Apparato gastrointestinale
June 23, 2011
Contents
1 Motilità
2 Secrezione
2.1
2.2
2.3
2.4
2.5
La saliva . . . . . . . .
La secrezione gastrica
Il succo pancreatico .
La bile . . . . . . . . .
Il succo intestinale . .
3
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
3 Digestione ed assorbimento
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
5
. 6
. 7
. 8
. 9
. 11
11
3.1 I carboidrati . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 12
3.2 Le proteine . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 12
3.3 I lipidi . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 12
1
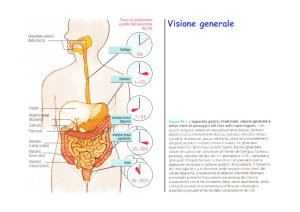
L'apparato gastrointestinale è deputato a far si che nutrienti, acqua ed elettroliti possano passare dall'ambiente
esterno, cioè il lume del sistema, a quello interno; questo compito è assolto per tramite di quattro diversi processi:
•
Digestione, cioè l'insieme delle procedure siche e chimiche che riducono gli alimenti in molecole assorbibili
•
Motilità, cioè l'insieme di movimenti che garantiscono la progressione del cibo lungo il sistema
•
Secrezione, cioè la fornitura di sostanze in grado di operare la digestione chimica
•
Assorbimento, cioè il passaggio sico di sostanze tra i due ambienti
I quattro processi fondamentali sono regolati da meccanismi sosticati di tipo nervoso ed endocrino.
La struttura dell'apparato GI è specializzata nei vari punti a seconda della funzione locale, ma è possibile
riconoscere uno schema di base abbastanza costante:
Lo strato più interno è rappresentato dalla mucosa, dove è evidente una supercie irregolare con villi che ha lo
scopo di massimizzare la supercie di scambio; l'epitelio che costituisce questa supercie ha funzioni di trasporto
in entrambe le direzioni, quindi sia assorbimento che secrezione. La mucosa è circondata in direzione esterna
dalla sottomucosa la quale contiene il sistema linfatico e i grandi vasi oltre alle ghiandole endocrine ed esocrine
locali. La muscolatura liscia del tubo è organizzata secondo due piani funzionali; esistono una muscolatura liscia
interna circolare ed una esterna longitudinale. La sierosa è l'ultima tonaca evidente e si fonde al peritoneo
per delimitare la parete esterna del canale. Questo schema anatomico è vericato solo se si escludono le estremità
del sistema: la bocca (no al primo terzo dell'esofago) e l'ano hanno una muscolatura di tipo striato e quindi anche
il controllo nervoso sarà diverso.
La regolazione di questo sistema è organizzata in maniera gerarchica con al vertice i centri encefalici superiori
che agiscono in periferia per tramite delle divisioni autonome del SNC. La grande particolarità è che il SNA non
agisce direttamente, ma va a regolare l'attività del sistema nervoso enterico, una divisione separata che controlla
la motilità e la secrezione del tratto GI in modo pressochè autonomo.
2
Il parasimpatico agisce sul SNE per tramite del nervo vago (no alla essura splenica) o per tramite dei nervi
pelvici, mentre il simpatico agisce grazie alle bre in uscita dalle colonne intermediolaterali dei segmenti spinali
da T1 a L3 circa. Il SNE enterico è organizzato in vari plessi tra i quali i principali sono il plesso sottomucoso e il
plesso mienterico, che sono gli unici a contenere anche i corpi cellulari dei neuroni e non le sole bre di passaggio.
In termini quantitativi i neuroni del SNE sono tanti quanti quelli del midollo spinale ma sono distribuiti in una
regione molto più ampia e quindi si ha un aspetto simile ad una rete; questo sistema è in grado di funzionare in quasi
totale autonomia e di fatto il ruolo dei centri encefalici superiori può essere paragonato a quello del cervelletto in
ambito motorio: sicuramente ranano i vari processi, ma non ne sono la causa scatenante e non sono fondamentali.
