IL CONCETTO DI SPECIE, RAZZA E TIPI Aspetti evolutivi e metodologici Marcello Andriola
Keywords: Specie, speciazione, mutazione, razza, razzismo, la genetica dei mutamenti.
Permalink: http://www.neuroscienze.net/?p=1195
Premessa
Problema tra i più controversi del nostro passato e del nostro presente, la questione dell'esistenza o
meno di sottodivisioni sostanziali della specie umana, chiamate spesso impropriamente «razze»,
costituisce tuttora motivo di scontro ideologico e politico. Tralasciando una qualsiasi speculazione
propriamente storica del divenire umano, mi propongo con questa ricerca di porre punti fermi per
l'indagine dell'annoso problema da un punto di vista meramente biologico (e più esattamente delle
radici biologiche) sulla base delle conoscenze scientifiche più recenti, e di riconoscere se sia lecita o
meno, e che limiti abbia, la fondazione di una teoria e di una prassi politica basata sul dato
biologico quanto più possibile esatto e senza interferenze di natura ideologica.
1
Il concetto di specie e razza: aspetti evolutivi e metodologici 1 Introduzione 3 1.1 Specie 7 1.2 Identificazione di una specie 7 1.3 Speciazione 8 1.4 Modalità di speciazione 8 1.5 Speciazione allopatrica 8 1.6 Isolamento riproduttivo 9 1.7 Speciazione simpatrica 9 1.8 Speciazione parapatrica 9 1.9 La specie ed i gruppi intraspecifici 9 1.10 L'evoluzione per equilibri intermittenti 11 1.11 La speciazione quantica e l'uomo 14 2.1 Razza: Evoluzione del concetto di razza 18 2.2 Razza e razzismo 18 2.3 Razze vegetali e animali 19 2.4 I geni e la storia 19 2.5 Altre origini del razzismo 20 2.6 Esiste una base scientifica del razzismo? 21 2.7 La natura delle differenze 22 2.8 Mutazioni visibili e invisibili 22 2.9 Razzismo 25 2.10 Le origini del razzismo 25 2.10.1 Dalla teoria razziale al razzismo 26 2.11 La Battaglia Contro Il Razzismo 27 2.12 Le razze fossili dell'uomo 28 Bibliografia 34 2
Introduzione
Non voglio qui perciò neppure accennare alla questione della «superiorità» o meno di un gruppo o
dell'altro, dato che dal punto di vista biologico il criterio di superiorità è costituito dalla pura e
semplice sopravvivenza ed espansione, comunque ottenuta, del patrimonio genico del gruppo 1.
Il paradosso che ogni studioso del problema razziale deve in primo luogo affrontare consiste nel
fatto che mentre da un lato risulta evidente che le diverse popolazioni umane possono differire
nell'aspetto fisico e comportamentale in modo anche radicale, dall'altro la linea di demarcazione fra
tipo e tipo è spesso graduale ed indistinta, con la persistenza di un'ampia base genetica comune a
tutte le divisioni infraspecifiche dell'uomo.
Inoltre le differenze fra gli individui di uno stesso gruppo sono spesso così rilevanti da rendere
difficile una generalizzazione, se non per media statistica, cosa che possiede sempre una validità per
il gruppo globalmente inteso e molto meno per l'individuo.
In effetti, nessuna altra specie, al di fuori del cane e di altri pochi animali che l'uomo stesso ha
addomesticato, presenta variazioni così macroscopiche di conformazione, statura, colore,
atteggiamento psichico.
Così ogni persona di senno riconosce oggi che le differenze fisiche immediatamente percepibili
nelle varie popolazioni umane possono raggiungere gradi anche estremi, e allo stesso modo ogni
studioso è al corrente dell'esistenza di differenze fisiologiche anche notevoli 2. Al contempo viene
però dai più negato che possano anche lontanamente esistere differenze di gruppo di origine
intellettuale, culturale, morale, di carattere e di inclinazioni, di valori insomma, che siano
geneticamente determinate, come geneticamente determinate sono le più immediatamente valutabili
differenze di ordine fisico-strutturale. Altri, al contrario, agganciano i differenti comportamenti
umani, con le loro motivazioni più o meno complesse, al carro del puro e semplice determinismo
biologico, misconoscendo o volutamente sottacendo le influenze ambientali, in specie storiche e
culturali, che pure sono parte non piccola dell'agire dell'uomo, unico essere vivente che non subisce
soltanto, ma trasforma l'ambiente e la propria condizione storica mediante autocoscienza e volontà.
Fra tali concezioni di quello che si può per semplicità e un poco impropriamente chiamare
«problema razziale», si dà quasi sempre un contrasto insanabile, con attuale prevalenza dei
sostenitori delle influenze ambientalistiche, i quali d'altra parte, in palese contraddizione con la loro
ideologia democraticamente «aperta», non esitano ad intralciare con ogni mezzo, dalla cortina di
silenzio presto calata intorno alle interpretazioni non conformistiche e dal pratico linciaggio
intellettuale fino all'aggressione fisica, la speculazione non diciamo di opposti irriducibili avversari,
ma di chi, preso da ragionevoli dubbi sulla bontà delle suddette tesi, si va accorgendo che alla base
di determinati comportamenti di gruppo non esiste soltanto e soprattutto il divenire storico e sociale,
ma la biologia3. Quanto ad identificare con esattezza le dimensioni qualitative dei gruppi umani
(sottospecie, razze, etnie, finanche impropriamente nazioni storiche e gruppi religiosi) coinvolti
1
La questione dei valori incarnati dai diversi raggruppamenti umani aveva ricevuto un'iniziale, suggestiva trattazione in
Gozzoli S., 1981 e anche in Vaj S., Biopolitica. Il nuovo paradigma, 2005, e più particolarmente nel capitolo "Specie e
razze" e successivi.
2
Ci si riferisce per il primo aspetto a colore della pelle, prognatismo ed altri indici cranici, forma del mento e di altre
particolarità della mandibola, forma e struttura dei capelli e dei denti, pelosità corporea, colore degli occhi, forma del
naso, ecc.; e per il secondo a gruppi sanguigni, metabolismo corporeo, preadattamento al freddo e al caldo, olfatto e
rappresentazione dei colori, dimensioni e peso degli organi interni, incidenza delle diverse malattie infettive,
neoplastiche ed ereditarie, ecc.
3
Come per la sua opera principale, Coon C.S., L'origine delle razze, Bompiani, 1970, anche per Ardrey R., L'istinto di
uccidere, Feltrinelli, 1968, fondamentale e suggestivo sguardo d'insieme sull'eredità animale del comportamento umano
(e indagine molto più profonda di quanto il rozzo titolo ad effetto dell'edizione italiana possa far supporre), c'è stata la
pratica messa al bando dalla comunità scientifica internazionale. In Italia le due opere citate risultano introvabili fin da
una decina di anni e non sono state più ristampate. Distrazione, insensibilità ai profitti editoriali, o non piuttosto
epurazione di fatto perché in troppi delle culture egemoni non vogliono neppure confrontarsi con tesi che risultano
eversive per le concezioni scientifiche e sociopolitiche dominanti?
3
nella questione «razziale» o a percentualizzare la misura di quanto sia dovuto all'ambiente e quanto
alla costituzione genetica di gruppo, noi riteniamo che questo sia cosa di difficile e per ora non
possibile attuazione, anche se un avvio mediante speculazione scientifica sta verificandosi in questi
anni con le nuove acquisizioni della sociobiologia.
Noi riteniamo inoltre che tale impresa potrà essere in futuro sempre più difficile, visto l'attuale
diffondersi per l'intero globo dell'ideologia occidentale, indifferenziata e livellatrice, dotata di
pseudo valori che lentamente e subdolamente vanno sgretolando le specifiche culture dei diversi
raggruppamenti umani,4 e che fornisce oltretutto ad entità statali da non molto resesi indipendenti
fra infiniti osanna progressisti, micidiali strumenti bellici con cui intraprendere stermini di massa e
genocidi,5 e visto pure il progressivo ampliamento del meticciato dei patrimoni genici con l'ipotesi
di una graduale, ancorché lontana, sparizione delle peculiari caratteristiche fisiche e mentali delle
diverse etnie, razze e perfino sottospecie, con l’eccezione di insignificanti gruppi periferici destinati
a configurarsi in un patetico folklore da riserva per le masse «moderne» in cerca di giustificativi per
le loro coscienze. In realtà tutti i tentativi, peraltro non numerosi, finora scientificamente compiuti
per separare i caratteri genetici dai caratteri non genetici, hanno avuto scarso e controverso
successo: non esiste finora una sola caratteristica mentale e psichica per la quale sia stata
quantitativamente stabilita con metodi scientifici una significativa differenza «razziale», nonostante
sia indubbio che tali differenze esistano. (Von Bertalanffy L., 1983, Dobzhansky Th., 1979).
Almeno fino ad oggi risulta, infatti, difficile e sempre contestabile misurare l'intelligenza innata di
un essere umano, senza misurare al contempo in qualche modo le opportunità che ha avuto e lo
sfondo culturale che gli è toccato, mentre risulta invece del tutto ovvia l'attribuzione ai diversi tipi
umani di un tipo di intelligenza (verbale, matematica, spaziale, ecc.) piuttosto che di un altro.
(Eysenck H.J., Kamin L., 1981) E d'altra parte, ai fini speculativi che ci poniamo, non ci sentiamo
certo di privilegiare l'aspetto «intelligenza», in parte numericamente quantificabile, rispetto ad altre
caratteristiche più sfuggenti, ma forse più essenziali per la sopravvivenza e l'espansione, fisica e
spirituale, dell'animale uomo, quali ad esempio la volontà, il coraggio, la creatività, l'inventività,
l'immaginazione, la coerenza responsabile, il senso morale, la solidarietà di gruppo, aspetti tutti
comunque meglio indagabili mediante lo studio della storia, naturale ed umana, piuttosto che con
l'applicazione di esatte metodiche scientifiche, sempre settoriali e che mai riusciranno a conferire ad
un gruppo la sua fisionomia sostanziale. (Larmat J., 1976) Per questa nostra posizione ci sentiamo
dunque in dovere di rigettare in toto il fanatismo ideologico totalizzante delle posizioni
ambientalistiche, che negano pretestuosamente e ferocemente il diritto di speculazione e di parola
alle opposte tesi, e che con il continuo celare e minimizzare la base biologica per assolutizzare
l'influsso ambientale e culturale sulle azioni umane (come se la cultura non fosse anch'essa
soprattutto un portato genetico), hanno da sempre provocato, a causa della menzogna loro
intrinseca, danni infinitamente più grandi e duraturi delle opposte tesi genetiche.
I geni, come è stato autorevolmente fatto notare da E.O. Wilson, tengono la cultura al guinzaglio,
permettendole sì escursioni in ogni direzione, ma in un ambito pur sempre limitato. Quando
sappiamo che il guinzaglio esiste, è possibile prendere decisioni più sagge riguardo agli elementi
4
Simile atteggiamento non è stato possibile adottare nei confronti di autori come K. Lorenz e e.O. Wilson, in quanto
appoggiati da numerosi discepoli e dalle due robuste scuole di etologia e sociobiologia, per cui non potendo
sommergere nell'oblio i numerosissimi testi a favore si preferisce ancor oggi o intimidire e demonizzare gli studiosi con
accuse «infamanti» che esulano dal campo scientifico, o minimizzare con sufficienza le loro teorie ed i notevoli risultati
ottenuti. Faccio accenno appena all'ostracismo e alla contestazione attiva di cui è stato per anni oggetto il professor H.J.
Eysenck da parte dei docenti e degli studenti «democratici» britannici, ostilità culminata nel 1973 nella selvaggia
aggressione all'interno della London School of Economics, con successivo ricovero ospedaliero dello studioso per le
lesioni subite. Con il portato di un irreversibile disfacimento culturale e fisico, come ad esempio per gli Eschimesi in
Canada e USA, gli Ik dell'Uganda, gli aborigeni australiani doverosamente recuperati alla civilizzazione ed al
progresso, oltre che al whisky, al «comfort» ed al tedio occidentali. Per gli Ik cfr., Gozzoli S., op. cit., pp. 222 e segg.
5
Come è avvenuto ad esempio per i Misquitos del Nicaragua, i Meo del Vietnam e gli Khmer della Cambogia, gli Ibo
della Nigeria ex-Biafra, i Watussi del Burundi, gli Eritrei dell'Etiopia.
4
della natura umana degni di essere coltivati, a quelli di cui si può gioire apertamente senza
artificiosi sensi di colpa, a quelli da maneggiare con precauzione.
Con similare linguaggio, espresso in altra forma, continua Ardrey: «Se usiamo la nostra intelligenza
in tutte le sue migliori possibilità, ci accorgiamo che l'animo non ha alcuna sovranità. Alleato
dell'istinto, il giudizio può operare. In conflitto con l'istinto il pensiero umano si degrada a
velleità».
Penso sia frutto solo di stolta presunzione, quando non di particolare interesse personale o di
conformismo intellettuale, il voler relegare in secondo piano l'aspetto genetico, soltanto perché
difficilmente quantizzabile con metodo scientifico; e che sarebbe veramente strano che l'animale
uomo, pur dotato di autocoscienza e volontà, fosse un qualcosa di unico e svincolato dal mondo
organico, nel senso che soltanto le caratteristiche morfologiche sarebbero controllate dai geni,
mentre tutti gli altri aspetti della psiche e del carattere sarebbero dovuti al condizionamento
ambientale o ad altri fattori non genetici.
Riteniamo infine che l'uomo attuale, prima di ogni eventuale discorso ideologico, religioso o
sociopolitico, intuitivo o razionale che sia, debba ormai tenere per indispensabile, quale base per un
ragionamento fondato e non mitico, l'acquisizione di dati scientifici ragionevolmente sicuri, e ciò
tanto più alla luce di conoscenze e riscontri affermatisi negli ultimi anni e provenienti in gran parte
dalla documentazione fossile.
