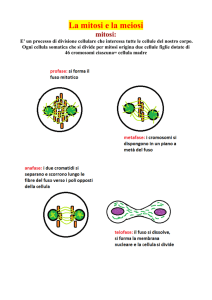
20/04/16 IL CICLO CELLULARE Tu#e le cellule derivano da altre cellule Le immagini presen2 sono prese dal libro “Biologia molecolare della cellula” Alberts et al, Zanichelli La divisione cellulare e il ciclo cellulare. La riproduzione cellulare inizia con la duplicazione del contenuto della cellula, seguita dalla distribuzione di questo contenuto nelle cellule figlie. La duplicazione dei cromosomi avviene durante la fase S del ciclo cellulare, mentre la maggior parte degli altri componen2 cellulari sono duplica2 con2nuamente durante il ciclo. Durante la fase M, i cromosomi replica2 sono segrega2 in nuclei individuali (mitosi) e la cellula poi si divide in due (citochinesi). La fase S e la fase M sono di solito separate da fasi gap chiamate G1 e G2, in cui la progressione del ciclo cellulare può essere regolata da vari segnali intra-­‐
extracellulari. Le fasi del ciclo cellulare. La cellula cresce in modo con2nuo in interfase, che consiste di tre fasi: la replicazione del DNA è confinata nella fase S; G1 è l’intervallo tra la fase M e la fase S, mentre G2 è l’intervallo fra la fase S e la fase M. Nella fase M il nucleo e quindi il citoplasma si dividono. 1 20/04/16 sequenza di even, che si ripete ogni 18-­‐24 ore Fase G1 in genere occupa tra le 6 e le 12 ore nelle cellule che crescono più rapidamente. Fase S dura 6-­‐ 8 ore Fase G2 è la fase più corta dell’interfase Fase M è di circa 1 ora. La durata del ciclo varia molto a seconda del tipo cellulare. 2 20/04/16 Pun8 di controllo del sistema di controllo del ciclo cellulare. Informazioni sul completamento degli even2 del ciclo cellulare, oltre che segnali provenien2 dall’ambiente, possono provocare l’arresto del ciclo da parte del sistema di controllo in corrispondenza di pun2 di controllo specifici. I pun2 di controllo operano generalmente aXraverso segnali intracellulari nega2vi, che arrestano il ciclo cellulare, invece che aXraverso la rimozione dei segnali posi2vi che normalmente s2molano la progressione del ciclo. Pun8 di controllo del sistema di controllo del ciclo cellulare. G1 checkpoint: controllo delle dimensioni cellulari, della presenza di un ambiente favorevole G1/S checkpoint: controllo dell’assenza di danni genetici, es. radiazioni ionizzanti possono indurre rotture nel DNA G2 checkpoint: controllo delle dimensioni, della completa duplicazione e dell’integrità di tutto il materiale genetico. È durante questo checkpoint che vengono riparati gli eventuali danni rilevati al DNA. M checkpoint: controllo dell’assemblamento corretto dei cromosomi al fuso mitotico 3 20/04/16 Il sistema di controllo del ciclo si basa su proteina chinasi a?vate ciclicamente. La Cdk si associa in successione con cicline diverse per scatenare i diversi even2 del ciclo. L’a[vità Cdk è in genere terminata dalla degradazione della ciclina. cicline D agiscono nei momenti iniziali della fase G1; si legano con le CdK4 e 6; il loro compito è quello di guidare la transizione tra la fase G1 e la fase S. cicline E, invece, agiscono più tardivamente nella fase G1 legandosi alle Cdk2. Anche il loro legame è richiesto per la transizione dalla fase G1 alla fase S. cicline A si legano alle Cdk1 e Cdk2; queste cicline stimolano la duplicazione dei cromosomi e partecipano alla regolazione delle prime fasi della mitosi. cicline B sono le cicline mitotiche e formano un complesso con le CdK1. Queste cicline stimolano l'ingresso nella mitosi. 