Nella parete del tratto GI sono presenti meccanocettori e chemocettori che monitorano costantemente le condizioni;
le informazioni raccolte vengono inviate per tramite di aerenze locali al SNE che le rielabora e mobilita gli eettori,
che possono essere cellule muscolari lisce, cellule endocrine, esocrine o i vasi ematici. Questo tipo di schema è quello
di un riesso e infatti si parla di riessi corti o riessi intrinseci di regolazione. Le informazioni dai recettori
vengono in realtà inviate anche al SNC tramite aerenze splancniche o vagali dove vengono valutate e in base ad esse
vengono inviati ordini soprattutto ai plessi mienterico e sottomucoso ma anche a vie dirette: si parla in questo caso
di riessi lunghi o riessi estrinseci di regolazione. I recettori sono sensibili sia a stimolazioni meccaniche che
a stimolazioni chimiche, ad esempio alla serotonina. I risultati delle elaborazioni possono portare a diverse risposte;
esistono motoneuroni eccitatori che liberano acetilcolina e sostanza P e fanno contrarre le cellule muscolari
circostanti, ma esistono anche motoneuroni inibitori che inibiscono le contrazioni per tramite di ossido nitrico
e peptide intestinale vasoattivo. In generale, a seconda del tipo di risposta attuata, si riconoscono meccanismi
endocrini (mediati da ormoni), meccanismi neurocrini (mediati dall'attività neuronale diretta) e meccanismi
paracrini (mediati da sostanze con azioni a medio raggio). Una particolarità del controllo neurocrino è la sua
capacità unica di poter essere anticipatorio e non esclusivamente a feedback: il pensiero del cibo ad esempio può
essere suciente ad iniziare a mobilitare il tratto GI.
1
Motilità
La maggior parte della muscolatura GI è di tipo liscio. La registrazione dei potenziali di queste cellule muscolari
lisce evidenzia un potenziale di riposo interrotto da onde lente di depolarizzazione la cui frequenza dipende dalla
sede anatomica analizzata. Queste onde lente sono molto più durature dei potenziali d'azione e non ne condividono
neanche la natura: si tratta infatti di semplici depolarizzazioni che vengono generate da cellule pacemaker e diuse
per accoppiamento di giunzioni elettriche. La contrazione insorge quando la depolarizzazione raggiunge e supera la
soglia di contrazione; esiste anche una seconda soglia, detta soglia elettrica, superata la quale si ha l'insorgenza
di un potenziale d'azione e la contrazione diventa più vigorosa. I movimenti che si possono ritrovare nell'apparato
GI sono essenzialmente quattro:
•
Propulsione, cioè i movimenti che fanno progredire i contenuti intestinali
•
Triturazione, cioè i movimenti che riducono i contenuti in pezzi più piccoli
3
•
Rimescolamento, cioè i movimenti che incorporano le varie secrezioni ai contenuti
Contrazioni toniche, cioè i movimenti che realizzano vari compartimenti a partire da un tubo omogeneo
Il più classico movimento propulsivo è la peristalsi, nel quale si distinguono un segmento recettivo ed un
segmento propulsivo. Il segmento recettivo vede la sua muscolatura longitudinale contrarsi e quella circolare
•
rilassarsi: il risultato è un aumento del diametro per poter accogliere il cibo. Il segmento propulsivo inverte lo
schema contrattile e di fatto spreme i suoi contenuti in avanti. Questo tipo di schema motorio può essere invertito
in caso di necessità, generando il vomito. Un discorso separato è da farsi per gli snteri, i quali presentano una
contrazione tonica in assenza di stimoli in quanto devono separare due compartimenti; molto importante è lo
sntere esofageo inferiore, il cui ruolo è prevenire l'arrivo di succhi gastrici acidi nell'esofago. La patologia del
reusso esofageo (GERD) nasce proprio da un mancato funzionamneto di questo sntere. Un caso particolare è
quello dell'ano, che presenta due snteri con controlli diversi; lo sntere anale interno è costituito da muscolatura
liscia ed è sotto controllo essenzialmente riesso, lo sntere anale esterno è striato e sotto controllo volontario.
La motilità inizia a livello della bocca dove si ritrova come primo movimento quello della masticazione, i cui
ruoli sono di triturazione e rimescolamento del cibo con la secrezione salivare. La masticazione è un movimento
ritmico e viene divisa in tre fasi: apertura e chiusura della mandibola e fase occlusale. Il generatore centrale di
schemi motori per questo movimento ritmico si trova a livello bulbo-pontino e, come per gli altri generatori, non
ha bisogno di aerenze per funzionare, ma in presenza di queste l'adattamento del movimento è molto migliore. Il
centro della masticazione deve coordinare questa attività motoria con quella di guancia e lingua, ma anche con il
respiro ed il linguaggio, in modo da non interferire con gli atti in corso.