Il discorso non dovrà poi vertere tanto sulle evidenti differenze attuali o su indagini e test di
laboratorio, quanto considerare piuttosto l'evoluzione storico-biologica dei diversi raggruppamenti
umani, avendo sempre presente, e questo non sarà mai sottolineato a sufficienza, che l'evoluzione
espressasi sulla Terra fino all'uomo attuale non è cronaca né dato di laboratorio, bensì storia, vale a
dire fatto dato per sempre; e che l'intima comprensione della dimensione temporale della questione
è cardine fondamentale per l'impostazione dell'intero problema.
Riprendendo l'insegnamento vichiano, si ricorda brevemente che «sapere è possedere l'origine di
una cosa, cioè il modo e la forma con cui è fatta», per cui criterio del vero risulta essere soltanto la
sua effettuazione, e la conoscenza vera e profonda della natura e della sua storia è possibile soltanto
a chi l'abbia fatta; l'uomo, nella sua esperienza, non può averne che una conoscenza «da imagine in
superficie»6.
Ogni invenzione concretizza sempre, infatti, un'idea, o progetto, o piano: quello che accade
nell'invenzione biologica, l'uomo lo ignora quasi totalmente. La nozione di tale progetto non può
essere sottoposta all’esperienza umana.
Sapienza vera risulta perciò alla fine essere per l'uomo non la scienza, ma lo studio della storia,
sereno e freddo, e studio, più che della storia naturale (di cui mancheranno in eterno le coordinate
precise, e di cui l'uomo non conoscerà mai la genesi profonda), delle «res gestae», compiute da lui
stesso, in quanto attività sua propria e dei suoi padri lontani. Ogni discorso biologico, peraltro
supporto indispensabile per l'uomo di oggi, risulterà perciò necessariamente per sempre
frammentario. Nonostante tutto questo, e anzi proprio per questa consapevolezza, dobbiamo poi
riconoscere l'insufficienza delle nostre categorie mentali, e che la storia umana non è
principalmente prodotto di una coscienza svincolata e non chiusa in un pensiero autonomo, ma che
sotto di essa, come i nove decimi dell'iceberg, persiste e pulsa la realtà biologica espressa nel DNA
e storicamente determinata dalle influenze in numeri del cosmo in esso per sempre fissate.
Occorre in secondo luogo prendere coscienza che, a prescindere dalla carenza e dalla talora voluta
omissione di dati biologici e paleobiologici, la scarsa comprensione della vastità del tempo
geologico ha finora spesso inficiato la ricerca speculativa, e che spesso per la decifrazione di eventi
lontani è stato fatto unico riferimento al tempo attuale, con l'illusione che i diversi fenomeni
6
Vico G., De antiquissima italorum sapientia, I°, 1, opera in cui il filosofo italiano, presentando le proprie tesi come
restauratrici di verità possedute dagli antichi italici, rivendica i limiti intrinseci della conoscenza umana, di contro alla
pretesa razionalistica di una matematica e di una fisica a priori, e in polemica con l'atomismo riduzionistico e con il
tema d'avvio della metafisica cartesiana, il famoso «cogito ergo sum».
5
paleobiologici fossero studiabili e riproducibili «scientificamente» in laboratorio o nel mondo
naturale di oggi.
È d'obbligo perciò il riferimento a tempi tanto lunghi da divenire talora incongrui alla mente umana,
alle centinaia di migliaia, ai miliardi di anni che hanno dilatato enormemente lo spazio ed il tempo
nei quali l'evoluzione ha operato sulla Terra, in maniera discontinua, ma progressiva per quanto
concerne la complicazione e l'aumento dello psichismo (e cioè la ricezione e l'elaborazione della
informazione ambientale) degli organismi fino a giungere, casualmente, all'uomo attuale7.
7
Voglio solo dire che le variazioni sul progetto di massima fissato nel DNA per tutti gli esseri viventi, hanno visto
sulla Terra infinite variazioni storiche, in dipendenza degli eventi propri di ogni segmento di tempo e di ogni
determinato strutturarsi del nostro pianeta e della sua posizione nel sistema solare e nell'universo. Definire la storicità
dell'evoluzione organica significa quindi definirne l'irreversibilità e irripetibilità: se per assurdo la vita potesse
ricominciare su una identica Terra primeva, né l'uomo né gli altri eventi evolverebbero di nuovo nelle forme attuali,
poiché da troppi eventi fortuiti è dipeso l'avvento delle attuali forme di vita sulla Terra. Così siamo d'accordo con
Ardrey che «l'uomo non è unico, né centrale, né necessariamente destinato ad essere, ma è il prodotto di circostanze
particolari fino all'incredibile». In caso contrario è d'obbligo concludere che tutto ciò che è razionale è reale, e
viceversa, e che quello da noi conosciuto è il migliore, ed anzi l'unico, dei mondi possibili.
6
1.1 Specie
La specie è un’unità fondamentale del sistema di classificazione tassonomica degli organismi
viventi, comprendente individui aventi caratteristiche simili, capaci di incrociarsi tra loro e di
produrre prole a sua volta fertile. Le categorie superiori e inferiori alla specie sono, rispettivamente,
il genere e la sottospecie (detta anche varietà o cultivar, nel mondo vegetale, e razza, nel mondo
animale). Il biologo tedesco Ernst Mayr, uno dei sostenitori della teoria dell’evoluzione, ha definito
le specie biologiche come “gruppi di popolazioni naturali, in grado di incrociarsi tra loro,
potenzialmente o di fatto, ma isolati riproduttivamente da altri gruppi analoghi”. Le specie e le altre
categorie tassonomiche superiori costituiscono un sistema formale, che permette di rappresentare i
modelli gerarchici di organizzazione degli organismi viventi. I componenti di una stessa specie
possono mostrare diverse distribuzioni geografiche, per cui ogni specie può comprendere
sottospecie più o meno differenti, che, pur tuttavia, sarebbero in grado di incrociarsi tra loro (specie
politipica).
Relazioni tra le specie biologiche
Predazione
Uccisione di un organismo da parte
di un altro vivente che se ne ciba.
Sono predatori i carnivori, che si cibano di erbivori (come il leone che
preda le gazzelle o l'airone che pesca i pesci); ma anche gli erbivori,
che si nutrono di vegetali, o, ancora, le piante carnivore, che catturano
insetti.
Competizione
Lotta, diretta o indiretta, tra diversi Tutti gli animali sono in competizione, sia per procurarsi il cibo, sia
organismi appartenenti a una stessa per appropriarsi di un luogo di nidificazione o di un territorio, sia per
specie (c. intraspecifica) o a specie conquistare un partner; ciò vale anche per le piante che, in
diverse (c. interspecifica), per particolare, sono in competizione per l'utilizzo della luce.
l'utilizzazione di una risorsa.
Commensalismo Relazione di tipo asimmetrico tra I pesci-pilota seguono altri pesci (di varie specie) e si cibano dei loro
due organismi: uno trae vantaggio da avanzi; gli aironi guardabuoi si cibano di piccoli animali che mettono
essa, mentre l'altro non ottiene né in fuga le mandrie. Ma un commensalismo 'assoluto' è raro: per
beneficio né danno.
esempio, nel caso delle remore, che si attaccano al corpo di squali e
balene e si lasciano trasportare da questi, si può pensare che anche i
loro trasportatori traggano un certo vantaggio, dato che le remore li
ripuliscono dai parassiti.
Simbiosi
Relazione tra due organismi che Numerosi protozoi e batteri risultano di aiuto per altri organismi,
risulta vantaggiosa per entrambi; durante la digestione, e ottengono, in cambio, un rifugio sicuro: per
spesso, ciascuno dei due non riesce a esempio, i protozoi flagellati vivono nel tubo digerente delle termiti e
vivere senza l'altro.
permettono la digestione della cellulosa. Alcuni funghi si legano alle
radici degli alberi (formando strutture dette micorrize) e ne
potenziano le capacità di assorbimento, in cambio di una piccola
quantità di linfa. Le acacie nutrono con le loro secrezioni zuccherine
alcuni tipi di formiche che, in cambio, le proteggono dagli attacchi di
insetti erbivori.
Parassitismo
Relazione di tipo asimmetrico tra Il cuculo depone le sue uova nel nido di uccelli di altre specie; il vischio
due
organismi
che
risulta succhia la linfa degli alberi sui quali si impianta. Spesso, il parassita è
vantaggiosa per l'uno e dannosa per molto più piccolo del suo ospite e può collocarsi all'esterno del corpo
l'altro: il parassita si nutre a spese di questo (ectoparassita) o all'interno (endoparassita).
dell'altro vivente (che prende il nome
di 'ospite').
1.2 Identificazione di una specie
Non è sempre possibile provare l’appartenenza di un individuo a una determinata specie, né
distinguere fra una singola specie politipica e un gruppo di specie simili che occupano aree vicine.
Inoltre, i tassonomisti che lavorano nei musei in genere si trovano a classificare esemplari
imbalsamati, senza poterne osservare il comportamento riproduttivo.
Di conseguenza, nei casi ambigui la sistematica continua a basarsi, nella pratica, sulla cosiddetta
specie tipologica, un concetto che risale a Platone e Aristotele ed è stato applicato alla moderna
tassonomia a partire dal XVII secolo da John Ray. In base al concetto di specie tipologica, una
7
specie è identificata da una serie di caratteristiche morfologiche (a cui si aggiunge qualunque
informazione disponibile sulla distribuzione, sul comportamento, sull’ecologia, sulla struttura
molecolare dell’organismo ecc.) che caratterizza in modo uniforme un particolare gruppo di
individui. Gli individui che non rientrano nel raggruppamento così definito vengono ritenuti
appartenenti a un’altra specie.
1.3 Speciazione
La speciazione è un processo di formazione di una nuova specie, a partire da una specie
preesistente, che avviene nel corso dell’evoluzione delle forme viventi.
Figura 1. Variazione clinale del mantello della zebra di Burchell. La zebra di Burchell (Equus burchelli) è
distribuita nelle regioni orientali e meridionali dell'Africa. Se si confrontano le popolazioni che occupano zone
diverse dell'aerale, scendendo da nord verso sud si osserva una progressiva riduzione dello spessore delle strisce
nere e della porzione del mantello da esse ricoperta. Sulla base di questi studi, gli zoologi ipotizzano che il
quagga, un tempo ritenuto una specie a se stante, sia in realtà una sottospecie della zebra di Burchell, la più
spinta delle versioni meridionali più povere di strisce. Per confermare l'ipotesi, stanno cercando di ricreare,
attraverso selezioni genetiche, esemplari il più possibile simili all'antico quagga, oggi estinto. Il fenomeno per cui
popolazioni diverse di una stessa specie presentano caratteristiche fisiche differenti a seconda della zona
geografica prende il nome di variazione clinale.
1.4 Modalità di speciazione
Due o più specie sono dette allopatriche, se occupano differenti areali; simpatriche, se coesistono; e
parapatriche, se i loro areali sono a contatto in una zona di confine. Benché tra gli studiosi non vi
sia accordo su come una specie dia origine a un'altra, il modello più comunemente accettato è
quello che porta alla formazione di due specie distinte e allopatriche a partire da un’unica specie
politipica (cioè una specie in cui si possono distinguere razze diverse), in seguito all'isolamento di
una delle razze che la compongono. Si formano, così, due nuove specie, che, anche se
ricongiungessero i loro areali o una invadesse il territorio dell'altra, non sarebbero in grado di
incrociarsi tra loro o, comunque, di produrre prole fertile.
1.5 Speciazione allopatrica
Tra i fattori che possono concorrere alla formazione di nuove specie vi sono l'isolamento
determinato da barriere geologiche o geografiche, o da stili di vita differenziati dal punto di vista
comportamentale o temporale.
In seguito a tale isolamento, la razza isolata inizia a evolversi progressivamente secondo una sua
direzione, distinta dalla via evolutiva intrapresa dalle altre razze appartenenti a quella stessa specie,
8
fino a che si crea un isolamento riproduttivo. Ciò significa che le differenze, accumulatesi nel corso
dei processi evolutivi, tra gli individui della razza rimasta isolata e quelli delle altre razze, sono tali
da impedire un eventuale accoppiamento e la possibilità di produrre una progenie.
1.6 Isolamento riproduttivo
L'isolamento riproduttivo, necessario alla formazione e alla definizione di una nuova specie
biologica, può avvenire prima o dopo l'accoppiamento. I meccanismi di isolamento precopula sono,
ad esempio, i diversi tempi di fioritura o i differenti rituali di corteggiamento che precedono
l'accoppiamento, mentre un classico esempio di isolamento postcopula è la sterilità del mulo,
risultato dell'accoppiamento di un cavallo e di un'asina. Entrambi questi meccanismi impediscono,
comunque, il rimescolamento genetico e la produzione di prole fertile a partire da individui
appartenenti a due nuove specie.
1.7 Speciazione simpatrica
Nel modello allopatrico l'isolamento è l'evento iniziale: esso avviene in modo casuale ed è seguito
da fenomeni di selezione naturale, che conducono le nuove specie lungo cammini adattativi
divergenti.
La speciazione simpatrica, invece, è considerata il risultato della selezione di adattamenti
alternativi. G.L. Bush, studiando i moscerini della frutta del genere Rhagoletis, osservò che alcune
specie si differenziano le une dalle altre, in quanto sfruttano ciascuna i frutti di alberi diversi, ma
presenti negli stessi frutteti, come le prugne e le ciliegie. Probabilmente queste specie hanno avuto
origine da una singola specie, inizialmente in grado di colonizzare entrambi i tipi di frutto e in
seguito specializzatasi nello sfruttamento di una o dell'altra pianta, dando così origine a due nuove
specie senza la necessità di un isolamento geografico.
1.8 Speciazione parapatrica
Il modello parapatrico prevede, invece, che due razze occupino territori adiacenti, con in mezzo una
zona di incrocio. La discendenza degli incroci interrazziali è probabilmente meno adattata di
ciascuno dei tipi parentali e, quindi, viene favorito qualunque meccanismo ereditario che porti ad
accoppiamenti tra simili. In presenza di caratteri adatti e di meccanismi di selezione
sufficientemente attivi, la conseguenza potrebbe alla fine consistere nella separazione delle razze in
due nuove specie.