4 20/04/16 L’a?vità della Cdk può essere soppressa sia da fosforilazione inibitrice che da proteine inibitrici La regolazione dell’a?vità Cdk da parte di fosforilazione inibitrice. Il complesso ciclina-­‐Cdk è spento quando la chiasi Wee1 fosforila due si2 molto vicini sopra il sito a[vo. La rimozione di ques2 fosfa2 da parte della fosfatasi Cdc25 porta all’a[vazione del complesso. L’inibizione di un complesso ciclina-­‐Cdk da parte di una CKI. La p27 si lega sia alla ciclina che alla Cdk nel complesso, distorcendo in sito a[vo della Cdk. Si inserisce anche nel sito che lega ATP, inibendo ulteriormente l’a[vità dell’enzima. Il controllo della proteolisi da parte di SCF e APC durante il ciclo cellulare La fosforilazione di una proteina bersaglio come la CKI, permeXe alla proteina di essere riconosciuta da SCF, che è cos2tu2vamente a[va . Con l’aiuto di due proteine addizionali E1 e E2, SCF serve da ubiqui2na ligasi che trasferice molecole mul2ple di ubiqui2na sulla proteina CKI, questa viene immediatamente riconosciuta e degradata in un proteasoma. L’ubiqui2nazione della ciclina M è aXuata da APC, che è a[vata dall’aggiunta di una subunità a[vatrice al complesso. Sia SCF che APC contengono si2 di legame che riconoscono sequenze specifiche di amminoacidi nella proteina bersaglio. 5 20/04/16 Complessi ciclina nella fase S-­‐Cdk iniziano la replicazione del DNA una sola volta per ciclo. La replicazione deve avvenire con estrema accuratezza per minimizzare il rischio di mutazioni nella successiva generazione cellulare, ma ogni nucleo2de del genoma deve essere replicato una volta soltanto per i m p e d i r e g l i e ff e [ d a n n o s i dell’amplificazione genica. La replicazione del DNA inizia a livello di origini di replicazione, che sono sparse in varie posizioni nel cromosoma. ORC resta associato ad un origine per tuXo il ciclo. All’inizio di G1 Cdc6 si associa ad ORC. Recluta2 da Cdc6, complessi Mcm si assemblano quindi sul DNA adiacente. Si forma il complesso prereplica2vo. S-­‐Cdk scatena quindi l’accensione dell’origine assemblando DNA pol e altre proteine di replicazione. Inoltre blocca anche la replicazione provocando la dissociazione di Cdc6 dalle origini, la sua degradazione e l’esportazione di MCm in eccessoi nel nucleo. Cdc6 e MCm non possono ritornare a ristabilire un origine contenente ORC per un altro ciclo cellulare finchè M-­‐Cdk non viene a[vata alla fine della mitosi. MITOSI La mitosi è il processo di riproduzione effeXuato esclusivamente dalle cellule eucario2che. Il termine mitosi deriva dal greco mìtos, "filo": nome dovuto all'aspeXo filiforme dei cromosomi durante la metafase. La mitosi è molto simile alla meiosi, ma si dis2ngue da essa poiché forma 2 cellule diploidi con lo stesso numero di cromosomi della cellula madre, mentre nella meiosi si formano 4 cellule aploidi con il corredo cromosomico dimezzato. La mitosi riguarda le cellule soma2che dell'organismo (ossia tuXe le cellule fuorché quelle che hanno funzione riprodu[va: i gametoci2 primari, i quali u2lizzano la meiosi) e le cellule germinali ancora indifferenziate. La mitosi si divide in varie fasi chiamate: • profase • prometafase • metafase • anafase • telofase 6 20/04/16 FASI DELLA MITOSI Profase: In profase, i cromosomi replica2, ciascuno consistente di due croma2di fratelli streXamente associa2, si condensano. Fuori dal nucleo, il fuso mito2co si assembla fra i due centrosomi, che si sono replica2 e separa2. FASI DELLA MITOSI Prometafase: La prometafase inizia all’improvviso, con la demolizione dell’involucro nucleare. I cromosomi possono ora aXaccarsi ai microtubuli del fuso tramite i loro cinetocori ed esercitare un movimento a[vo. 7 20/04/16 FASI DELLA MITOSI Metafase: In metafase, i cromosomi sono allinea2 all’equatore del fuso, a metà strada fra i poli del fuso. I microtubuli del cinetocore aXaccano i croma2di fratelli a poli oppos2 del fuso. FASI DELLA MITOSI Anafase: In anafase, i croma2di fratelli si separano in modo sincrono a formare due cromosomi figli e ciascuno di essi viene 2rato lentamente verso il polo del fuso che lo fronteggia. I microtubuli del cinetocore si accorciano e anche i poli del fusosi separano: entrambi i processi contribuiscono alla separazione dei cromosomi. 