La masticazione frantuma il cibo e lo mescola con la saliva, rendendolo così pronto per la deglutizione, cioè
il passaggio in direzione dello stomaco. La delgutizione è un movimento propulsivo di origine volontaria ma che
prosegue in modo involontario; si tratta di un'attività motoria ritmica con un generatore centrale situato a livello
del bulbo. Il movimento viene diviso in una fase orofaringea e in una fase esofagea. La fase orofaringea inizia
quando il bolo di cibo viene spinto nel terzo posteriore della lingua, sede dalla quale ha solo due vie possibili: il
rigetto in avanti, come nei bambini che non vogliono mangiare, o la discesa in esofago; la salita in direzione delle vie
aeree è impedita dalle modiche cui vanno incontro la faringe e il palato molle. La fase orofaringea ha una durata di
circa un secondo o poco più, al termine del quale il cibo si trova ad essere all'imbocco dell'esofago. Il generatore dei
movimenti di deglutizione, come quello della masticazione, coordina questa attività a quella della respirazione e della
fonazione (entrambe risultano inibite). L'intera deglutizione può essere analizzata secondo gradienti pressori, che
in ultima analisi sono ciò che guida il cibo nel suo percorso.
La pressione al di sopra dello sntere esofageo superiore è uguale a quella atmosferica per via della comunicazione
diretta. A livello dello sntere esofageo superiore la pressione è molto più alta per via della contrazione tonica della
muscolatura. La porzione esofagea all'interno del mediastino vede al suo interno una pressione subatmosferica grazie
agli eetti della pressione mediastinica generata dall'apparato respiratorio mentre a livello sottodiaframmatico si
ha una pressione superatmosferica per via della muscolatura addominale. Quando inizia la fase esofagea lo sntere
superiore si rilascia e il cibo inizia a seguire il gradiente pressorio e attraversa dunque l'esofago: si parla di peristalsi
primaria. L'esofago deve risultare svuotato a seguito della deglutizione, quindi se la peristalsi primaria non riesce
a farlo interviene una peristalsi secondaria di natura riessa; l'attivazione dei meccanocettori esofagei produce
4
questo secondo tipo di peristalsi (che non ha dunque un centro generatore come la primaria) che assicura l'assenza
di residui nell'esofago. La fase esofagea dura circa otto secondi per un cibo di natura solida, ma si riduce a un solo
secondo per i liquidi; un atto di deglutizione oscilla in durata dunque tra i due e i nove secondi.
Lo stomaco è suddiviso in termini anatomici in fondo, corpo, antro e zona pilorica; secondo un punto di vista
funzionale è invece diviso in due sole regioni: una regione serbatoio che presenta contrazioni toniche e una
pompa antrale che presenta condizioni fasiche. La regione serbatoio ha il compito di dilatarsi quanto più
possibile per accogliere il cibo in arrivo senza orire vistosi aumenti di pressione; una prima dilatazione prende il
nome di rilasciamento recettivo: si tratta di un meccanismo riesso attivato da recettori a livello di bocca e
faringe. Grazie al rilasciamento recettivo lo stomaco si fa trovare già dilatato quando il cibo vi fa ingresso. Un
secondo meccanismo contiene gli aumenti pressori: si tratta del rilasciamento adattativo; questo rilasciamento
viene generato dall'attivazione di recettori da stiramento interni allo stomaco stesso, con il risultato che l'organo
più si riempie e più si rilascia. Terzo e ultimo contributo al rilasciamento gastrico arriva dal duodeno, il quale si
presenta dotato di chemocettori che monitorano i contenuti in arrivo e regola per via paracrina l'attività gastrica.
Il contenuto gastrico è ricco in HCl e questa acidità è monitorata dal duodeno: un meccanismo a feedback centrato
sulla molecola di secretina, inibisce lo svuotamento e riduce il pH duodenale. La regione della pompa antrale ha
un ruolo sia nella progressione del cibo che nella progressione. Le contrazioni fasiche di questa regione spingono
i contenuti verso lo sntere pilorico, tuttavia appena una quantità di essi lo supera va ad attivare i chemocettori
duodenali che promuovono la contrazione del piloro stesso; le contrazioni della pompa intanto continuano ma questa
volta lo sntere è chiuso quindi il cibo torna indietro: questo rimescola e tritura il cibo in modo molto ecace.