La speciazione e i meccanismi che sono alla base di questo processo biologico costituiscono il
principale campo di indagine della biologia evolutiva. Benché quasi tutte le ipotesi formulate a
questo riguardo siano oggetto di forti controversie, molti ritengono che il più comune meccanismo
di speciazione sia probabilmente quello allopatrico, sebbene anche i modelli simpatrici e
parapatrici, nonché il meccanismo di isolamento precopula, abbiano i loro fautori.
1.9 La specie ed i gruppi intraspecifici
Basilare per l'inizio del discorso è la puntualizzazione dei concetti di specie, di speciazione e di
razza. Accanto alla concezione tipologica e statica dei tassonomisti che studiano le specie
attualmente viventi, dobbiamo sempre avere presente, per i fini che ci proponiamo, il concetto di
specie inteso in senso operativo e dinamico, o crono specie, per cui, soprattutto alla luce di una
nuova teoria dell'evoluzione, ci sarà di molto facilitata la comprensione dei mutamenti di tali
raggruppamenti, invarianti solo se visti con ottica attualista. Definiamo come specie non solo
raggruppamenti di individui morfologicamente simili, bensì comunità riproduttive i cui membri si
9
riconoscono e si ricercano l'un l'altro come potenziali compagni sessuali, con nascita di prole fertile
dalla loro unione8.
La specie risulta perciò essere un'unità ecologica che interagisce come tale con le altre unità insieme
alle quali vive nel medesimo ambiente, ed un'unità genetica consistente in un patrimonio genico
intercomunicante, essendo l'individuo soltanto un temporaneo recipiente che trattiene per breve
tempo una piccola porzione del portato genico del gruppo9.
Per giudicare invece se due individui fossili appartengono o no alla stessa specie, è evidentemente
impossibile il ricorso al criterio riproduttivo, visto che i reperti fossili non sono più forme
dinamiche. Occorre in questo caso valutare attentamente tutte le differenze morfologiche
riscontrabili, con successivo studio induttivo, parallelo e prudente, dell'habitat in cui sono stati
reperiti i fossili, dalla datazione temporale, dei confronti con organismi similari attualmente viventi.
Le forme fossili che differiscono in aspetto circa quanto differiscono le razze di oggi, sono definite
razze; quelle che differiscono quanto le specie attuali, sono dette specie.
Gli zoologi ed i paleontologi, oltre a basare i loro giudizi su tutti i caratteri che sono in grado di
identificare e misurare (caratteri che considerati nell'insieme conferiscono all'individuo la sua natura
quanto più essenziale) devono quindi operare quasi sotto forma di artisti-scienziati, poiché, come
afferma Coon, «la determinazione delle specie non può essere fatta mettendo delle schede in una
macchina calcolatrice. In un certo senso è un'arte, praticata da uomini esperti che sanno, avanti
tutto, come le specie si sono formate».
Divisione tassonomica immediatamente inferiore alla specie è la sottospecie, popolazione regionale
di una specie politipica che si distingue dalle popolazioni sorelle per occupare un territorio
geografico distinto e pressoché isolato, e che pur essendo legata alle altre dal criterio della
riproduttività fertile, è tuttavia dotata di differenze morfologiche e fisiologiche considerevoli.
In persistenza di un isolamento territoriale completo, la sottospecie assume i caratteri di una specie
in potenza, può cioè col tempo dare origine a nuove specie, alla fine anche molto diverse da quella
originaria. Nell'ambito di tali popolazioni inoltre, a causa di meccanismi di varia natura, possono
instaurarsi singoli complessi di geni per cui determinati sottogruppi vengono a differenziarsi da altri
della medesima sottospecie.
A tali gruppi, e solo ad essi, si deve attribuire il termine di razza10, vocabolo che in passato è
impropriamente servito, e tuttora impropriamente serve, per designare le sottospecie umane, o
addirittura al contrario etnie, popolazioni nazionali e gruppi religiosi. Il Chiarelli (2010) e altri
eminenti studiosi ritengono questo termine non sia consono a definire gruppi di popolazioni umani,
ma che sia più appropriato il termine di tipi. Condivido la visione del Chiarelli per l’uso del termine
tipo sia più corretto anche se sia un termine più empirico. Il termine razza ha assunto, nelle vicende
storiche, una connotazione equivoca ed equivocabile. Sarebbe giusto relegarlo al mondo vegetale e
animale.
8
Una delle più esaurienti impostazioni della questione si può trovare in Mayr E., L'evoluzione delle specie animali,
Einaudi, 1970, massiccia opera di alto livello specialistico che riporta numerosissimi dati biologici. A titolo puramente
suggestivo notiamo che sono a tutt'oggi conosciute circa 1.700.000 specie viventi, animali e vegetali, delle quali quasi
un milione appartengono alla classe degli insetti. Si ritiene che il numero totale di specie attualmente viventi possa
assommare a quattro milioni. Per quanto concerne le varietà estinte, sono state catalogate e depositate nei musei di tutto
il mondo circa 250.000 specie fossili, animali e vegetali. Si calcola che tale numero rappresenti solo una piccola
frazione di quella che è stata nel passato la varietà della vita: sulla base dei tassi di variazione evolutiva è stata avanzata
l'ipotesi che il numero totale delle specie estinte sia addirittura cento volte più grande, cioè 25 milioni di specie estinte
in oltre tre miliardi di anni.
9
Questo tipo di prospettiva, del tutto inusuale e sconvolgente per le correnti concezioni dell'individuo umano quale
essere autonomo e responsabile, è sviluppato in modo radicale in Dawkins R., Il gene egoista, Zanichelli, 1979.
10
Chiarelli B., L'origine dell'uomo, Laterza, 1981, pp. 49 sgg. Testo eminentemente didattico e riassuntivo, può essere
utile soprattutto per l'ampia trattazione dell'evoluzione dei Primati e delle differenze cromosomiche tra gli attuali
componenti di questo Ordine.
10
Definizione di tipo, si basa su particolari caratteri morfologici, anatomici, genetici, e psicologici,
presenti nella massima parte degli individui del gruppo.11.
1.10 L'evoluzione per equilibri intermittenti
Non è certo questa la sede per una disamina delle diverse teorie evoluzionistiche (selettive,
istruttive, neutraliste, organiciste, vitaliste)12 e neppure per una discussione sulla validità o meno
delle opposte concezioni antievoluzionistiche e fissiste13.
Mi limito perciò a ricordare soltanto che la teoria formulata per la prima volta nel 1859 da Darwin
tiene tuttora il campo, con gli aggiornamenti legati all'acquisizione di nuove conoscenze, fra la
maggior parte degli studiosi di cose biologiche e configura, a livello di opinione pubblica e presso i
non specialisti di media cultura, la teoria evolutiva tout court.
In realtà la dottrina darwiniana dell'evoluzione corrisponde oggi, come è stato autorevolmente
rilevato, a ciò che più si avvicina ad un laico «principio di fede». Mentre, infatti, l'evoluzione come
fatto non è praticamente più in discussione, tutto è ancora aperto per quanto riguarda
l'interpretazione del fatto, a dispetto di una certa ortodossia neo-darwiniana che tende a confondere i
due ordini di cose14.
Se cento anni fa era più che comprensibile la formulazione della teoria darwiniana in quei termini
ed in quei limiti che appaiono oggi sempre più evidenti, riteniamo che oggi si debba compiere un
ulteriore passo avanti, visto che l'azione della massima forza modulatrice – la selezione naturale
gradualistica - oltre ad ammettere un numero infinito di casi fortuiti, che non avrebbero mai potuto
verificarsi per mancanza di tempo e di un numero di generazioni sufficientemente elevato, è
pressoché totalmente negativa e comporta in primo luogo la morte15.
Pur in carenza di approfondite nozioni genetiche, di una scala temporale sufficientemente lunga, e
soprattutto di testimonianze fossili sufficientemente numerose, Darwin mise, infatti, l'accento, ai
fini dell'evoluzione delle specie, soprattutto sulla selezione naturale operante in maniera
estremamente graduale sull'intera specie nel corso di lunghissimi periodi di tempo. Ma mentre il
concetto di evoluzione organica veniva rapidamente e largamente accettato dal mondo scientifico
dell'epoca, la stessa cosa non poteva dirsi per l'importanza attribuita al meccanismo della selezione
naturale, ritenuto dai più insufficiente a produrre mutamenti di tale portata, e in ogni caso da porre
in netto subordine al mutamento interno degli organismi viventi.
Inoltre se la selezione fosse stata il primum movens, ciò voleva dire che gli organismi più bassi, da
cui si sarebbero originati tutti i successivi, dovevano contenere in sé da sempre “può essere
fastidioso per il ricercatore che preferirebbe poter ordinare in ben precisi reparti del suo
casellario, ma non le rende dei fenomeni biologici meno veri e reali” tutte le potenzialità evolutive
11
Come si può intuire da quanto finora esposto, ritengo del tutto ovvio, logico, scientifico e morale, non applicare anche
alla specie umana la nozione di razza, concetto che molti studiosi moderni, e primo fra tutti l'antropologo Ashley
Montagu, non ritengono scientificamente fondato, bensì conformante soltanto uno spregevole mito storico e puro
strumento di azione politica. Senza approfondire qui tale tematica, che richiederebbe adeguata trattazione in un nuovo
saggio, mi limito a citare il raro equilibrio intellettuale di Dobzhansky , op. cit., pp. 236 e 258-292: “Le razze e le classi
non sono né dal punto di vista biologico né da quello sociologico unità distinte o chiaramente definite”.
12
Per l'esposizione storica del fatto e del concetto di evoluzione, e del loro impatto sul pensiero occidentale, cfr. Eiseley
L., Il secolo di Darwin, Feltrinelli, 1975 e G Reene J.C., La morte di Adamo, Feltrinelli, 1971.
13
Riassunte modernamente in Sermonti G., Fondi R., Dopo Darwin, critica all'evoluzionismo, Rusconi, 1980, e in
Sermonti G., Le forme della vita, Armando, 1981.
14
La polemica antidarwiniana, vivace e da condividere quasi totalmente, svolta in Grassè P.P., L'evoluzione del vivente,
Adelphi, 1979. Occorre inoltre tenere presenti le sottolineature di von Bertalanffy, op. cit., pp. 357 e segg.: «Il moderno
evoluzionista, guidato dalla teoria della mutazione casuale e dalla selezione, non riesce a vedere che un organismo è,
ovviamente, qualcosa in più di un ammasso di caratteristiche ereditarie, o geni, messe insieme per accidente», con il
rigetto di ogni ipotesi riduzionista.
15
O una semiletalità ed una subvitalità che potrebbero togliere «ogni significato a tutta la teoria biologica
dell'evoluzione» (interpretata coi canoni dei neo-darwinismo, aggiungiamo noi), come è costretto ad ammettere
Dobzhansky , op. cit., pp. 140-142.
11
successive (in termini moderni diremmo che il primo essere vivente doveva contenere in sé geni
bastanti a generare le flore e le faune passate, presenti e future).
La selezione darwiniana e neo-darwiniana può, infatti, svolgere soltanto compiti di conservazione, o
di estinzione, e non di innovazione. Ciò si vide a maggior ragione dopo la riscoperta degli studi
genetici di Mendel, compiuta separatamente nel 1900 da De Vries, von Tschermak e Correns.
La mutazione, come fu chiamata la causa intrinseca del cambiamento, entrava ora a pieno diritto,
come prima forza agente, nella teoria evoluzionistica dominante. La mutazione (i cui meccanismi
intimi iniziano ad essere conosciuti solo oggi con i più recenti studi del citoplasma, del corredo
cromosomico, dei geni «regolatori» e di altre possibilità della cellula vivente prima neppure
sospettate e che fanno vacillare ipotesi, teorie, certezze e dogmi espressi anche solo pochi anni or
sono16 , allargava, infatti, continuamente l'ambito della variabilità delle popolazioni, molto prima
che queste fossero messe alla prova dalla selezione naturale, alla quale veniva negata quella
fondamentale importanza plasmatrice riconosciutale da Darwin.
La mutazione, intesa allora come macromutazione o mutazione discontinua di grande effetto,
poteva poi anche spiegare in modo più soddisfacente i cambiamenti avvenuti nei viventi nel breve
lasso di tempo dell'esistenza della Terra (venti-quaranta milioni di anni al massimo, secondo i
calcoli di Lord Kelvin e di altri fisici).
Nel decennio 1930-40, con nuove più ampie conoscenze di genetica molecolare e di popolazione, e
con l'estensione della scala cronologica a quasi cinque miliardi di anni, ritornava però in auge l'idea
darwiniana secondo cui l'evoluzione procede lentamente con accumulo di micromutazioni e che la
selezione a cui vengono sottoposti i risultati delle mutazioni è un processo opportunistico costituito
da tante tappe infinitesimali, con graduale trasformazione di specie già ben consolidate, nell'arco di
miriadi di generazioni o, il che è lo stesso, di decine e centinaia di milioni di anni, a velocità
costante e pressoché in tutti i gruppi di viventi. La selezione naturale, che riprendeva il ruolo
decisivo conferitole da Darwin, veniva suggestivamente rappresentata come una macchina immensa
che, pur lentissima, possedeva un'enorme potenza, tale da alterare drasticamente tutte le stirpi che
sopravvivevano per lunghi periodi di tempo.
Tale teoria, definita Sintesi Moderna da Julian Huxley e Theodosius Dobzhansky, era dovuta in
realtà ad una netta vittoria della genetica gradualistica nei confronti di altre discipline, e trovò
quindi presto i suoi oppositori. Tali furono il fitogeografo J.C. Willis e il genetista «eretico»
Richard Goldschmidt, che sostenevano l'improvvisa comparsa delle specie in seguito a
macromutazione radicale.