8 20/04/16 FASI DELLA MITOSI Telofase: Durante la telofase, le due serie di cromosomi figli arrivano ai poli del fuso e si decondensano. Un nuovo involucro nucleare si riassembla intorno a ciascuna serie, completando la formazione di due nuclei e segnando la fine della mitosi. La divisione del citoplasma inizia con l’assemblaggio dell’anello contra[le. FASI DELLA MITOSI Citochinesi: Durante la citochinesi, il citoplasma è diviso in due da un anello contra[le di filamen2 di ac2na e di miosina, che stringe la cellula e la divide in due per creare due cellule figlie, ciascuna con un nucleo. 9 20/04/16 L’a?vazione dei complessi M-­‐Cdk scatena l’ingresso in mitosi. Funzioni M-­‐Cdk: -­‐ Condensazione cromosomi -­‐ demolizione involucro nucleare -­‐ Riarrangiamento citoscheletro -­‐ Riorganizzazione del Golgi e del RE L’a[vazione della M-­‐Cdk inizia con l’accumulo di ciclina M (B) che aumenta durante la fase G2. questo aumento porta ad un accumulo graduale di M-­‐Cdk quando la cellula si avvicina alla mitosi. Il complesso M-­‐Cdk è fosforilato su un sito a[vatore dalla chinasi che a[va Cdk (CAK) e su una coppia di si2 inibitori dalla chinasi Wee1. Il complesso ina[vo che ne deriva viene quindi a[vato alla fine di G2 dalla fosfatasi Cdc25. Cdc25 è s2molata in parte dalla chinasi Polo, e dalla stessa M-­‐Cdk a[va, il che porta ad un feedback pos2vo. Questo feedback è aumentato dalla capacità di M-­‐Cdk di inibire Wee1. M-­‐Cdk prepara i cromosomi duplica8 per la separazione dei croma8di scatenata da proteolisi Il punto di controllo dell’aPacco al fuso Il punto di controllo opera per assicurare che tu[ i cromosomi siano aXacca2 in modo appropriato al fuso prima che avvenga la separazione dei croma2di fratelli. Il meccanismo sensore monitora lo stato del cinetocore,regione che si aXacca ai microtubuli del fuso. Qualunque cinetocore che non è aXaccato in modo appropriato al fuso manda un segnale nega2vo al sistema di controllo bloccando l’a[vazione di Cdc20-­‐APC e la separazione dei croma2di fratelli. L’a[vazione di APC da parte di Cdc20 porta all’ubiqui2nazione della securina che normalmente 2ene la separasi ina[va. La distruzione della securina permeXe alla separasi di tagliare una subunità del complesso delle coesine che 2ene insieme i croma2di fratelli. Le forze traen2 del fuso mito2co separano allora i croma2di fratelli. 10 20/04/16 La progressione nel ciclo cellulare è bloccata dal danneggiamento del DNA e dalla p53: pu8 di controllo del danno al DNA. Quando il DNA è danneggiato, vengono a[vate proteince chinasi che fosforilano p53. Mdm2 normalmente si lega a p53 e promuove la sua distruzione nei proteasomi. La fosforilazione di p53 blocca il suo legame a Mdm2; come risultato, la p53 si accumula ad al2 livelli e s2mola la trascrizione del gene che codifica la proteina CKI p21. La p21 si lega ai complessi g1/S-­‐Cdk e S-­‐Cdk e li ina[va, arrestando la cellula in G1. in alcuni casi il danno al DNA induce anche la fosforilazione di MDm2 o una diminuzione della sua produzione, che provoca un aumento di p53. MEIOSI La meiosi è un processo caraXeris2co delle cellule eucariote, essa riguarda unicamente la produzione delle cellule sessuali o game7 degli organismi pluricellulari. Con la meiosi, aXraverso un processo piuXosto complesso, una singola cellula diploide, dopo aver replicato una sola volta il suo DNA, da origine a quaXro cellule figlie, i game2 appunto, dotate di un patrimonio dimezzato di cromosomi e deXe perciò aploidi. La meiosi si differenzia dall'altro processo di divisione cellulare, la mitosi, nella quale si formano due cellule figlie aven2 lo stesso patrimonio gene2co della cellula madre. 