I primi novanta minuti a seguito dell'assunzione di cibo vedono una totale assenza di movimenti nel tratto GI:
si parla di ileo siologico o fase I. L'ileo siologico si contrappone all'ileo paralitico, che è invece il risultato di
manovre di tipo chirurgico a livello addominale. La fase I viene interrotta dalle successive fasi II e III in cui si ha
prima un'attività motoria irregolare (II) e poi più intensa e regolare (III): queste attività originano nello stomaco e
si dirigono poi in senso anale no alla valvola ileocecale. L'attività motoria prende il nome di complesso motorio
migrante ed è generata da cellule pacemaker (cellule del Kajal?) le quali trasmettono l'impulso per tramite di gap
junctions e accoppiamenti elettrici. La caratteristica fondamentale del complesso motorio migrante è che questo
avviene solo a patto che non venga interrotto da segnali inibitori, cioè è presente solo lontano dai pasti. In rapporto
ai pasti i movimenti tipici sono quelli di segmentazione e di peristalsi; i movimenti di segmentazione tendono a
rimescolare il contenuto dell'intestino ma non hanno la funzione di indirizzarlo verso una particolare regione. I
movimenti peristaltici corti sono invece relativi a piccoli tratti dell'intestino e vengono generati dalle cellule del
Kajal: questi sono limitati nella diusione in quanto le sinapsi elettriche vengono disaccoppiate tra loro, e il loro
ruolo è quello di spingere in avanti il cibo.
Nel colon la muscolatura si presenta in maniera totalmente diversa: si perdono due strati circolari distinti a
favore di tre bande muscolari, le tenie. Il risultato funzionale è la presenza di una motilità segmentaria che
non ha una funzione di propulsione, ma piuttosto di compattamento del materiale fecale. La progressione del
contenuto del colon è in realtà garantita dai movimenti di massa, che coinvolgono l'intero organo. La permanenza
del materiale nel colon è molto lunga, no a 48 ore, in modo tale da garantire il tempo di assorbire tutto quel che
è possibile.
A livello anale si trovano due snteri funzionalmente diversi tra loro; lo sntere anale interno è formato da
muscolatura liscia mentre lo sntere anale esterno da muscolatura striata sotto controllo volontario. Il riempimento del retto fa aumentare la pressione sullo sntere interno che risponde per via riessa con un rilassamento:
a trattenere le feci rimane solo lo sntere esterno. Se la defecazione non è possibile la contrazione dello sntere
anale esterno viene mantenuta e dopo un certo periodo anche lo sntere interno riprende tono e spinge le feci indietro; quando la defecazione è invece possibile basta rilasciare volontariamente anche lo sntere esterno e procurarsi
qualcosa da leggere.
2
Secrezione
Nel sistema GI arrivano ogni giorno liquidi di natura esogena nella quantità di due litri, e liquidi di natura endogena
nella quantità di sette litri; in particolare la suddivisione è la seguente:
• 2L tra cibi e bevande
• 1,5L di saliva
• 0,5L di bile
• 2L di succo gastrico
5
• 1,5L di succo pancreatico
• 1,5L di succo intestinale
La quantità totale di liquidi in circolo è dunque pari a circa nove litri, dei quali:
• 7,5L sono assorbiti dall'intestino tenue
• 1,4L sono assorbiti dall'intestino crasso
• 0,1L sono espulsi con le feci
2.1
La saliva
La saliva è prodotta dalle ghiandole parotide, sottomandibolare e sottolinguale, ma anche da ghiandole minori
di guance, lingua e labbra. L'unità funzionale di tutte queste ghiandole prende il nome di salivone e tutte sono
sotto esclusivo controllo nervoso. L'innervazione parasimpatica proviene dai nuclei salivatori che inviano
le loro bre tramite il nervo faciale (CNVII) per la sottomandibolare e la sottolinguale o tramite il nervo
glossofaringeo (CNIX); l'innervazione simpatica è legata invece ai primi segmenti toracici. Le funzioni principali
della saliva sono:
• La solubilizzazione dei cibi e la mitigazione delle pressioni osmotiche
• La lubricazione
• La fonazione
• L'abbattimento della carica batterica dei cibi
Dal punto di vista chimico la saliva è una soluzione acquosa (90%) di elettroliti e sostanze proteiche, in particolare:
•
Ptialina,o alfa amilasi, con ruolo di scissione degli amidi
•
Lipasi
•
Mucoproteine
•
Lisozima e anticorpi
La saliva è sempre ipotonica per poter abbassare le pressioni osmotiche dei cibi secchi o molto salati; l'osmolalità è
molto bassa quando la velocità di secrezione è minore, cioè lontano dai pasti, mentre nei momenti di grande richiesta
si abbassa pur non raggiungendo quella del plasma. Le concentrazioni delle quattro specie ioniche fondamentali
nella secrezione (sodio, cloro, potassio e bicarbonato) dipendono dalla velocità di secrezione; questo è legato al
fatto che tutte le secrezioni avvengono a due stadi. Il primo stadio di secrezione, a livello degli acini, produce un
secreto isotonico con il plasma; La secrezione primaria viene rimaneggiata lungo i dotti e si hanno due processi:
riassorbimento di sodio e cloro e secrezione di potassio e bicarbonato. Quando la velocità di secrezione aumenta
si ha meno tempo per riassorbire sodio e cloro e per secernere il potassio: il risultato è che l'osmolalità si alza.