Su posizioni di parziale contestazione, anche se all'interno della medesima visione selettiva
dell'evoluzione, si situava anche un piccolo gruppo di biologi che, pur legati alla Sintesi Moderna,
stavano rivisitando con attenzione la tesi non gradualistica secondo cui la maggior parte delle più
importanti trasformazioni evolutive si doveva verificare in piccole popolazioni isolate e non
nell'intera specie (E. Mayr, Verne Grant, G.G. Simpson). A sostegno di tali sparute resistenze (e tali
le chiamiamo considerato il pratico terrorismo culturale operato dagli illuminati sostenitori della
Sintesi Moderna ai danni delle opposte teorie e basato sulle consuete accuse di oscurantismo
filosofico-religioso e di ingenuità scientifica, e ci riferiamo qui in primo luogo ai francesi Jacob e
Monod) giunse negli ultimi decenni una documentazione fossile sempre più vasta, della cui enorme
consistenza la maggior parte dei non addetti ai lavori ha ancor oggi solo una pallida idea.
Tra le discipline scientifiche soltanto la paleontologia fornisce in realtà le prove dirette, con vari
gradi di compiutezza, dei più importanti mutamenti sequenziali avvenuti nella flora e nella fauna
terrestri: il fatto evolutivo si manifesta al naturalista unicamente mediante la concretezza delle
forme fossili. Come dice Eiseley: «Se non fosse mai esistita la testimonianza delle rocce, se le
pietre fossero rimaste mute e le ossa morte non avessero mai parlato, l'uomo sarebbe ancora nel
dubbio».
16
Oltre che ai più recenti testi di biologia e citologia di livello universitario, rimandiamo a Grasse’, op. cit., pp. 267273, 300-307 e 316-331.
12
E inoltre altro e peculiare vantaggio offertoci dai fossili, è la cronologia dell'evoluzione, la
possibilità cioè di datare con precisione, attraverso di essa, il tempo geologico17.
Agli inizi degli anni Settanta vediamo così affermarsi, nettamente, di contro al gradualismo
darwiniano e proprio ad opera di due paleontologi, Niles Eldredge e Stephen Jay Gould, una nuova
teoria evoluzionistica, quella dell'evoluzione per equilibri intermittenti, o puntuati, secondo cui la
maggior parte dei cambiamenti è dovuta ad una speciazione «rapida» che interessa piccole
popolazioni (speciazione quantica) isolate dall'ambiente specifico e dotate di patrimoni genici
conchiusi in sé stessi, mentre le specie da cui tali popolazioni sono state isolate possono continuare
nei loro ecosistemi la loro vita per decine o centinaia di milioni di anni18.
A questo punto veniamo ad accorgerci che il vecchio assioma «natura non facit saltus» deve essere
completamente ribaltato: la speciazione, cioè l'acquisizione radicale di nuovi caratteri integrati, non
è più un fenomeno gradualistico che abbisogna di decine di milioni di anni per piccoli balzi, bensì
nuove specie (ovviamente ognuna con una sua propria potenzialità temporale) possono formarsi
anche nell'arco di migliaia di anni, un'inezia rispetto al tempo geologico ed al tempo cosmico, cioè
persino di poche centinaia di generazioni, con enormi balzi evolutivi e notevoli discontinuità
biologiche.
Diviene ora facile riconoscere come la ricerca dei famosi «anelli mancanti», o stati intermedi tra
diversi gruppi di organismi, sia risultata e debba in futuro risultare sempre vana, e non per l'inabilità
o la sfortuna dei ricercatori, ma perché essi, puro prodotto della mente umana, non sono mai esistiti
né possono esistere nella forma con cui sono stati concepiti dalla teoria gradualistica. Ed inoltre il
tempo geologico non fluisce per le specie con la medesima velocità e con lo stesso ritmo, bensì, per
cause che forse resteranno ignote per sempre, vede concentrate in «brevi» spazi temporali
esplosioni di vita imponenti, diversificazioni di molte nuove forme da antenati comuni («radiazioni
adattative»), rispetto ad altri momenti di relativa calma, quando non di stasi e di estinzione19.
Basti pensare alla Classe dei Mammiferi, i cui lontani progenitori comparvero, originati da un
gruppo particolare di «rettili», circa duecento milioni di anni or sono, con piccoli animali
morfologicamente simili ai toporagni insettivori attuali, e che rimasero pressoché stazionari per
oltre 135 milioni di anni, fino a quando si estinsero le fortune dei dinosauri. Quali che fossero le
cause di tale estinzione, vediamo che solo in quel periodo i piccoli mammiferi si accinsero a
raccoglierne l'eredità con una spettacolare diffusione su tutto il globo. La documentazione fossile
indica inoltre che le balene, con la loro mole, struttura, modi di vita, habitat acquatico, ed i
pipistrelli, con le loro ali ed il loro specifico ed anzi unico adattamento al volo, si sono allora
evoluti, in un arco di tempo massimo di dodici milioni di anni, da piccoli mammiferi simili a gatti; e
questo insieme a migliaia di altre specie e alla maggior parte degli Ordini di Mammiferi tuttora
viventi.
Un'altrettanto rapida «radiazione», e senza dubbio più spettacolare per il numero e la qualità
tassonomica dei gruppi coinvolti, è quella che ha riguardato gli organismi pluricellulari primitivi,
tutti viventi su fondali marini, che si è verificata all'inizio del Cambriano, circa settecento milioni di
anni fa, in un arco di tempo di poco meno di duecento milioni di anni (ma quasi totalmente da 570 a
520 milioni di anni fa) con l'origine di pressoché tutti i piani strutturali fondamentali degli esseri
17
La trattazione più recente della questione paleontologica si trova in Paul C., Storia naturale dei fossili, Etas libri,
1982. Con particolare riferimento ai mutamenti dell'ambiente terrestre, cfr. l'opera più scorrevole di Colbert E.H.,
Animali e continenti alla deriva, Mondadori, 1978.
18
L'esposizione più completa della teoria dell'evoluzione per equilibri intermittenti si ha in Stanley S.M., L'evoluzione
dell'evoluzione, Mondadori, 1982. Nata negli ambienti neo-darwiniani più avveduti (abbiamo già citato quali precursori
Mayr e Simpson, che cercarono di sganciare l'essenza del darwinismo, cioè la selezione naturale, da un insostenibile
gradualismo) tale aspetto della meccanica evolutiva è suscettibile di profondi sviluppi se inserito in più compiute teorie
evoluzionistiche, ad esempio quelle organiciste, che prospettano per il fatto evolutivo una differente, più articolata
dinamica.
19
Rileviamo da Ardrey, op. cit., pp. 244-246, di che genere potrebbero essere state alcune delle influenze cosmiche
sull'evoluzione del vivente. Per i critici eventi che posero fine al Cretaceo, cfr. Desmond A., L'enigma dei dinosauri,
Newton Compton, 1979, pp. 252-271 e Asimov I., Esplorando la terra e il cosmo, Mondadori, 1983, pp. 271-272.
13
animali, e cioè di pressoché tutti i trentacinque Phyla, o Tipi, evolutivi, di cui ventisei sono tuttora
viventi20.
Allo stesso ed opposto modo, un momento di cruciale rottura per estinzione può essere identificato
225 milioni di anni fa, alla fine del Permiano, estinzione della cui ampiezza può rendere
testimonianza la scomparsa del quaranta per cento delle famiglie di invertebrati dotate di scheletro e
viventi sulle piattaforme continentali. Fino a non molti anni fa, specie formatesi di recente erano
quasi sconosciute, ma ultimamente ne sono state identificate diverse.
Valga l'esempio delle falene hawaiane del genere Hedylepta, diversificatesi solo un migliaio di anni
fa dopo l'introduzione in quelle isole degli alberi di banano da parte dei polinesiani. Tale genere di
falene è andato incontro ad una speciazione multipla durante questo breve periodo, sviluppando
specie che si nutrono solo su banani e confinate in una o due isole soltanto.
Dell'iniziale formazione di nuovi generi di viventi testimoniano invece i pesci ciprinodonti della
Valle della Morte californiana, o i pesci ciclidi del lago vulcanico Barombi Bo del Camerun e del
lago Nabugabu, formatosi per distacco dal lago Vittoria, esseri tutti confinati in un complesso di
ambienti acquatici particolari costituitisi non più di qualche centinaio di migliaia, o addirittura di
qualche migliaio di anni fa.
A questo punto sono chiare le conclusioni di quanto finora esposto: nuove specie si possono
formare solo da piccoli gruppi isolati dalla popolazione principale, che evolve verso il suo destino,
sia esso l'estinzione sia esso l'arrivo fino ai tempi attuali, con minimi cambiamenti o sempre uguale
a sé stessa; è di fondamentale importanza l'assenza di esoincrocio e la costanza di inincrocio del
gruppo, cioè l'accoppiamento esclusivo all'interno della popolazione, senza apporto di patrimonio
genico esterno, col risultato del graduale rafforzamento del nuovo carattere, dovuto alla diffusione
tra i consanguinei; tali speciazioni, ognuna con una differente e sua propria velocità temporale,
possono avvenire anche nell'arco di qualche centinaio di generazioni, cioè soltanto di migliaia o
decine di migliaia di anni.
1.11 La speciazione quantica e l'uomo
In nessun settore della biologia, della zoologia e della paleontologia la concezione gradualistica è
difesa con maggior tenacia che nello studio dell'uomo. Le origini di tale concezione, che vede un
progresso monofiletico inarrestabile dagli esseri prescimmieschi alla spiritualità dell'uomo attuale,
può essere fatta risalire agli anni del primo darwinismo. La forte convinzione gradualistica che si
sviluppò allora, permise, infatti, all'uomo di riconciliarsi in qualche modo con la scomoda idea di
avere antenati animaleschi; era un modo per salvare la dignità umana: anche se l'uomo non era più
alla sommità di una Scala Naturae immutabile di origine divina, rappresentava pur sempre il
culmine del processo di selezione naturale.
Sostituita in tal modo la presenza attiva e rassicurante della divinità, il gradualismo ricreò, per un
intero secolo, una seconda gratificante illusione per l'essere umano, sbalzato di colpo dal trono di
una creazione immutabile all'accidentalità di esistenza in una natura estranea e cangiante e in un
cosmo lontanissimo ed indifferente, quando non ostile21.
Pur comprendendo appieno le ragioni di tale comportamento e riconoscendo che non poteva forse
allora avvenire diversamente, riteniamo tuttavia che l'uomo di oggi, inchiodato e quasi condannato
alle estreme conseguenze della sua razionalità, non possa più nascondere dietro paraventi
ideologici, filosofici o religiosi che siano, i risultati sempre più esatti e numerosi freddamente
offertigli dalle diverse discipline scientifiche (pur sospendendo necessariamente il giudizio nei
20
Grassè , op. cit., pp. 54-60 e Storia naturale ed evoluzione, editore Le Scienze s.p.a., 1979, pp. 113-125.
Vivida esposizione critica dell'Illusione della Posizione Centrale dell'uomo tacitamente accettata da Darwin (come
pure dell'Illusione della Bontà Originaria di stampo rousseauiano e marxista) leggiamo in Ardrey , op. cit., pp. 160-181.
Una piana ed esauriente sintesi delle conoscenze più attuali sulla nascita e l'evoluzione dell'universo si può invece
trovare, oltre che brevemente in Asimov, op. cit., pp. 264-284, in Reeves H., L'evoluzione cosmica, Feltrinelli, 1982.
21
14
confronti di quella Realtà Ultima che sta alle spalle dei legami fosforici e delle basi puriniche e
pirimidiniche del DNA).
Si può quindi capire perché, a differenza del gradualismo altrove applicato e che soltanto ora si
comincia a riconoscere non fondato, in campo umano tale deplorevole errore continui a persistere,
difeso accanitamente dalla maggior parte degli studiosi, con uno stupefacente salto di logica e
sacrificando la razionalità al sentimento.
Al contrario la teoria dell'evoluzione per equilibri intermittenti non solo fa piazza pulita di quello
che tutto sommato è il «finalismo spurio» darwiniano, secondo cui la selezione naturale con il
paziente lavorio dei millenni ha necessariamente condotto all'uomo attuale quale essere il più adatto
(con l'imposizione alla natura di una tautologica ferrea legge del massimo profitto),22 ma costituisce
una formidabile base di comprensione teorica per il principale studio sulle razze umane fossili,
quello portato a compimento da Carleton Coon nei primi anni Sessanta.
Per restare al primo punto del discorso, vediamo che il gradualismo darwiniano applicato
all'evoluzione umana consiste essenzialmente nel credere che ogni popolazione situata lungo quella
linea di continuità che porta dagli antropomorfi all'uomo, debba avere avuto un carattere intermedio
fra la popolazione precedente e quella seguente. Lo schema dell'evoluzione per equilibri
intermittenti complica invece le cose in quanto ammette deviazioni «illogiche» e sostanziali, dalle
quali, sempre parlando di Homo, avrebbero potuto avere origine esseri umani dotati di
caratteristiche fisiche e mentali del tutto diverse da quelle dell'uomo attuale. I mutamenti associati
alla speciazione riflettono sì le condizioni particolari di quella data situazione storica e geografica,
ma riflettono soprattutto l'accidentalità dei cambiamenti genici. Per soddisfare quale necessità, ad
esempio, gli uomini hanno acquistato gli uni capelli lisci, gli altri capelli crespi, altri ancora capelli
lanosi, nasi aquilini, camusi o schiacciati? È forse necessaria per la longevità dell'individuo, per la
sua adattabilità alle circostanze ambientali, per la sopravvivenza del gruppo, e infine per il
perpetuarsi del genere umano, il fatto che gli uomini abbiano o no gli occhi a mandorla o la plica
mongolica, le labbra tumide o il toro sopraorbitale?
A quale valore adattativo risponde la temperatura variabile dei bradipi in un ambiente in cui la
massima parte degli altri animali presenta costanza di temperatura corporea, o l'articolazione
semplice, e non doppia come quella dei rettili, della mandibola dei mammiferi con il cranio? O il
polmone destro di certe specie di rettili, allungato fino a raggiungere la cloaca, il quale possiede una
struttura alveolare respiratoria soltanto nel terzo anteriore, mentre per il resto è null'altro che un
sacco aereo senza funzione apparente?
