati insieme e la vitamina C è importante per
stabilire la formazione di questi legami.
I sintomi dello scorbuto
(mancanza di vitamina C)
quali infiammazione delle
gengive, perdita dei denti,
ossa fragili, difficile
guarigione delle ferite,
indebolimento della parete
dei vasi sanguigni... sono
dovuti proprio alla
mancanza di legami crociati
per formare il collagene.
Anche la
diminuita
elasticità della
pelle e
l'aumentata
fragilità delle
ossa negli
anziani è dovuta
a problemi nella
formazione del
collagene.
I tendini che connettono
i muscoli con le ossa
devono
resistere
a
notevoli forze di trazione
e questo è possibile
grazie all'organizzazione
tridimensionale
particolare
delle
molecole di collagene.
I proteoglicani sono un complesso di proteine e polisaccaridi che attirano a sè molecole di acqua per
cui si forma un gel poroso e umido che può riempire gli spazi extracellulari e resistere alle forze di
compressione. I proteoglicani e i collageni insieme formano la cartilagine, se sono impregnati di fosfato
di calcio compongono la matrice extracellulare dell'osso.
La fibronectina è una proteina della matrice extracellulare coinvolta nella migrazione cellulare, in
particolare durante lo sviluppo embrionale. La fibrioncetina costituisce una sorta di letto lungo il quale
le cellule migrano, in altre parole segnerebbe il percorso di migrazione delle cellule. La fibronectina è
fondamentale per la formazione delle strutture di un organismo, per la formazione delle fessure e
della ramificazione di organi quali ghiandole salivari, rene e polmone.
La laminina è anch'essa una
glicoproteina coinvolta nella
migrazione cellulare, quindi
nella crescita e nel
differenziamento cellulari.
Le integrine sono proteine integrali di membrana capaci di legarsi selettivamente a certe sostanze
(ligandi) dell'ambiente extracellulare. Esistono diversi tipi di integrine per legare ligandi diversi. L'integrina
è formata da una subunità alfa e una subunità beta. Le due catene sporgenti all'esterno della cellula
possono assumere due conformazioni:
1) una ripiegata e inattiva, cioè incapace di legare sostanze
2) conformazione distesa e attiva
Ciò che determina il cambiamento di configurazione dipende dalla parte intracellulare delle due
estremità alfa e beta. In particolare, se nell'estremità interna beta si lega una proteina (talina), l'integrina
viene attivata.
L'integrina attiva
potrebbe legarsi
ad es. al collagene.
L'aggregazione delle piastrine per formare un coagulo avviene in conseguenza all'attivazione delle
integrine che, così attivate, legano il fibrinogeno.
Notare che l'aggregazione piastrinica in una zona o in un momento sbagliati possono portare a un
pericoloso coagulo di sangue (trombo). La sua pericolosità consiste nel fatto che esso può bloccare
l'afflusso di sangue in organi vitali (infarto e ictus).
Che cosa media l'interazione tra cellule e fa si che una cellula aderisca soltanto a un'altra dello stesso
tipo ?
1) selectine, riconoscono e legano zuccheri nella superficie di altre cellule, riguardano le cellule
endoteliali, le piastrine e i leucociti. Mediano le interazioni fra i leucociti circolanti e le pareti dei vasi, nei
luoghi di infiammazione e coagulazione. Mediano l'adesione degli embrioni alla parete dell'utero.
2) certe immunoglobuline. Solitamente mediano le interazioni specifiche tra linfociti e macrofagi, o altri
linfociti o cellule bersaglio. In questo caso favoriscono l'adesione tra cellule del sistema nervoso (anche
crescita degli assoni). Mutazioni
nei geni di queste proteine
provocano ritardo mentale,
difficoltà nel controllo dei
movimenti.
Queste immunoglobuline
legano ligandi proteici,
ad es. le integrine.
3) alcune integrine.
Le immunoglobuline
appena viste legano ligandi
proteici, ad es. le integrine.
4) caderine. Legano altre
caderine dello stesso tipo
perciò mediano l'adesione
tra cellule dello stesso tipo.
Durante l'infiammazione i
tessuti danneggiati emettono
segnali chimici che modificano
l'adesività delle cellule
endoteliali delle venule.
Le cellule endoteliali rispondono
esprimendo selectine.
Le pareti delle venule diventano
adesive per cellule circolanti
quali neutrofili e leucociti.
Queste cellule si appiattiscono e
passano, attraverso le cellule
endoteliali, nel tessuto
danneggiato.
Nella sindrome da deficienza di
adesione leucocitaria, manca
una subunità dell'integrina e i
leucociti non riescono ad
aderire allo strato endoteliale
delle venule. Questi pazienti
soffrono di infezioni e tali
infezioni tendono ad aggravarsi
anzichè risolversi.
Nel cancro le cellule diventano in grado di proliferare in modo non regolato. Finchè tali cellule
rimangono confinate in una massa compatta si può esportarle chirurgicamente. Purtroppo in molti
tumori maligni le cellule riescono a staccarsi dalla massa tumorale, a spostarsi attraverso il circolo
sanguigno o linfatico e a far crescere una nuova massa tumorale. Questa diffusione si chiama metastasi.
L'adesività cellulare è fortemente implicata in questo processo di formazione di nuove masse tumorali.
Pensiamo che le cellule che si staccano da una massa principale devono avere una minore adesività per
potersi liberare, devono essere in grado di penetrare barriere quali la matrice extracellulare del tessuto
connettivo, devono invadere un tessuto normale. Un difetto o una diminuita produzione della proteina
“caderina E” provoca la formazione di metastasi.
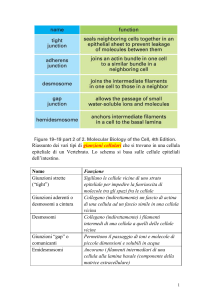
I meccanismi fondamentali coinvolti nell'ancoraggio cellula-cellula sono : le giunzioni aderenti, le giunzioni
strette e i desmosomi.
Le giunzioni occludenti sono formate da occludina e claudine. Uniscono cellule epiteliali vicine. Sono alla
base della funzione renale per permettere una permeabilità selettiva, sono importanti nell'epidermide
per trattenere l'acqua ed evitare la disidratazione, sono importanti nell'endotelio che riveste le pareti dei
capillari, nel cervello per formare la barriera emato-encefalica.
Le giunzioni aderenti sono legami tra le estremità extracellulare delle molecole di caderina, quindi
connettono l'ambiente extracellulare al citoscheletro di actina e servono a trasmettere segnali
dall'esterno all'interno della cellula.
Giunzione aderente
I desmosomi sono giunzioni contenenti caderine e sono tipici dei tessuti sottoposti a stress meccanico.
Uniscono le cellule in alcuni punti precisi come una sorta di bottoni ma permettono il movimento di
materiale tra le cellule. Ha un ruolo meccanico.
Le giunzioni comunicanti o serrate (gap junctions) sono siti di comunicazione tra i citoplasmi di cellule
adiacenti, sono formati da connessina e permettono il passaggio di sostanze da una cellula all'altra,
sembrerebbero poco selettivi. Queste giunzioni possono mettere in comunicazione le cellule di un
organo. In questo modo, se un ormone ha stimolato alcune cellule, questo stimolo può essere
trasmesso alle cellule vicine propagando l'AMP ciclico o inisitoli fosfati attraverso le giunzioni
comunicanti.
La propagazione della contrazione delle cellule cardiache avviene perchè un flusso ionico può propagarsi
da una cellula all'altra tramite le giunzioni comunicanti. Similmente succede per la propagazione delle
onde peristaltiche nell'intestino.