Il bicarbonato è invece secreto attivamente e la sua concentrazione infatti aumenta all'aumentare della velocità di
secrezione: il pH passa da 6,4 a riposo a 8 al momento di picco.
Il sistema nervoso simpatico e parasimpatico hanno sulle ghiandole salivari essenzialmente lo stesso eetto:
entrambi inducono una secrezione, ma in modo diverso. Le principali dierenze sono:
Quantità Grande per via parasimpatica, scarsa se via simpatica
Durata Lo stimolo parasimpatico dura di più
Composizione La stimolazione parasimpatica produce una saliva con poche proteine e molto potassio e bicarbonato, l'opposto con una stimolazione simpatica
Denervazione La ghiandola denervata del parasimpatico diventa atroca, del simpatico riduce semplicemente la
secrezione
6
2.2
La secrezione gastrica
Il succo gastrico è una secrezione, per la maggior parte acquosa, contenente:
• Cationi: magnesio, potassio, sodio e soprattutto idrogeno
• Anioni: cloro, fosfato e solfato
• Pepsinee
• Lipasi gastrica
• Muco
• Fattore intrinseco
Il fattore intrinseco è l'unico componente indispensabile alla vita per quanto riguarda la secrezione gastrica:
individui privi di stomaco dipendono da iniezioni di vitamina B12. L'unità fondamentale per la secrezione di questa
soluzione è la ghiandola gastrica, la quale presenta almeno tre diverse tipologie cellulari; le cellule parietali
secernono HCl e fattore intrinseco, le cellule principali tutti gli enzimi in forma di zimogeni e le cellule mucose
glicoproteine con ruolo protettivo. Il succo gastrico è leggermente ipotonico e diventa isotonico con l'aumento della
velocità di secrezione; il pH è sempre molto basso e oscilla tra uno e tre: un ambiente simile è in grado di uccidere
quasi tutti i microorganismi in arrivo con il cibo. Gli ioni importanti sono il cloro e il sodio, con concentrazione
inferiore a quella plasmatica, e potassio e idrogeno, con concentrazione superiore a quella plasmatica. L'aumento
della velocità di secrezione segna una riduzione della presenza del sodio e un aumento delle altre tre specie, in
particolare del cloro: il succo gastrico diventa di fatto una soluzione di HCl.
L'HCl viene prodotto dalle ghiandole gastriche a partire da idrogenioni e ioni cloro. L'idrogenione proviene
dall'anidride carbonica derivante dal metabolismo cellulare: questa entra nella cellula dove viene idratata a dare
acido carbonico il quale viene scisso generando bicarbonato e idrogenione. L'idrogenione viene espulso nel lume
gastrico tramite una pompa H-K ATPasica mentre il bicarbonato viene espulso nel versate ematico scambiandolo
con uno ione cloro. Il cloro così ottenuto viene riversato nello stomaco per tramite di un canale dedicato. Alla ne
del processo sul versante gastrico si ha produzione di HCl mentre su quello ematico espulsione di bicarbonato: il
sangue reuo dallo stomaco secernente è infatti basico. Il pH acido così ottenuto contribuisce da un lato all'attività
microbicida e dall'altro all'attivazione del pepsinogeno in pepsina.
Il muco prodotto dalle cellule mucose ha un ruolo protettivo nei confronti della mucosa gastrica, altrimenti
sensibile al pH acido e alle pepsine attivate. Il lm mucoso che si viene a creare è fondamentale per la salute dello
stomaco; il suo continuo ricambio viene inibito dai farmaci anti inammatori non steroidei aspecici (FANS inibitori
sia a COX1 che a COX2) che a lungo andare producono infatti gastriti ed ulcere.