Rimandando per un'ulteriore esemplificazione ad alcuni dei testi più recenti23 riportiamo a questo
punto un pensiero di Adolf Portmann: «La forma funzionale pura e semplice, che taluni tengono in
grandissima considerazione come la più conforme alla natura, è un caso raro e del tutto
particolare (...) Guardando l'involucro degli animali per lo più abbiamo l'impressione di trovarci di
fronte al prodotto di una fantasia senza scopo e, più che ad una necessità di ordine funzionale, ci
vien fatto di pensare all'opera festosa di un capriccioso, libero gioco delle forze creatrici».
Un capriccioso gratuito gioco dell'evoluzione (farei qualche riserva sull'uso dell'aggettivo festoso)
possiamo inoltre vedere nello studio della transizione ad Homo degli ominidi più recenti, con la
formazione di due linee evolutive, all'incirca tre milioni di anni fa, a partire da Australopithecus
afarensis, presente in un’area dell'Africa orientale che va dal lago Vittoria al Mar Rosso24. Le più
22
Sostiene il darwinismo in tutte le sue formulazioni che risultato ultimo dell'evoluzione mediante selezione naturale è
la sopravvivenza del più adatto. Ma alla domanda: chi è il più adatto?, abbiamo come risposta il ripiegarsi del
ragionamento su sé stesso: il più adatto è chi è sopravvissuto.
23
Facciamo riferimento al testo di un neo-darwinista: Attenborough D., La vita sulla Terra, Rizzoli, 1979, non perché
crediamo nella validità della spiegazione neo-darwiniana dell'evoluzione organica, ma perché riteniamo di scorgere,
negli innumeri esempi in esso riportati con l'ausilio di un'affascinante documentazione iconografica, sostegno alle tesi
espresse da Grasse’ , op. cit., pp. 203 e segg.
24
Quella che è stata definita «la scoperta del nostro più lontano antenato», e i problemi da essa posti alla ricostruzione
dell'albero genealogico del genere Homo, sono trattati in Johanson D.C., Edey M.A., Lucy, le origini dell'umanità,
15
recenti conclusioni spostano, infatti, Australopithecus africanus (il primo australopitecino scoperto)
dalla linea che porta ad Homo, per porlo all'inizio di una linea collaterale che si interrompe 1,5
milioni di anni fa con il più specializzato Australopithecus robustus. L'emergere della seconda linea
si verifica prima dell'estinzione di Australopithecus robustus, i cui ultimi esemplari vedono alla fine
come loro contemporanei i primi Homo erectus, e vede come punto di partenza i reperti fossili di tre
località dell'Africa orientale, i primi ai quali, sulla base dei riscontri morfologici e di una primitiva
industria litica, possa essere assegnata la qualifica di Homo (Homo abilis). L'impressione è che tutto
il movimento «ascendente» verso Homo sapiens sapiens, noi stessi, sia avvenuto soltanto per
tentativi ed errori; vi è sempre maggiore certezza che in qualche remoto periodo abbiano camminato
nella savana africana due o più specie umanoidi, entrambe ai loro tempi progenitrici potenziali
dell'uomo attuale e di ogni altro Homo sapiens mai nato. Con lo stesso procedere «gratuito» il
mento a punta dell'Homo sapiens, le arcate sopraccigliari sottili, la fronte alta e bombata,
compaiono dal nulla nella documentazione fossile: tali caratteri sono del tutto sconosciuti ed
imprevedibili rispetto a ciò che li ha preceduti.
E inoltre c'è un esempio particolarmente chiaro di incoerenza nella direzione dell'evoluzione
umana: le arcate sopraccigliari di Homo sapiens sono meno robuste di quelle di Homo erectus,
eppure quest'ultimo le aveva più robuste di quelle del suo predecessore abilis, e del suo remoto
antenato Australopithecus africanus.
Similare, e forse più significativo per la vastità dei reperti fossili, mutamento casuale di popolazione
si può riscontrare nella comparsa e nell'estinzione delle stirpi neandertaliane (specie a sé stante, o
più giustamente particolare gruppo caucasoide?). Tale vasto raggruppamento può, infatti, essere
visto come uno dei vicoli «ciechi» dell'evoluzione, estintosi forse a causa di competizione col più
evoluto cugino sapiens, dopo avere ceduto parte del suo patrimonio genico ai nostri diretti
progenitori caucasoidi. Riconsiderando dunque tutti i dati fin qui esposti, sentiamo che è tempo di
puntualizzare una prima conclusione: l'apparizione dell'uomo, quale attualmente è, costituisce il
risultato accidentale di un processo storico complicato ed enormemente lungo, in cui il puro caso ha
giocato un ruolo fondamentale nello stabilire le caratteristiche umane attuali.
E ciò vediamo con ancora maggiore profondità se risaliamo ancora più all'indietro nel tempo, più in
là dei più lontani antenati antropomorfi, fino alle origini di altre Classi e di altri Tipi di viventi, fino
agli immani sconvolgimenti geologici (mutamento del chimismo delle acque e dello stato
dell'atmosfera; radioattività endogena; deriva dei continenti con terremoti, vulcanismo e formazione
di nuove catene montuose e di mari; glaciazioni) ed astronomici (esplosione di supernovae;
radiazioni cosmiche; attività solare, precipitazione di asteroidi e meteore, il mutare posizione delle
galassie nelle loro rivoluzioni) con la formazione di eventi storici e genetici del tutto imprevedibili a
priori25.
I nostri denti ad esempio, iniziarono la loro storia come rivestimento squamoso del corpo di qualche
oscura creatura pisciforme nei mari di più di quattrocento milioni di anni fa. Se tali esseri non
avessero posseduto quelle particolari scaglie, certamente la storia intera dei Mammiferi sarebbe
stata diversa, poiché in questa storia i denti hanno avuto un’importanza primaria.
Inoltre le nostre braccia e gambe sono derivate dalle pinne pettorali e pelviche dei Ripidisti,
sottogruppo estinto dei Crossopterigi, progenitori di tutti i vertebrati terrestri (incidentalmente,
l'altro sottogruppo è tuttora vivente con minime differenze morfologiche rispetto ai lontanissimi
antenati). Se tali pesci avessero avuto un terzo paio di pinne, i vertebrati terrestri, uomo quindi
compreso, sarebbero potuto essere dotati di sei arti come gli insetti.
Se nelle pinne anteriori degli stessi pesci non si fossero trovate due ossa, ma solo una, l'evoluzione
umana non avrebbe potuto usufruire dei complessi movimenti di manipolazione permessi agli arti
Mondadori, 1981, specie alle pp. 271-285. Per quanto concerne la possibile meccanica cromosomica dell'ominazione,
oltre che a Chiarelli , op. cit., rimandiamo.a Ruffiè J., Dalla biologia alla cultura, Armando, 1978, pp. 214-220.
25
A meno di non ammettere che tutto fosse già minuziosamente scritto fin dall'inizio del Tempo, quindici miliardi di
anni fa, il che ovviamente ci consegnerebbe legati mani e piedi, per il bene e per il male, a un Destino sovrano, e
renderebbe vana qualsiasi speculazione nostra e di ogni altro essere autocosciente mai esistito nell'universo.
16
superiori dal radio e dall'ulna. E tuttavia per quel particolare pesce le due ossa non rappresentavano
alcunché di fondamentale, essendo semplicemente parte di una pinna flessibile che poteva essere
utilizzata anche per i movimenti sulla terraferma.
17
2.1 Razza: Evoluzione del concetto di razza
All’interno delle specie animali o vegetali, insieme di individui aventi caratteristiche simili; in
antropologia, classificazione della specie umana in “tipi”.
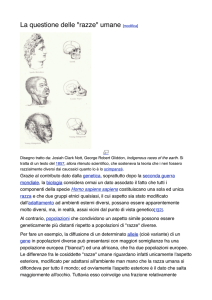
Figura 2. Principali razze umane Nel 1870 l'antropologo Blumenbach individuò cinque razze principali
(mongolica, caucasica, malese, americana ed etiopica), utilizzando parametri quali il colore della pelle, la forma e
il colore di occhi e capelli, la morfologia del cranio, la costituzione e la statura. Più tardi, nel 1889, De
Quadrefages individuò tre razze pure, la gialla, la bianca e la nera, e due razze miste, l'americana e l’oceanica,
basandosi sul cranio e sulla pelle. Un'altra classificazione è quella che si fonda su un criterio geografico, e
identifica gruppi razziali in base alle caratteristiche climatiche; questa classificazione è qui rappresentata, in
modo semplificato. In realtà, l'indagine su caratteri come la distribuzione dei gruppi sanguigni o del fattore Rh,
la sensibilità a particolari malattie e così via permette l'identificazione di gruppi che spesso non corrispondono a
quelli basati sui caratteri antropometrici esterni; gli studi sul genoma, inoltre, hanno dimostrato l'estrema
somiglianza genetica degli individui di "razze" diverse. Pertanto, una definizione esaustiva del concetto di razza
risulta difficile e infondata ogni classificazione che identifichi i diversi popoli come biologicamente distinti.
Per molto tempo, il concetto si è basato sulla pretesa di distinguere gli individui a seconda delle loro
caratteristiche fisiche, che, secondo la “teoria climatica”, venivano in gran parte determinate
dall’ambiente naturale. Lo studio sulle “razze” si affermò all’epoca delle grandi esplorazioni
geografiche, che, fornendo un abbondante varietà di “tipi umani”, stimolarono i tentativi di
sistemazione delle diverse popolazioni in uno schema generale e “scientifico”.
Nel XVII e XVIII secolo, di pari passo con lo sviluppo degli studi antropologici, furono elaborate
classificazioni che avevano in comune un approccio biologico alla questione e si basavano su una
combinazione di fattori anatomici e fisiognomici (colore della pelle, tipo di capelli, forma del
cranio, del naso, statura ecc.). Ad esempio, alcune di queste classificazioni dividevano il genere
umano in tre principali razze: negroide, mongolide e caucasica, ognuna a sua volta suddivisa in
sottorazze.
2.2 Razza e razzismo
Queste classificazioni, in realtà, più che descrivere le razze, svelavano la concezione che le ispirava,
che pretendeva di far risalire alla “razza” le diversità culturali, storiche ed economiche delle diverse
popolazioni. Non a caso, verso la fine del XIX secolo, si affermò, con Joseph Arthur Gobineau, una
teoria della razza fondata esclusivamente sulla cultura e sul “grado di civiltà”, che divideva le razze
in evolute o superiori e primitive o inferiori.
Nel XX secolo, lo sviluppo della ricerca genetica dimostrò la fragilità dei precedenti approcci
“scientifici”; infatti, finché venivano prese in considerazione soltanto le caratteristiche “esterne” era
18
possibile stabilire delle distinzioni, che si rivelavano però marginali a un’analisi “interna”, da cui
risultavano solo minime variazioni genetiche tra una “razza” e l’altra e spesso in contraddizione con
le variazioni fisiche esterne; infatti, si osservò che le variazioni genetiche avvenivano soprattutto
all’interno delle singole popolazioni, determinando un rimescolamento dei geni e la comparsa di
caratteri nuovi. Tuttavia, se il concetto di razza, per come è stato volta per volta definito nel corso di
molti secoli, non ha alcun fondamento scientifico, secondo la genetica moderna esso può continuare
a essere utilizzato: non per descrivere una realtà immutabile, ma l’effetto di un processo di
selezione naturale in continua evoluzione, che determina la diversa distribuzione delle
caratteristiche genetiche nelle differenti popolazioni.
2.3 Razze vegetali e animali
Una particolare accezione del termine razza è quella che definisce le differenti varietà di piante
(cultivar) o di animali, create dall’uomo allo scopo di ottenere individui dotati di particolari
caratteristiche, come ad esempio la resistenza a certi parassiti o la capacità di produrre maggiori
quantità di latte o di carne. In questo caso, la razza è l’effetto di un processo di selezione artificiale,
operata dall’uomo.
2.4 I geni e la storia
Gli studi che Luigi Luca Cavalli-Sforza conduce da quasi mezzo secolo hanno come grandioso
obiettivo quello di tracciare una “cartografia mondiale” dei geni umani, che consenta di identificare
i rapporti evolutivi tra le diverse popolazioni. L’albero genealogico che questa ambiziosa analisi ha
portato a costruire è stato messo a confronto con dati ricavabili da altri ambiti disciplinari, come la
linguistica o l’archeologia. I risultati indicano sorprendenti convergenze di prospettive nel
descrivere l’affascinante storia dell’umanità fin dai tempi delle più remote migrazioni umane
dall’Africa a tutti gli altri continenti del pianeta. Uno dei punti chiave di questa impressionante
ricostruzione della storia del genere umano – attraverso il suo patrimonio genetico, le sue lingue e la
storia dei popoli e delle civiltà – è il principio secondo cui le tradizionali teorie della razza, così
come erano state impostate dagli antropologi sette e ottocenteschi, non hanno alcuna valida base
scientifica.
Ogni popolazione pensa di essere la migliore del mondo. Con poche eccezioni, ognuno ama il
microcosmo nel quale è stato allevato e non vorrebbe mai lasciarlo. Per i bianchi, la più grande
civiltà è quella europea; la più grande razza è la razza bianca (francese in Francia, inglese in
Inghilterra, ecc.). Ma cosa ne pensano i cinesi? E i giapponesi? E gli extracomunitari che vengono a
cercare lavoro in Europa, proprio come un tempo gli europei in America, o adesso i messicani negli
Stati Uniti: non rientrerebbero forse nei loro Paesi d’origine, se solo potessero viverci
decentemente? Siamo forse i migliori, ma la storia ci mostra che ogni primato dura poco.
