…ogni cellula deriva da
un’altra cellula…
“Omnis
cellula e cellula”
Virchow 1858
LA MAGGIOR PARTE DEI
PROCESSI NUCLEARI E
CELLULARI DURANTE LA
MITOSI E’
INDISTINGUIBILE IN
PIANTE,ANIMALI E
FUNGHI.
DIVISIONE CELLULARE IN
UN BATTERIO.
L’ATTACCO DEL CROMOSOMA
A UNA INTROFLESSIONE
DELLA MEMBRANA
PLASMATICA ASSICURA CHE
QUEL CROMOSOMA,
DUPLICATO, VENGA
DISTRIBUITO A CIASCUNA
DELLE CELLULE FIGLIE,
QUANDO LA MEMBRANA
PLASMATICA SI ACCRESCE.
INTERFASE
ANAFASE
PROFASE
TELEOFASE
METAFASE
Legenda: sinistra foto microscopio elettronico;
destra rappresentazione schematica
La capacità di moltiplicarsi è una
…perchè assicurano la conservazione
proprietà fondamentale delle cellule,
del materiale ereditario e la sua
così come quella di riprodursi lo è
trasmissione da una generazione
per ogni organismo vivente. Negli
all’altra. La replicazione del DNA, la
eucarioti la duplicazione dei
duplicazione dei cromosomi e la loro
cromosomi e la divisione cellulare
ripartizione in due nuclei distinti, sono
rappresentano processi
processi che si verificano durante il
fondamentali….
ciclo cellulare.
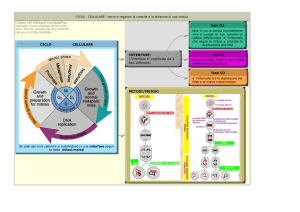
PERCHÈ LE
CELLULE
SI DIVIDONO?
Per la riproduzione
Per l’accrescimento
Per la riparazione
CICLOdurasse
CELLULARE
Se il cicloIL
cellulare
1 ora: È UNA SERIE ORDINATA DI
In realtà
durata del ciclo
è diversa
la fase G1
durerebbe
minuti
EVENTI
CHE50DETERMINANO
LAlaCRESCITA
DELLA
per ogni cellula; inoltre dipende da
circa e tutte le altre fasi insieme i
CELLULA E LA SUA DIVISIONE IN DUE CELLULE
molti fattori quali la temperatura e
successivi 10 minuti.
FIGLIE.
le sostanze nutritive che la cellula ha
a disposizione.
La maggior parte delle
attività di una cellula si
svolgono durante la fase G1
Alcune cellule, come le cellule
Nervose, smettono di moltiplicarsi dopo
Lo sviluppo embrionale.
In questi casi la fase G1 dura per tutta
La vita dell’individuo e viene detta fase G0
INTERFASE
(G1 + S + G2)
In questa fase avviene l’accrescimento cellulare con
• Attività metabolica
• Sintesi proteica
• Sintesi di DNA
G1 (Gap): la cellula è in intensa attività
metabolica, aumenta le sue dimensioni,
raddoppia gli organuli cellulari e si procura
il materiale per la sintesi del DNA
S (sintesi): avviene la duplicazione del DNA e delle proteine ad esso
associate: il cromosoma duplicato adesso è costituito da due cromatidi uniti
in una regione chiamata centromero
G2 (Gap): preparazione alla mitosi con sintesi di proteine
per la divisione cellulare
ALLA FINE DELL’INTERFASE LA
CELLULA APPARE COSÌ.....
Finchè la cellula
eucariotica non si divide,
il DNA è una “matassa” di
filamenti sottili chiamati
cromatina
DNA
umano
DNA umano in ciascuna
cellula:
è lungo complessivamente
2,5 m
è ripartito in 46 cromosomi
DNA di ciascun cromosoma:
è lungo diversi cm
è spesso 2 nm
G1
Interfase
S
G2
PROFASE
Ciclo cellulare
METAFASE
Mitosi
Divisione
cellulare
ANAFASE
TELOFASE
Citodieresi (divisione del citoplasma)
Piastra
metafasica
Cromosoma, cromatidi, centromero, telomeri.
Un cromosoma:
• prima di duplicarsi, è costituito da un cromatidio
• dopo la replicazione del DNA è formato da due cromatidi
“fratelli”, che appaiono uniti nella regione del centromero
• Quando i due cromatidi si separano, si avranno due nuovi cromosomi
uguali (ciascuno costituito da un cromatidio)
Schema di un cromosoma
completamente
condensato.