7
Le cellule parietali sono sotto controllo stretto lungo almeno tre vie: un meccanismo neurocrino, uno paracrino ed
uno endocrino. La via neurocrina vede terminazioni nervose colinergiche scaricare acetilcolina sui recettori
muscarinici delle cellule parietali. Il meccanismo paracrino si realizza grazie alle cellule enterocromani
simili che liberano istamina la quale raggiunge le cellule parietali e in particolare ai recettori H2. Il meccanismo
endocrino è legato invece alla secrezione duodenale di gastrina la quale è in grado di stimolare le cellule ECL
a secernere istamina o in alternativa di stimolare direttamente le cellule parietali. Questi tre diversi meccanismi
forniscono contributi diversi a seconda delle fasi: si individuano una fase cefalica, una gastrica ed una intestinale.
Nella fase cefalica gli stimoli sono la masticazione, la deglutizione, il gusto e l'odore dei cibi e inne i riessi
condizionati; questa fase, legata a meccanismi neurocrini, conta per il 35% del totale di succo gastrico prodotto.
Nella fase gastrica viene prodotto il 60-65% del succo totale e gli stimoli sono la distensione dello stomaco e la
presenza di prodotti di digestione delle proteine; i meccanismi sono riessi locali o vagovagali che stimolano le
cellule parietali grazie a istamina o gastrina. La fase intestinale è guidata da meccanismi endocrini e paracrini;
il sistema ruota intorno al pH del duodeno. Quando il pH del duodeno è superiore a tre prevalgono gli stimoli
eccitatori che inducono lo stomaco a secernere di più, mentre per valori inferiori la secrezione viene inibita.
2.3
Il succo pancreatico
L'unità funzionale del pancreas è il lobulo pancreatico con i suoi dotti convergenti che arrivano a formare il dotto
principale. Il pH del succo pancreatico è alcalino in quanto il componente più importante è il bicarbonato. Gli
enzimi contenuti sono in grado di digerire ogni principio alimentare e pertanto il succo viene denito completo.
Questi enzimi sono raggruppati a seconda del principio alimentare che attaccano: si hanno enzimi proteolitici,
amilolitici, lipolitici e anche nucleolitici. Il succo pancreatico è isoosmotico con il plasma a qualsiasi velocità di
secrezione, quello che cambia è il pH che tende a diventare marcatamente basico ad alte velocità di secrezione: il
bicarbonato viene secreto infatti in modo attivo. La secretina secreta dal duodeno in risposta ad un basso pH da un
lato inibisce lo svuotamento gastrico ma dall'altro stimola la secrezione pancreatica in modo da sfruttare il potere
tamponante di questa soluzione. Lo ione bicarbonato, principale componente e tampone della secrezione, viene
prodotto grazie al processo inverso a quello che portava alla formazione di HCl nello stomaco; poichè il processo è
esattamente inverso il sangue in uscita dal pancreas in secrezione è acido.
I meccanismi di secrezione pancreatica sono nuovamente divisi nelle tre fasi canoniche. La fase cefalica ha
un'importanza modesta (5-10%), risponde agli stessi stimoli validi per lo stomaco e agisce in modo analogo, cioè
con liberazione di ACh vagale. La fase gastrica è legata a stimoli di distensione gastrica e presenza di prodotti di
digestione: qui il meccanismo privilegiato è quello paracrino, con importante ruolo della gastrina. La fase intestinale
è quella più importante ed è legata all'acidità dell'ambiente intestinale soprattutto, ma anche alla presenza di
molecole quali gli acidi grassi che scatenano la secrezione di colecistochinina.
8
2.4
La bile
La bile è un secreto GI particolare in quanto non contiene enzimi e quindi ha un ruolo indiretto nei processi digestivi;
alcuni dei composti della bile sono comunque necessari per la digestione e l'assorbimento ottimale dei lipidi. La
secrezione di bile è anche la via di escrezione di sostanze esogene, quali metalli pesanti e farmaci, ed endogene,
quali i pigmenti biliari, il colesterolo ed i fosfolipidi; questa soluzione ha anche un ruolo di assorbimento nei
confronti delle vitamine liposolubili.
In termini chimici la bile è una secrezione acquosa contenente vari ioni (N a+ , Ca++ , K + , Cl− , HCO3− ) e composti
organici tra i quali i più importanti sono gli acidi biliari, i pigmenti biliari, il colesterolo ed i fosfolipidi. La bile viene
suddivisa in due tipologie: una bile epatica prodotta direttamente dal fegato e una bile cistica prodotta dalla
colecisti a partire da quella epatica. La bile cistica è fondamentalmente il risultato di un processo di concentrazione
e di riassorbimento di ioni e acqua. Il pH è nell'ambito della neutralità per quanto riguarda la bile cistica ed è
leggermente basico per la bile epatica (per via del bicarbonato, che invece nella cistica viene riassorbito); entrambe
le tipologie di bile sono isotoniche al plasma.