È soprattutto durante i momenti di grande successo che l’orgoglio delle nazioni è più forte. Quando
ci si sente forti è più facile pensare: “Noi siamo i migliori” e, in effetti, c’è un fondo di verità. Il
successo ha, comunque, origini abbastanza particolari: uno, o qualche uomo impone regole di una
certa efficacia, appropriate alla situazione, o si rende responsabile di azioni politiche intelligenti.
Anche se, spesso, queste azioni sono molto crudeli, dato che l’ascesa al potere richiede in generale
l’esercizio di una cera violenza, non necessariamente fisica, esse possono avere conseguenze
positive durature, talvolta sufficienti a determinare un periodo benefico per tutta la nazione. Un
susseguirsi di circostanze favorevoli può anche aiutare a mantenere la situazione stabile, ma questo
successo non dura mai troppo a lungo. È difficile che gli uomini politici intelligenti siano
rimpiazzati da successori ugualmente abili. La speranza che l’eredità biologica potesse produrre
questi successori si è rivelata assai poco affidabile, ma l’amore paterno continua a rendere ciechi i
politici, che seguitano a credere nella trasmissione del potere di padre in figlio. Il successo talvolta
dura per qualche generazione dopo la prima, grazie a un sistema politico ben congegnato che può
19
resistere all’imbecillità di qualche re e primo ministro succeduti ai fondatori di un impero o di un
nuovo regime. Durante queste generazioni fortunate, il popolo ha il tempo di convincersi che il
successo è dovuto alle proprie eccellenti qualità. Automaticamente, si decide che sono le qualità
intrinseche, innate, ereditabili dai nostri tipi umani che ci hanno fatti grandi; ma è quasi certamente
un’illusione. Ci si illude che questo processo possa durare fino a un futuro lontano, sfidando le
regolarità che la storia ci insegna. L’autocritica è rara e tende a mancare soprattutto quando le cose
vanno bene.
Come ha detto molto chiaramente, fra gli altri, Claude Lévy-Strauss, il razzismo è la persuasione
che una razza (la nostra, naturalmente) sia biologicamente la migliore – o che, come minimo, sia
eccellente. È alla superiorità dei nostri geni, dei nostri cromosomi, del nostro DNA che dobbiamo il
vantaggio su tutti gli altri. È più facile essere convinti della propria superiorità quando tutto va bene,
ed è molto facile dimenticare che il successo, al quale dobbiamo il nostro sentimento di superiorità,
non sembra durare a lungo, almeno se si giudica sulla base della storia passata. Ma non è veramente
necessario essere il numero uno per convincerci della nostra superiorità. Naturalmente, anche un
successo parziale aiuta molto, può costituire una prova importante agli occhi di tutti. Un solo
popolo è il numero uno in un certo momento, ma ce ne sono molti che lo sono stati un tempo, o
aspirano a diventarlo, e i numeri due, tre, quattro, ecc. pensano di avere lo stesso diritto di
inorgoglirsi. Ci sono dunque molti aspiranti a essere i migliori o, forse, molti candidati alla
superiorità – che si immagina, naturalmente, biologica e dunque eterna.
2.5 Altre origini del razzismo
In generale, qualsiasi popolo può trovare buone ragioni per considerarsi il solo eccellente al mondo,
o almeno uno dei migliori in qualche attività, sia questa la pittura, il calcio, gli scacchi o la cucina.
Il solo fatto di essere molto competenti in un determinato campo (artistico, intellettuale, sportivo,
amministrativo, politico, ecc.) basta spesso a dare a questa attività un’importanza maggiore di
quella che merita. La via è così aperta perché qualunque popolo possa considerare se stesso (o,
molto più di rado, un popolo diverso) superiore agli altri.
Figura 3. Manifestazione di afroamericani per i diritti civili. Negli Stati Uniti, durante gli anni Cinquanta e
Sessanta del XX secolo, tra la comunità afroamericana si affermò un forte movimento per i diritti civili e contro
la segregazione razziale ancora vigente in alcuni stati del Sud. Nonostante gli importanti risultati conseguiti in
quel periodo attraverso la lotta non violenta, negli Stati Uniti una consistente parte della popolazione
afroamericana e chicana (cioè quella di origine messicana) risente tuttora di una condizione di discriminazione
che ne pregiudica lo sviluppo sociale ed economico.
20
Ma svariati meccanismi ci dirigono verso conclusioni razziste, anche se non siamo necessariamente
al centro dell’attenzione o in un momento felice. Eccone uno che mi sembra importante. Ognuno di
noi è legato a una serie di abitudini, che sono alla base della vita di tutti i giorni, e alle quali ci
sarebbe difficile rinunciare. L’osservazione più superficiale mostra che le abitudini, i costumi, sono
differenti nei diversi Paesi. Anche se non conosciamo la natura o la fonte di queste differenze, il
semplice fatto che esistano ci porta a temerle. Non ci piace cambiare, anche se non siamo soddisfatti
di quello che abbiamo (ben poche persone lo sono, o lo sono completamente). L’attaccamento a
queste abitudini e la paura di essere costretti a cambiarle possono bastare a suscitare in noi un
autocompiacimento che potremmo chiamare razzismo. Credo che questo sentimento sia assai
diffuso e che molti di noi abbiano la tendenza a considerarsi migliori degli altri: quasi perfetti, o
abbastanza perfetti da far apparire sciocca la ricerca di cambiamenti significativi. Ma l’idea che le
nostre abitudini siano importanti nella genesi del razzismo è soltanto un’ipotesi, e non conosco
tentativi di provarne la validità.
L’esistenza di differenze tra le nazioni, tra i popoli, è un fatto chiaro a chiunque abbia viaggiato
almeno un poco. L’uomo della strada non si chiede a che cosa siano dovute. Ma, quando si tratta di
differenze di linguaggio, di colore della pelle, di gusti (culinari in particolare), o del modo di
salutarsi quando ci si incontra, esse sono là, a convincerci che gli altri sono diversi da noi. La
conclusione è solitamente che le nostre abitudini, i nostri costumi, sono i migliori. Tanto peggio per
gli altri, che sono diversi da noi. Sono chiaramente loro i barbari (i “balbuzienti”), come pensavano
i Greci. Può naturalmente accadere che qualcuno, non soddisfatto della vita che conduce nel proprio
Paese, sia pronto a sopportare la mancanza di comodità, e anche di sicurezza, che deriva
dall’emigrazione in un’altra regione, Stato o continente; che accetti dunque di sfidare la necessità di
imparare molte cose nuove, di affrontare lo sforzo che l’andare a vivere in un Paese diverso dal
proprio comporta. Ma in genere si preferisce, se si può, restare nel bozzolo nel quale si è nati. C’è
diffidenza, anche paura degli stranieri. Non si vuole dover imparare molte cose nuove, o rinunciare
a qualcuna delle proprie abitudini.
Tutto ciò ci porta a creare, e a mantenere, una buona opinione di noi stessi e di quelli che ci
circondano, una preferenza che può generare un senso di superiorità della propria persona e
soprattutto del proprio gruppo nei confronti degli altri; del tutto ingiustificata, forse, ma ciò
nondimeno potente. Ma vi sono anche molti altri motivi per diventare razzisti. Uno di essi,
probabilmente molto importante, è il desiderio di scaricare su qualcuno il proprio malcontento. Non
è un segreto che l’alienazione sia causa di irritazioni molto profonde, nella società moderna, e di
vera rabbia. Ciò può derivare dall’essere costretti a fare un lavoro disumanizzante, dal terrore della
disoccupazione, dalla realtà della povertà e dell’ingiustizia, dal senso di impotenza che ne deriva,
dall’osservazione spesso invidiosa delle ricchezze incredibili di un numero ristretto di individui.
Quale che sia la fonte della rabbia, ne saranno quasi sempre vittime le persone di livello sociale
inferiore; sono le sole che non possono reagire efficacemente. Basta un senso di superiorità relativa
per generare disprezzo verso quelli che pensiamo essere più in basso di noi nella scala sociale, e il
disprezzo sarà spesso più forte da parte di quelli che si credono, a torto o a ragione, disprezzati dalle
classi superiori. Effettivamente, sembra che non ci siano poveri così poveri che non possano
ricavare qualche soddisfazione nel prendersi la rivincita su quelli ancora più poveri di loro.
2.6 Esiste una base scientifica del razzismo?
Non si può negare che il razzismo sia molto diffuso. Nelle situazioni di equilibrio tra le nazioni
(nell’assenza, cioè, di guerra) e all’interno di esse (quando il conflitto tra le classi sociali non è
acuto), lo notiamo di meno. Ma le ragioni dello sviluppo delle ostilità tra i Paesi, le regioni, i popoli,
le classi, le religioni, i gruppi politici, l’arrivo di genti differenti, soprattutto l’immigrazione di
massa dai Paesi poveri, svelano il razzismo che si manifesta allora in tutta la sua potenza.
Si può condannare il razzismo perché è cattivo, e, in effetti, è condannato da quasi tutte le religioni
moderne, e da numerosi sistemi etici. Ma possiamo escludere che vi sia una razza superiore, o
21
almeno che esistano gradazioni di superiorità tra le razze; possiamo escludere che lo si potrebbe
dimostrare scientificamente? Le ragioni psicologiche del razzismo discusse più sopra sono difficili
da provare; possono essere false, o poco importanti. Ma si può esaminare se le vere differenze
genetiche tra i gruppi umani (ossia ereditate biologicamente, almeno in parte), di cui si può
constatare l’esistenza, diano una superiorità a un gruppo o all’altro. Ora, è certo che esistono
differenze tra i vari gruppi umani per il colore della pelle, i capelli, la forma degli occhi, della
faccia, del corpo; alcune di esse sono visibili a occhio nudo, e non c’è dubbio che siano, almeno in
parte, ereditarie. Esiste quindi già un insieme di conoscenze non trascurabile, del quale vogliamo
esaminare l’estensione, i significati e la spiegazione storica. Vediamo quali sono queste differenze,
e se diano una giustificazione scientifica al razzismo.
2.7 La natura delle differenze
Si deve dire, per prima cosa, che non è facile distinguere tra eredità biologica ed eredità culturale.
Talvolta, bisogna riconoscerlo, è difficile giudicare la fonte di una differenza. È sempre possibile
che le sue cause siano di origine biologica (le chiameremo genetiche) o che siano dovute ad
apprendimento (le chiameremo culturali), o che ambedue le sorgenti diano un contributo. Ma, come
già detto, esistono differenze tra le popolazioni umane che sono senza dubbio genetiche, vale a dire
ereditate biologicamente. E saranno queste a dover essere utilizzate per distinguere e studiare le
razze, per la semplice ragione che sono molto stabili nel tempo, mentre per la maggior parte le
differenze che fanno parte del tirocinio sociale sono ben più facilmente soggette a cambiamenti, e
possono a volte sparire in un tempo assai breve. Se le differenze strettamente genetiche fossero
davvero importanti da un certo punto di vista che può essere considerato motivo di superiorità di un
popolo sull’altro, il razzismo potrebbe essere giustificato, almeno formalmente. Bisognerebbe però
che la definizione di razzismo fosse ben chiara, e limitata a differenze genetiche.
Alcuni vogliono estendere la nozione di razzismo a qualsiasi differenza tra i gruppi, anche la più
superficiale e benché sia palesemente di origine culturale o sociale. Il solo vantaggio di questa
definizione estesa è che per parecchi caratteri è difficile dire se abbiano una componente genetica o
no. Ma non mi sembra appropriato definire razzismo il disprezzo di qualcuno per una persona che
parla a voce troppo alta, o è incapace di pronunciare correttamente i nomi stranieri, o non sa usare le
posate, o fa rumori grossolani quando mangia. Questo genere d’intolleranza mi sembra molto più
facile da correggere che il vero razzismo. In effetti, si tratta di un tipo d’insofferenza ben controllato
dalle persone che hanno un minimo di sensibilità e le sue cause tendono a diminuire con il tempo e
con il diffondersi dell’educazione. Sarebbe profondamente ingiusto condannare persone che non
hanno potuto ricevere un’educazione adeguata, specie se si tratta di difetti facilmente correggibili.
Qualche volta la correzione può essere disagevole, e per questo è importante esercitare la tolleranza,
o far uso di avvertimenti cortesi, anche se fermi. Credo che abbia profondamente ragione Voltaire
quando dice che vi è un’unica situazione in cui l’intolleranza è accettabile: quando si ha a che fare
con persone intolleranti.
2.8 Mutazioni visibili e invisibili
Le differenze tra le razze che colpivano i nostri antenati, e che impressionano molta gente ancora
oggi, sono quelle del colore della pelle, degli occhi e dei capelli, della forma del corpo, della faccia
e di tutti i dettagli, che ci permettono spesso di fare una diagnosi dell’origine di una persona con
una sola occhiata. Tranne che nel caso di incroci, è abbastanza facile riconoscere un europeo, un
africano, un orientale. Molti di questi caratteri sono abbastanza omogenei in ogni continente e ci
danno perciò l’impressione che esistano razze “pure” e che le differenze tra queste razze siano
molto forti. Questa diversità è, almeno in parte, di origine genetica. Il colore della pelle e le
dimensioni del corpo sono le meno ereditarie, essendo influenzate dall’esposizione al sole e
dall’alimentazione; ma c’è sempre una componente di eredità biologica che può essere importante.
22
Differenze del genere ci influenzano molto perché le possiamo vedere con i nostri occhi, e sono
chiare e incontestabili. A che cosa sono dovute? Lo sappiamo: sono quasi tutte dovute alle
differenze climatiche che sono state incontrate dagli uomini durante l’espansione nel mondo intero,
a partire dalla regione d’origine, l’Africa. E fino al momento in cui l’influenza tecnologica sul clima
da parte dell’uomo è stata modesta, e limitata alla costruzione di case molto semplici o alla
produzione di vesti di pelli d’animale per proteggersi dal freddo, un adattamento biologico è stato
evidentemente necessario. Abbiamo quindi un insieme di fenomeni, di cui possiamo abbozzare la
storia come segue (le giustificazioni delle ipotesi che facciamo verranno dopo).