11 20/04/16 PRIMA DIVISIONE MEIOTICA Nella prima divisione meio2ca si evidenziano i cromosomi, ciascuno cos2tuito da due croma2di. Ques2 cromosomi (metà di origine paterna e metà di origine materna), dopo aver subito alcuni processi durante la profase (in par2colare il crossing-­‐over, di cui parleremo successivamente), si portano al piano equatoriale della cellula. Qui, senza dividersi nei due croma2di, si aXaccano alle fibre del fuso per migrare verso i due poli in modo tale che, di ogni coppia di cromosomi omologhi, una si dirige verso un polo e l'altra al polo opposto. A conclusione della prima divisione meio2ca, si hanno così due cellule, ciascuna con la metà esaXa dei cromosomi omologhi. PRIMA DIVISIONE MEIOTICA PROFASE I : La croma2na visibile nel nucleo cellulare, che rappresenta la massa del DNA quando la cellula svolge le sue normali a[vità metaboliche, si condensa, in modo che si formano struXure bastoncellari, i cromosomi. Ciascun cromosoma appare a forma di X, poiché è formato da due croma2di fratelli, uni2 in un punto deXo centromero. I croma2di derivano da un processo di duplicazione del DNA; pertanto, ciascuno è gene2camente iden2co all’altro. In questa fase, una volta che i due cromosomi omologhi sono uni2 tra di loro, possono avvenire scambi incrocia2 di par2 più o meno lunghe di croma2di omologhi (fenomeno di crossing-­‐over). La membrana che avvolge il nucleo si disgrega. Si forma un fascio di microtubuli proteici, che si estende da un polo all’altro della cellula e le cui due estremità fanno capo a due coppie di organuli, de[ centrioli. METAFASE I : Le tetradi omologhe si dispongono simmetricamente lundo una linea immaginaria, trasversale rispeXo al fuso. In tal modo, ognuna è rivolta verso uno dei due poli della cellula. ANAFASE I : Le fibre del fuso prendono contaXo con i centromeri; ciascuna tetrade migra verso un polo della cellula. TELOFASE I : Ai due poli della cellula madre si formano due agglomera2 di cromosomi aploidi, in cui è presente un solo cromosoma per ciascun 2po. I cromosomi sono ancora allo stadio della tetrade. Il citoplasma delle due cellule si ripar2sce e avviene la citodieresi, ossia la vera e propria divisione della cellula originaria in due cellule figlie dis2nte (in alcuni casi, la ripar2zione può essere incompleta). Le fibre del fuso si disgregano; i cromosomi si despiralizzano. 12 20/04/16 SECONDA DIVISIONE MEIOTICA La seconda divisione meio2ca non è preceduta da alcuna duplicazione del DNA. I cromosomi, cos2tui2 da due croma2di, si portano all'equatore e si aXaccano alle fibre del fuso; i due croma2di di ciascun cromosoma si separano migrando ai poli. Si formano così quaXro cellule, ciascuna con un corredo aploide di cromosomi e con un diverso assor2mento dei cromosomi di origine materna e paterna. Durante questa separazione vi è una distribuzione indipendente dei cromosomi paterni e materni per cui, alla fine, vi sarà un diverso assor2mento dei cromosomi nelle quaXro cellule figlie. SECONDA DIVISIONE MEIOTICA PROFASE II : La croma2na si condensa nuovamente, in modo che si possono osservare i cromosomi, forma2 da due croma2di uni2 dal centromero. Si forma nuovamente il fuso di microtubuli. METAFASE II : I cromosomi si dispongono su una linea equatoriale, trasversale rispeXo alle fibre del fuso, in modo che