Il centromero, l’area
ristretta al centro, è il
punto di unione dei due
cromatidi.
I cinetocori, uno per
ciascun cromatidio , sono
Ai cinetocori si legano i
strutture contenenti
microtubuli che formano
proteine associate con il
parte del fuso.
centromero.
METAFASE
PIASTRA
METAFASICA
Stato di ciascun
cromosoma
Organizzazione
della cellula
Separazione
CITODIERESI
LA MITOSI
•
Alcuni ruoli svolti dai complessi CDK-ciclina nel ciclo cellulare. Sia laa replicazione del DNA durante la
fase S sia l’assemblaggio del fuso mitotico nella fase M sono controllati da specifici complessi CDKciclina che si assemblano nei momenti appropriati del ciclo cellulare
•
Distribuzione e assemblaggio dei complessi CDK-ciclina nel ciclo cellulare. Complessi proteolitici
specializzati si legano a cicline specifiche, che sono attive in momenti diversi del ciclo cellulare. La
degradazione di queste cicline rimuove l’attività dello specifico complesso CDK-ciclina, promuovendo la
transazione della cellula alla fase successiva del ciclo. Il complesso proteolitico è formato da diverse
proteine necessarie per il riconoscimento specifico delle proteine bersaglio.. Un meccanismo che controlla
l’azione dei complessi proteoliti è la fosforilazione proteica. Il complesso SCF che ha per bersaglio la ciclina
G1 non riconosce questa ciclina fino a quando essa non sia stata fosfolrilata da una chinasi. I complessi
proteolitici possono avere come bersagli più tipi di proteine. Ad esempio, il complesso ACP degrada anche
proteine che sono essenziali in una fase precoce della mitosi, consentendo alle fasi successive di realizzarsi.
•
La via WEE/CDC25 nel lievito. La fosforilazione della chinasi ciclina-dipendente
(CDK) che promuove il passaggio della fase G2 alla fase M del ciclo cellulare del
lievito inibisce l’attività di questa CDK, impedendo la transizione di fase. La
defosforilazione, che porta alla riattivazione della CDK, permette la transizione e
quindi consente al ciclo di proseguire. La fosforilazione e la defosforilazione sono
catalizzate, rispettivamente dalla chinasi WEE e dalla fosfatasi CDC25.
•
.
Regolazione della CDK che promuove la transizione da G2 a M Questa chinasi
ciclina-dipendente (CDK) può essere inibita dal legame con una proteina inibitrice,
oltre che dalla fosforilazione della subunità catalitica. L’inibizione ooperata da SIC!,
un inibitore di CDK, è annullata dalla fosforilazione di SIC1 da parte di una chinasi.
La fosforilazione consente a SIC1 di essere riconosciuto e degradato dal complesso
proteolitico SCF, in un modo analogo alla degradazione della ciclina G1 .
Via metabolica della proteina del retinoblastoma. La proteina del retinoblastoma (Rb) è
parte del meccanismo che controlla la transizione da G1 a S. La replicazione del DNA
nella fase S è attivata dal fattore di trascrizione E2F. Prima della fase S, nella fase G1.
l’azione di E2F è inibita dal suo legame con Rb. Il ruolo della CDK attiva durante la
transizione da G1 a S è quello di fosforilare Rb, ciò infatti causa il rilascio di Rb da E2F e
permette a E2F di attivare la sintesi del DNA. Anche la stessa CDK è soggetta a diversi
livelli di regolazione. Come mostra la figura, la sua attività dipende dalla fosforilazione
della subunità catalitica ( mediante la chinasi CAK), ed è inibita dal legame con un
inibitore della CDK.
Checkpoint nel ciclo cellulare. Il completamento della replicazione del DNA è necessario
perché arrivino a completamento negli stadi finali del ciclo cellulare. Nel lievito, se la
chinasi « checkpoint 1» (CDK1) individua che la replicazione del DNA non è completa,
essa previene la transizione alla fase M agendo sulle proteine che determinano l’attività
della CDK necessaria per questa transizione. CHK1 attiva la chinasi WEE e inibisce la
fosfatasi CDC25, cosi la subunità catalitica di CDK rimane nella sua forma fosforilata,
inattiva.
MEIOSI
MEIOSI
DIFFERENZE MITOSI -MEIOSI
mitosi
meiosi
Per effetto
del
crossing-over,
le 4 cellule
risultanti
dalla meiosi
avranno 4
cromatidi
diversi
FINE