Figura 4. Manifestazione per i diritti civili, Memphis. Una manifestazione per i diritti civili, organizzata nel 1968
a Memphis, nello stato americano del Tennessee, tenuta sotto controllo dalla Guardia Nazionale per evitare il
verificarsi di disordini. Sebbene il Congresso degli Stati Uniti avesse approvato il Civil Rights Act nel 1964, i neri
americani incontravano grosse difficoltà ad affermare i propri diritti.
1) L’espansione dell’uomo moderno dall’Africa agli altri quattro continenti ha comportato un
adattamento alle condizioni ecologiche, soprattutto di clima, molto diverse da quelle del continente
d’origine (a eccezione dell’Australia e di altre regioni tropicali). L’adattamento è stato sia culturale
sia biologico. Nel tempo trascorso da allora (cinque o sei decine di migliaia di anni o non molto di
più), si è potuta sviluppare una vera e propria differenziazione genetica. Ne vediamo chiare tracce
nel colore della pelle, nei capelli, nella forma del naso, degli occhi, del corpo in generale. Gli
antropologi hanno dimostrato che le differenze morfologiche tra i gruppi etnici sono state prodotte
dalla selezione naturale dovuta al clima. Il colore nero della pelle protegge coloro che vivono vicino
all’equatore dalle infiammazioni cutanee dovute agli ultravioletti della radiazione solare (che
possono provocare anche tumori maligni, come gli epiteliomi). L’alimentazione quasi
esclusivamente a base di cereali non permetterebbe agli europei di evitare il rachitismo, dovuto alla
mancanza di vitamina D in questi cibi. Ma i bianchi possono produrne abbastanza, partendo dai
precursori contenuti nei cereali, giacché la loro pelle povera di pigmenti melanici permette agli
ultravioletti di penetrarla e trasformare questi precursori in vitamina D, negli strati sottocutanei. La
forma e la dimensione del corpo sono adattate sia alla temperatura sia all’umidità; nei climi caldi e
umidi, come nella foresta tropicale, conviene essere piccoli per aumentare la superficie rispetto al
volume. È alla superficie che avviene l’evaporazione del sudore, che permette al corpo di
raffreddarsi. Essere piccoli aiuta, in certi ambienti tropicali, ad avere meno bisogno di energia, e
dunque a produrre meno calore all’interno del corpo quando ci si muove. In questo modo si può
diminuire la possibilità di surriscaldamento, che è alla base del pericolo di un colpo di calore. Gli
abitanti della foresta tropicale, e non soltanto i pigmei, sono dunque piccoli. I capelli crespi
permettono al sudore di restare più a lungo e prolungare l’effetto raffreddante della traspirazione.
23
Al contrario, la faccia e il corpo mongolici sono costruiti in modo da proteggere contro il freddo,
molto intenso nella parte dell’Asia dove vivono questi popoli. Il corpo e soprattutto la testa tenendo
il più possibile alla rotondità, e il volume del corpo è maggiore. Tutto ciò diminuisce la superficie in
rapporto al volume corporeo, e riduce la perdita di calore verso l’esterno. Il naso è piccolo – minore
pericolo di congelamento – e così pure le narici, in modo che l’aria arrivi ai polmoni più lentamente
e abbia il tempo di essere umidificata e scaldata. Gli occhi sono protetti dal freddo grazie alle
palpebre, che sono vere e proprie borse di grasso (fornendo in tal modo un isolamento termico
eccellente) e lasciano un’apertura molto sottile, dalla quale gli orientali riescono a vedere, pur
restando protetti contro i venti freddissimi dell’inverno siberiano. Gli occhi orientali sono spesso
molto belli, e Charles Darwin ha suggerito che le differenze che vediamo tra le razze derivino dai
nostri gusti. Può darsi che la forma degli occhi degli orientali, invece di essere un adattamento al
clima, sia dovuta a selezione sessuale? Però gli occhi degli orientali non piacciono solo in Oriente,
ma anche altrove; dunque perché è così frequente solo nelle popolazioni di origine orientale? Certo
il freddo ha la sua importanza, ma bisogna dire, peraltro, che gli occhi tirati sono comuni anche tra i
boscimani, che vivono in clima caldo. Comunque, è possibile che questo carattere si sia originato
per adattamento al clima in Siberia e si sia diffuso per selezione sessuale dalla Siberia agli orientali
del sud-est dell’Asia, dove il clima non è freddo. Anche se l’adattamento climatico è stato
all’origine il fattore più importante, non dobbiamo dimenticare la selezione sessuale, che può ben
avere contribuito all’aspetto esterno. In ogni caso, non c’è dubbio che si tratti di adattamento
ereditato biologicamente. Purtroppo, la base genetica di questi adattamenti non è ben conosciuta, ed
è quasi sicuramente piuttosto complessa.
2) I caratteri responsabili dell’adattamento climatico sono di solito molto omogenei, ed è
ragionevole che vi sia poca variazione individuale entro uno stesso gruppo, sottoposto allo stesso
clima. Tipicamente, essi mostrano una differenza elevata tra gruppi esposti ad ambienti diversi. Non
ci stupisce che l’abitudine a un determinato clima interessi tutti coloro che vivono nella stessa
regione, e che gli abitanti di tutto il mondo si siano genericamente adattati ai climi differenti che si
trovano in luoghi diversi. Ci si attende perciò che un carattere di adattamento climatico sia uniforme
nella stessa regione, ma differente da quello che troviamo in regioni climatiche diverse, pur se può
esservi qualche somiglianza tra popolazioni lontane, ma con clima simile, oppure in cui si sia avuto
per caso lo stesso tipo di selezione sessuale.
Ci si può chiedere se le decine di migliaia d’anni trascorse dall’occupazione dei continenti a oggi
possano essere state sufficienti per un adattamento biologico di intensità pari a quella che
corrisponde alle differenze osservate tra i gruppi. Questo genere di adattamento deve essere dovuto
a una selezione naturale molto forte, ed è assai verosimile che il tempo sia stato sufficiente.
Possiamo notare, al proposito, che gli ebrei aschenaziti, che sono vissuti al massimo per duemila
anni nell’Europa orientale e centrale, hanno un colore di pelle più chiaro degli ebrei sefarditi, vissuti
per un periodo altrettanto lungo (forse anche duemilacinquecento anni) nella regione mediterranea.
È possibile che si tratti di un adattamento dovuto alla selezione naturale, che avrebbe dunque
richiesto un tempo assai breve; ma è anche possibile che l’effetto sia dovuto, almeno in parte, al
flusso genico (matrimoni o unioni illegittime) con le popolazioni vicine.
3) I caratteri d’adattamento climatico sono tipicamente caratteri della superficie corporea. Questo è
prevedibile, in quanto la superficie del corpo costituisce l’interfaccia tra l’interno e l’esterno del
corpo stesso, ed è quindi di grande importanza nella regolazione del passaggio del calore
dall’esterno all’interno o viceversa e, perciò, per l’adattamento climatico.
4) La superficie del corpo è, naturalmente, molto visibile, quasi per definizione; siamo dunque
inevitabilmente influenzati dai caratteri che la riguardano. È un campione ambiguo di caratteri, che
ci orienta verso due conclusioni, ambedue false: le razze sono pure, e le differenze fra razze sono
forti. È soprattutto l’estensione di queste conclusioni agli altri caratteri che non è del tutto valida. In
effetti, è difficile trovare un altro motivo che spieghi la persuasione dei filosofi del diciannovesimo
secolo, come Gobineau, che era abbagliato dall’importanza di mantenere pura la razza, e totalmente
convinto che il successo della razza bianca dovesse essere attribuito alla grande differenza dalle
24
altre per i caratteri che riteneva ne fossero responsabili. Effettivamente, se ci si limita ai caratteri
visibili, i soli conosciuti a quel tempo, non è assurdo credere che possano esistere razze
relativamente “pure”. Ma allora non si sapeva che, per ottenere una “purezza”, cioè un’omogeneità
genetica (che comunque non sarebbe mai completa, negli animali superiori), si dovrebbero
incrociare per molte generazioni (una ventina almeno) parenti molto stretti, come fratello e sorella,
o genitori e figli. Ciò avrebbe conseguenze assai negative sulla fecondità e la salute dei figli e, ne
siamo sicuri, non è mai accaduto nella storia dell’umanità, se non per brevi periodi e in condizioni
molto particolari, come in alcune dinastie egizie o persiane. Non si sapeva nemmeno che, se si
studiano altri tipi di variazioni invisibili, non c’è alcuna omogeneità.
Vi è, al contrario, una grande eterogeneità genetica fra individui, qualunque sia la popolazione di
origine. Vale una regola del tutto generale [...]: questa variazione invisibile è sempre grande in
qualunque gruppo, sia esso quello di un continente, una regione, una città o un villaggio, ed è più
grande di quella che si trova fra continenti, regioni, città o villaggi. La purezza della razza è quindi
inesistente, impossibile e totalmente indesiderabile. Inoltre [...] le diversità tra le razze, definite
sulla base del continente d’origine, mostrano differenze molto modeste se paragonate a quelle
proprie dei caratteri visibili cari agli antropologi di una volta.
2.9 Razzismo
Il razzismo è l’insieme di teorie e comportamenti basati su una supposta divisione dell'umanità in
razze “superiori” e razze “inferiori”. Secondo le teorie razziste il patrimonio biologico
determinerebbe, oltre ai comportamenti individuali, gli sviluppi (culturali, politici, economici ecc.)
dei gruppi e delle società. Stabilendo questa connessione fra tratti razziali ed evoluzione sociale, le
concezioni razzistiche ritengono superiori le razze in grado di costruire società più 'evolute'.
2.10 Le origini del razzismo
Un atteggiamento di tipo razzistico è costantemente presente nella storia dell'umanità, come
testimonia la pratica antica della schiavitù. Gli antichi greci, e in seguito i romani, chiamavano
“barbari” (stranieri) quelli che non parlavano la loro lingua, avevano costumi, religioni, istituzioni
diverse e vivevano al “limite” del loro mondo.
Tuttavia, il razzismo per come noi lo intendiamo si sviluppò a partire dal XVII secolo, in seguito
alle scoperte geografiche e al colonialismo. In questo periodo si affermò la convinzione che il
progresso – intellettuale, scientifico, economico, politico – fosse un'esclusiva prerogativa dei
bianchi e che gli altri popoli non potessero conseguire gli stessi risultati proprio a causa di una
differenza biologica. Se fino a quel punto l'interpretazione prevalente del determinarsi delle varie
razze era stata quella 'climatica' – secondo la quale a un'origine comune erano seguiti sviluppi
dovuti soprattutto alle condizioni ambientali – dal XVIII secolo si affermò la teoria 'poligenetica',
che fa risalire le popolazioni del mondo a progenitori diversi.
L'affermarsi di questa convinzione portò a ritenere inalterabili le differenze tra individui e popoli e a
stabilire un principio di gerarchia secondo il quale la razza bianca era una razza superiore,
predominante sulle altre; in questo modo veniva giustificato il dominio sugli altri popoli da parte
dei bianchi e l'attribuzione a questi di una missione di civilizzazione.
25
2.10.1 Dalla teoria razziale al razzismo
Figura 5. Tensioni razziali in Sudafrica. Un gruppo di sudafricani legge il resoconto di uno scontro tra la polizia e i minatori
neri avvenuto nel 1973; lo scontro fece undici vittime.
Nel XIX secolo si consumò il passaggio dalla teoria razziale al razzismo, soprattutto con l'opera di
Joseph Arthur Gobineau Saggio sull'ineguaglianza delle razze (1853-1855). Gobineau affermò che
la razza è alla base della civiltà e che quindi la degenerazione della razza comporta un decadimento
della civiltà. Egli sostenne che per arrestare il decadimento della razza “ariana”, iniziato agli inizi
dell'era cristiana, non si potesse che perseguire un disegno di discriminazione delle razze
“inferiori”.
La pubblicazione del libro di Charles Darwin L'origine della specie (1859) ispirò in seguito una
nuova forma di razzismo, il cosiddetto “razzismo scientifico”, basato sull'idea che il pregiudizio
razziale svolgesse addirittura una funzione evolutiva.
2.10.2 Dal razzismo al genocidio
Durante tutto il XIX secolo il razzismo ebbe un'ampia diffusione in Europa, alimentato anche
dall'insorgere del nazionalismo, e negli Stati Uniti, dove era alla base del sistema schiavistico. Ma
fu dopo la prima guerra mondiale, nel quadro di crisi economica e sociale ereditato dal conflitto,
che le teorie basate sulla discriminazione razziale presero corpo in un disegno politico; infatti, la
Germania nazionalsocialista, a partire proprio dalla diffusione del mito della superiorità della razza
ariana, riuscì a mobilitare grandi masse e a raccoglierle attorno al progetto che aspirava a imporre la
supremazia germanica nel mondo.
Il mito della razza e lo stigma nazista nei confronti degli ebrei, che furono considerati
Untermenschen (sottouomini), legittimò e rese possibile il genocidio di sei milioni di ebrei e di altri
cinque milioni di persone considerate marginali, inferiori o devianti (accanto agli ebrei, zingari,
comunisti, omosessuali, disabili); non è un caso che il progetto di sterminio perseguito lucidamente
dai nazisti sia stato chiamato 'soluzione finale'.
È bene ricordare che la Germania nazista non fu l'unico paese a essere segnato dal razzismo; in
Italia, nel 1938 vennero emanate le 'leggi per la difesa della razza', che determinarono la
discriminazione degli ebrei e ne favorirono successivamente la deportazione nei campi di sterminio.
26
2.11 La Battaglia Contro Il Razzismo
Nella battaglia contro il razzismo un ruolo fondamentale è stato attribuito all'Organizzazione delle
Nazioni Unite (ONU), fondata nel 1945 anche per 'salvaguardare le generazioni future dalla
sciagura della guerra e dal razzismo'.