ciascun croma2dio sia rivolto verso uno dei due poli della cellula. I centromeri prendono contaXo con le fibre. ANAFASE II : I croma2di migrano ciascuno verso un polo della cellula, spostandosi verso le fibre del fuso. In tal modo, ciascun croma2dio diviene un nuovo cromosoma. TELOFASE II : Ai poli della cellula, si formano due aggrega2 di cromosomi, le fibre del fuso si disgregano, i cromosomi cominciano a decondensarsi, e si forma infine una membrana nucleare. Il citoplasma della cellula si divide in due, cosi da portare alla formazione di due cellule figlie aploidi. 13 20/04/16 Morte cellulare programmata (apoptosi) EVENTI CARATTERISTICI DELL’APOPTOSI 1. Il citoplasma inizia a contrarsi in seguito alla proteolisi di filamenti di actina. Perdita d’acqua, insolubilizzazione proteine citoplasmatiche.. 2. La cromatina viene degradata in modo regolare (laddering) e così pure le proteine nucleari; il nucleo si condensa. 3. La cellula continua a contrarsi riducendosi ad una forma che permetta la facile eliminazione da parte dei macrofagi. Si verificano cambiamenti della membrana favorenti la fagocitosi 4. Formazione dei corpi apoptotici. 14 20/04/16 A?vazione extracellulare Un linfocita killer che porta il ligando FAS, si lega, a[vandole , a proteine Fas sulla cellula bersaglio. Proteine adaXatrici si legano alla regione intracellulare delle proteine Fas aggregate provocando l’aggregazione di molecole procaspasi-­‐8. Queste allora si tagliano a vicenda per iniziare la cascata delle caspasi. A?vazione intracellulare I motocondri rilasciano il citocromo c, che si lega, facendole aggregare, a molecole di procaspasi-­‐9, il che porta al taglio di queste molecole e allo scatenamento della cascata delle caspasi. Altre proteine che contribuiscono all’apoptosi sono anch’esse rilasciate dallo spazio mitocondriale intermembrana. 15 20/04/16 Il recettore Fas è uno degli inneschi principali dell’apoptosi La via estrinseca si basa su un sistema classico di interazione recettore-­‐ligando che, una volta attivato, determina l’interazione di più pro-­‐caspasi con alcune proteine attivatrici. Questi recettori di morte appartengono ad una famiglia genica che comprende recettori simili al TNF-­‐R (Recettore per il Tumor Necrosis Factor) tra i quali il più studiato in termini di segnalazione apoptotica è il Fas. Il Fas è costituito da: •un dominio extracellulare interazione con ligando specifico •un dominio trans-­‐membrana •dominio di morte (DD) regione citoplasmatica presente sulla membrana plasmatica di tutte le cellule, mentre il suo ligando, FasL, è espresso solo da tipi cellulari specifici. 16 20/04/16 FasL induce l’aggregazione del recettore successivamente la proteina FADD si lega al dominio di morte del recettore mediando il reclutamento del proenzima Caspasi-­‐8 presso il recettore. L’avvicinamento di più Pro-­‐Caspasi-­‐8 stimola la dimerizzazione di due pro-­‐
enzimi con il conseguente cambio conformazionale richiesto per l’innesco dell’attività proteolitica. p53 proteina soppressore del tumore (da tumor suppressor gene) in quanto se mutata consente la formazione di foci tumorali. ha il compito di controllare (G1) se sono avvenute mutazioni od alterazioni a carico del DNA. In tal caso, la p53 provvede a bloccare il ciclo specialmente perche', da fattore di trascrizione induce la sintesi ex novo di p21 (una CIP/KIP cdk inhibitor) che blocca il complesso Ciclina D/cdk4,6 (ed altri della stessa classe). p21 inoltre blocca PCNA, proteina necessaria all'assemblaggio della pinza di sostegno richiesta dalla DNA polimerasi durante la duplicazione del DNA. Se il danno al DNA e' troppo grave, p53 manda la cellula in apoptosi 17