Gli acidi biliari derivano dal colesterolo e hanno un ruolo importante per la digestione e l'assorbimento dei
lipidi in quanto hanno la capacità di emulsionare queste molecole: i lipidi vengono così frammentati per orire
una vasta supercie di attacco per le lipasi. Questi acidi sono molecole anpatiche, cioè presentano una porzione
idrofobica e una idrolica, e quando presenti in concentrazioni adeguate tendono a formare micelle attorno ai lipidi.
Le micelle così formate arrivano a contatto con l'orletto a spazzola di duodeno e digiuno e vengono svuotate dei
contenuti; gli acidi biliari non vengono invece assorbiti a questo livello, ma verranno recuperati nell'ileo terminale.
9
A livello del lume intestinale i batteri presenti possono modicare gli acidi biliari, trasformandoli da primari a
secondari per aggiunta di gruppi ossidrilici. Il fegato è in grado di riconvertire gli acidi biliari secondari in primari,
e inoltre ha il ruolo di coniugare queste molecole con taurine e glicine che li rendono più stabili e ne prolungano la
vita in circolo. Il pool di acidi biliari presente in un dato momento ammonta a circa tre grammi ma è in grado di
circolare dalle quattro alle dodici volte e quindi il pool reale è compreso tra 12 e 36 grammi circa. Il fegato riceve
gran parte degli acidi biliari per ricircolo e li rimette in viaggio, per poi sostituire la piccola frazione che ogni giorno
viene persa tra feci ed urine.
I globuli rossi hanno una vita di poco più di cento giorni e vengono continuamente eliminati negli organi di
eritrocateresi. Il gruppo EME viene trasformato a livello del reticolo endoplasmatico in bilirubina, la quale
passa nel sangue associata ad un'albumina plasmatica. La bilirubina giunge così al fegato dove viene coniugata a
due molecole di acido glucuronico formando così il bilirubin-diglucuronide che viene secreto nella bile. La bile
nel suo passaggio a livello intestinale può essere rimaneggiata dai batteri che tendono a staccare l'acido glucuronico
e produrre bilirubina libera; altre popolazioni batteriche agiscono sulla bilirubina libera prodotta trasfromandola
in urobilinogeno, del quale una parte viene ulteriormente rimaneggiato a dare stercobilina. La stercobilina e
parte dell'urobilinogeno vengono escreti insieme al bilirubin diglucuronide nelle feci, mentre un'altra porzione di
urobilinogeno viene captato dal rene dove viene trasormato in urobilina ed escreto nelle urine.
10
La struttura secernente del fegato è simile a quella pancreatica e anche qui si ritrovano una secrezione primaria
ed una secondaria. La secrezione primaria viene rimaneggiata nei dotti dove vengono riassorbiti sodio e cloro e
quindi anche acqua per osmosi. Se lo sntere di Oddi è chiuso, quindi lontano dai pasti, la bile prodotta non arriva
all'intestino ma viene ad acccumularsi nella colecisti. La colecisti ha una capacità di 30 − 60mL mentre il fegato
secerne oltre un litro di bile al giorno: esistono dunque dei meccanismi di concentrazione legati all'assorbimento di
sodio, cloro e bicarbonato e quindi di acqua. La concentrazione della bile avviene secondo un meccanismo detto di
gradiente osmotico stazionario.
La contrazione della colecisti e quindi il rilascio di bile è un fenomeno legato a tre diverse fasi: cefalica, gastrica
ed intestinale. Nelle prime due fasi il meccanismo è di natura nervosa e legato a riessi vagovagali. Nella fase
intestinale è la presenza di lipidi ad indurre la liberazione di colecistochinina la quale potenzia la secrezione
biliare, quella pancreatica e inibisce lo svuotamento dello stomaco.
2.5
Il succo intestinale
L'intestino contribuisce alle secrezioni GI con un volume di circa un litro e mezzo. Il succo intestinale è una soluzione
isotonica di acqua ed elettroliti, la quantità di enzimi è molto modesta mentre quella di muco è rilevante. Gli stimoli
alla secrezione sono gli stessi delle regioni precedenti del tratto GI.
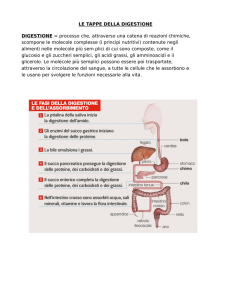
3
Digestione ed assorbimento
La maggior parte del lavoro di digestione ed assorbimento è svolta dalla prima porzione dell'intestino tenue, dunque
duodeno e digiuno prossimale. La supercie assorbente raggiunge circa 200m2 grazie alla presenza dei villi e dei
microvilli: questo accorgimento permette di ottenere un assorbimento di quasi il 90% di quanto ingerito. Le sostanze
ingerite possono essere divise in tre gruppi fondamentali: carboidrati, proteine e lipidi. I processi di digestione sono
da dividersi in una digestione meccanica, operata a livello della bocca e dello stomaco, e una digestione chimica,
operata dai vari enzimi lungo più o meno tutto il tratto GI. La digestione chimica è un fenomeno che avviene per
stadi: si distinguono una digestione luminale, una digestione di membrana ed eventualmente una digestione
intracellulare.
11
3.1
I carboidrati
I carboidrati introdotti con la dieta possono avere origine animale, quindi polimeri di glicogeno, o origine vegetale,
quindi polimeri di amilopectine (amido). I legami di queste molecole rientrano in tre categorie:
•
Alfa 1-4 glicosidici
•
Beta 1-4 glicosidici
•
Alfa 1-6 glicosidici (unico legame ramicato)
La digestione luminale avviene ad opera delle alfa amilasi deramicanti della saliva o del succo pancreatico; questi
enzimi scindono selettivamente il solo legame 1-4 glicosidico e lasciano interi gli altri due. L'azione delle amilasi
produce una miscela di maltosio, maltotriosio e di polimeri contenenti il legame 1-6 detti destrine alfa-limite.
La digestione di membrana chiama in causa enzimi quali l'alfa destrinasi (o isomaltasi) che scindono i legami di
tipo 1-6, e altri enzimi (glucoamilasi, saccarasi) che digeriscono i vari prodotti no ad arrivare a monosaccaridi
assorbibili. I monosaccaridi così prodotti vengono assorbiti dagli epatociti per tramite di trasportatori, alcuni
dei quali sono sodio dipendenti (SGLT1) e altri indipendenti (GLUT5). Tutti questi meccanismi avvengono nel
duodeno e nella prima parte del digiuno: in condizioni siologiche la digestione e l'assorbimento dei carboidrati
sono completi.
3.2
Le proteine
Le proteine vengono degradate dagli enzimi proteolitici che nell'uomo iniziano ad essere presenti solo a livello
dello stomaco. Questi enzimi vengono secreti sotto forma di pepsinogeni, quindi zimogeni inattivi, e attivati dal
basso pH gastrico. Le varie pepsine sono in grado di scindere esclusivamente i legami peptidici interni alle catene
polipeptidiche, quelli terminali vengono lasciati intatti: il risultato è la produzione di peptidi più o meno lunghi.
La digestione proteica è dunque sempre incompleta. I prodotti di digestione delle pepsine vengono rimaneggiati
a livello dell'orletto a spazzola da proteine integrali di membrana le quali producono aminoacidi singoli o piccoli
peptidi; a livello della membrana degli eritrociti esistono dei trasportatori specici per aminoacidi singoli o peptidi
di due, tre o quattro elementi. Il citosol degli enterociti è la sede per la terza fase di digestione, cioè la riduzione a
singoli aminoacidi di quanto assunto con i trasportatori: alla ne i singoli elementi vengono espulsi dalla cellula e
immessi nel liquido interstiziale.
3.3
I lipidi
I lipidi sono un gruppo eterogeneo di sostanze idrofobiche bersaglio degli enzimi lipolitici (lipasi). Questi enzimi
scindono il legame estere dei trigliceridi e quello dei fosfolipidi. Le lipasi si dividono in preduodenali, cioè gastriche
e salivari, e pancreatiche. La bile ha un ruolo emulsionante nei confronti dei lipidi, aumenta cioè la supercie
di attacco per le lipasi grazie alla formazione delle micelle. I monogliceridi, i lipofosfatidi ed il colesterolo ottenuti
vengono assorbiti a livello dell'orletto a spazzola e convogliati al reticolo endoplasmatico liscio dove viene operata
una risintesi dei legami esteri. Questi trigliceridi di sintesi endogena vengono poi impacchettati dal Golgi in strutture
dette chilomicroni e legati alle apolipoproteine. I chilomicroni hanno dimensioni troppo importanti per entrare nei
capillari sanguigni e infatti passano nei vasi linfatici i quali hanno giunzioni molto più lasse.
12