Figura 6. Deportazione di ebrei nei campi di concentramento Con l'avanzare dell'occupazione nazista in Europa durante la
seconda guerra mondiale, gli ebrei furono deportati in massa nei campi di concentramento. Le vittime furono oltre 6 milioni.
Nel 1965 l'Assemblea generale delle Nazioni Unite votò una Convenzione internazionale che definì
discriminazione razziale “ogni differenza, esclusione e restrizione basata sulla razza, il colore della
pelle, la discendenza e le origini nazionali o etniche, che abbia lo scopo o l'effetto di annullare o
rendere impari il riconoscimento, il godimento o l'esercizio su uno stesso piano dei diritti umani e
delle libertà fondamentali nella sfera politica, economica, sociale, culturale o in ogni altra sfera
della vita pubblica”.
Negli anni Sessanta negli Stati Uniti si sviluppò un ampio movimento contrario alla discriminazione
della popolazione nera, alla quale, dopo un secolo dall'abolizione della schiavitù, non veniva ancora
riconosciuta un'effettiva parità con i bianchi. Il dibattito sul razzismo sorto in quegli anni permise
inoltre di svelare e denunciare le ingiustizie che i neri americani avevano a lungo sofferto a causa
dei bianchi.
Nonostante sia ormai chiaro quali possano essere le conseguenze della diffusione del pregiudizio
razzista, questo continua a esistere e a riesplodere ogni qualvolta ci sia una 'responsabilità' da
attribuire a qualcuno.
27
Immigrati a Roma. L’Italia, paese d’origine di numerosi emigrati che si sono trasferiti principalmente nel resto
d’Europa e in America, ha recentemente conosciuto il fenomeno dell’immigrazione da parte di popolazioni provenienti
da diversi paesi, in particolare l’Africa, l’Europa orientale e il Sud-Est asiatico. L’intensità del flusso migratorio (seppur
inferiore rispetto a quello di altre nazioni), le difficili condizioni economiche interne e la mancanza di adeguate
infrastrutture hanno determinato un difficoltoso inserimento degli immigrati nella società italiana, provocando
sentimenti di intolleranza e talvolta deplorevoli episodi di razzismo.
Nel mondo contemporaneo, travagliato da conflitti e problemi, purtroppo queste occasioni non
mancano e, infatti, stiamo assistendo, accanto al riemergere di un nazionalismo aggressivo, alla
ricomparsa del fenomeno del razzismo, sempre alla ricerca dei 'capri espiatori' ai quali attribuire
responsabilità: ieri della degenerazione della razza, oggi della disoccupazione, della violenza e degli
altri innumerevoli problemi che affliggono le società contemporanee.
2.12 Le razze fossili dell'uomo
Se, come abbiamo detto, vi sono contrasti nell'attribuzione di determinate caratteristiche fisiche e
psichiche alle «razze» umane attuali, non esiste peraltro a tutt'oggi neppure accordo sul numero e
l'entità delle divisioni sottospecifiche e delle razze. La maggior parte degli studiosi riconosce
comunque attualmente poco più di una trentina di tipi viventi raggruppate in cinque sottospecie:
caucasoide, mongoloide, australoide, capoide, congoide, con sparsi, poco numerosi gruppi minori di
incerta classificazione26.
Il problema di quando tali sottospecie si siano differenziate costituisce il problema paleobiologico,
affrontato da Coon nella sua poderosa opera sull'origine delle razze umane27. Ma cosa viene a dirci
lo studioso americano? Semplicemente, e in maniera rivoluzionaria e provocatoria per le teorie
correnti, espone la tesi che le poche decine di migliaia di anni trascorsi dalla comparsa dell'Homo
sapiens moderno non sono periodo sufficiente per la comparsa e la stabilizzazione delle sottospecie
umane attuali, e che il momento di tale differenziazione deve essere posto molto più indietro, ad
26
Sguardo d'insieme a questo proposito può offrire, pp. 118-167, Kroeber A.L., Antropologia, Feltrinelli, 1983, testo
che risulta tuttavia datato, visto che l'edizione originale risale al 1948 (ma ampliata e riveduta da una precedente del
1923).
27
Dopo la pubblicazione negli Stati Uniti, nel 1962, della sua opera principale, l'intera questione delle somiglianze e
differenze razziali è stata abbandonata, e la ricerca accademica è pressoché cessata. Quello «razziale» risulta ancora
oggi un argomento così delicato che il voler investigare questioni del genere risulta offensivo per la maggior parte della
«pubblica opinione» e degli studiosi, anche se tali problemi sono parte sostanziale del mistero fondamentale che è alla
base della comparsa dell'uomo. Lo stesso Coon, colpito dalle polemiche violentemente scoppiate intorno alle sue tesi, e
dopo avere rischiato di essere messo al bando morale dalla comunità «scientifica» internazionale, si piegò ad una tacita
abiura di quanto sostenuto fino ad allora e, con un'ulteriore forma di raffinata captatio benevolentiae, si presentò, due
anni dopo, quale firmatario di una dichiarazione dell'Unesco sulla questione della razza, che sminuiva di molto le
conclusioni rivoluzionarie (e destabilizzanti per il conformismo intellettuale imperante) della sua ricerca.
28
un'età finanche di mezzo milione di anni (e oltre, diciamo noi sulla base delle scoperte di questi
ultimi vent'anni), ed all'interno di una specie umana non sapiens, l'Homo erectus.
Simile era stata negli anni Trenta l'intuizione di Franz Weidenreich, studioso dei resti fossili del
Sinanthropus pekinensis, intuizione frettolosamente liquidata, in parte anche per le sue potenziali
implicazioni razziste, dalla maggior parte del mondo accademico dell'epoca. Era, infatti, opinione
corrente che le sottospecie umane viventi si fossero differenziate da un antenato comune solo dopo
avere raggiunto lo stadio di sapiens (il che conferiva loro un'identica antirazzistica dignità formale),
i cui primi resti fossili erano valutati essere quelli degli uomini del Paleolitico superiore europeo, o
uomini di Crô-Magnon, apparsi circa quarantamila anni fa ed autori, tra l'altro, delle pitture di
Lascaux e Altamira.
Secondo tale concezione, il Sinanthropus, non essendo dunque partecipe delle caratteristiche di
sapiens, non poteva appartenere alla linea filetica di una sottospecie moderna, la mongoloide nel
suo caso, bensì alla linea filetica del genere umano, in qualità di specie pre-sapiens non ancora
differenziata.
In realtà uno degli errori capitali di tale interpretazione consisteva, come rilevò Coon, nel fatto che
non si era tenuto nel minimo conto, nella messa a punto dello studio degli uomini fossili, che oltre
alla dimensione temporale c'era anche, altrettanto importante, quella spaziale; quasi tutti gli autori si
erano preoccupati unicamente della sistemazione cronologica dei reperti, ignorando pressoché
completamente la collocazione geografica delle stazioni fossili. Dal lungo, minuzioso studio
ventennale di tali reperti inseriti in una corretta dimensione spaziale, lo studioso americano giunge
quindi ad evidenziare che la sottospeciazione umana deve essere situata molto a ritroso nel tempo,
quando tipi oggi estinti di un'altra specie umana, l'Homo erectus, dotata di proprie caratteristiche
fisiche, culturali, e spirituali, erano ancora vivi e non erano apparsi i successivi sapiens.
La transizione a sapiens risulterebbe inoltre essersi attuata, per le diverse sottospecie, in gran parte
indipendentemente, in zone territoriali separate e nel corso di centinaia di migliaia di anni, con un
minimo flusso intraspecifico di geni, tale comunque da permettere, anche con l'ausilio della relativa
brevità del tempo trascorso, l'unità della specie, ma non da eliminare le sottospecie28. Si ricorda a
questo punto l'interessante ipotesi di Darlington, secondo la quale il colore originario dei primi
erectus non ancora migrati dalla loro culla est-africana e risalenti almeno ad un milione e mezzo di
anni fa, avrebbe dovuto essere bruno chiaro giallastro e che mutazioni sarebbero avvenute
indipendentemente verso il bianco nelle terre circum-mediterranee e vicino-orientali per dare i
caucasoidi, verso il bruno scuro/nero nell'Asia sudorientale per dare gli australoidi, verso il nero in
Africa per dare i congoidi, mentre le rimanenti due sottospecie avrebbero mantenuto con minime
variazioni il colore originario. Anche le evidenze offerte dall’etnologia comparata, dalla genetica
dei gruppi, dalla linguistica e dall'archeologia preistorica, parlano a favore di una separazione molto
antica delle sottospecie umane. Inoltre il presunto Homo sapiens preistorico, fonte secondo la teoria
corrente di tutti i tipi attuali, risulta morfologicamente simile all'europeo vivente (certo con le
minori caratterizzazioni razziali e sottospecifiche dovute all'antichità dei reperti) e sicuramente
nessuno è in grado di sostenere fondatamente la possibilità di far discendere, nell'arco di poche
centinaia di generazioni, un aborigeno australiano od un pigmeo del Congo da antenati europei di
tipo moderno.
Bernard Rensch afferma poi, a ulteriore convalida, che il particolare adattamento razziale dell'uomo
a differenti habitat, adattamento che riguarda un certo numero di geni, richiede necessariamente
decine di migliaia di generazioni, cioè diverse centinaia di migliaia di anni, per instaurarsi
completamente. Egli porta inoltre diagrammi statistici a sostegno della tesi per cui la mutazione di
un gene interessante l'ipofisi potrebbe aver dato l'avvio alla transizione da erectus a sapiens, pur non
riuscendo ad impedire la fertilità tra i possessori di questo carattere e coloro che ne erano privi.
Dato infine che simile mutazione si verificherebbe una volta su centomila persone nel corso di una
28
Per il problema dell'emergenza dei gruppi sottospecifici e del successivo passaggio a nuova specie col meccanismo
dell'evoluzione parallela, rimandiamo, oltre ai migliori testi universitari di genetica e di biologia, a Mayr , op. cit., e a
Darlington C.D., L'evoluzione dell'uomo e della società, Longanesi, 1973, pp. 21-24.
29
generazione, il calcolo delle probabilità non escluderebbe la possibilità che più di una sottospecie
abbia varcato indipendentemente la soglia fra erectus e sapiens, e questo anche senza considerare
l'«aiuto» genico offerto alle sottospecie più tardive dalle prime che compirono la transizione. Ma a
prescindere da tali speculazioni genetiche, restano inconfutabili le misurazioni ed i dati
paleontologici a convalidare la tesi di Coon, in precedenza avanzata a livello speculativo anche da
altri studiosi, quali Frank Livingstone e Loring Brace.
In realtà, ripeto, lo studio attento di Coon, preciso fino alla pedanteria, con la considerazione di ogni
dettaglio e con la misurazione minuziosa di ogni possibile dato offerto dai fossili umani provenienti
da quasi trecentocinquanta località del Vecchio Mondo, rende sempre più salda, fino quasi alla
certezza, la tesi che gli uomini differivano fra loro per peculiari caratteristiche «tipiche» (o meglio
sottospecifiche) già allo stadio di erectus. Gli alberi genealogici che vanno attualmente per la
maggiore continuano al contrario pervicacemente ad illustrare una linea filetica unica, che
dall'australopitecino vivente nel solo continente africano, sale all'Homo erectus vivente in ogni parte
del Vecchio Mondo e in ogni parte del Vecchio Mondo affermato con sue specifiche industrie
litiche, e quindi all'Homo sapiens presente inizialmente nelle sole terre circum-mediterranee, che
inizia solo a questo punto, a suddividersi in gruppi razziali che si sposteranno a grande velocità
(sono abituati a muoversi in fretta, visto che sono cacciatori ed hanno i garretti ben solidi!) per
occupare i siti lasciati opportunamente liberi dagli erectus svaniti ovunque nel nulla dopo un
milione di anni in cui non hanno minimamente pensato a generare gruppi sottospecifici.
Dobbiamo invece riconoscere che risulta molto più convincente sul piano della teoria e della logica
e molto meglio documentata29, l'opposta concezione di Coon; in ogni caso le linee sottospecifiche
risultano tracciate in modo così compiuto da escludere che ulteriori ritrovamenti possano, almeno
per le sottospecie eurasiatiche, determinare sostanziali modifiche.
Il materiale africano (linee capoide e congoide) è invece meno ampiamente documentato, ed è
possibile che nuovi reperti possano indurre a conclusioni leggermente diverse, soprattutto riguardo
alla propagazione spaziale delle due sottospecie.
In ogni caso non è per puro accidente che il complesso territoriale Asia occidentale-EuropaNordafrica ha costituito la zona più densa di gruppi razziali più diversificati, e quella che prima fra
tutte, circa 250.000 anni fa, ha visto la transizione dell'Homo erectus ad Homo sapiens arcaico.
L'alternarsi di climi freddi ed umidi durante le ripetute avanzate dei ghiacci, e di climi caldi ed aridi
durante i periodi interglaciali, determinarono in tale ambito territoriale, già di per sé frammentato da
catene montuose e da distese marine continuamente mutanti, l'avvicendarsi di numerosissime specie
animali. Le oscillazioni climatiche con la formazione di molteplici differenti habitat geografici
furono abbastanza grandi da stimolare fatti evolutivi diversi e numerosi.
Tale ambito territoriale rappresentò per oltre mezzo milione di anni, la sede potenzialmente, e di
fatto, più adatta per l'evoluzione dell'uomo. In tale sede e per tale periodo, anche se in tempi diversi,
la sottospecie caucasoide venne a contatto con almeno tre altre sottospecie (australoide, capoide,
congoide) considerato che l'Asia centrale rappresentò per lunghissimo tempo una barriera glaciale,
paludosa e desertica pressoché insormontabile per quei primi gruppi umani.
29
Sottolineiamo ancora la capitale contraddizione, anche puramente logica, di una teoria che afferma che, mentre la
specie erectus sarebbe sopravvissuta senza sottospeciazione per oltre un milione di anni in ambienti quanto mai variati,
la selezione naturale sarebbe piombata invece inesorabilmente sui sapiens, e in poche migliaia di anni avrebbe condotto
a gruppi umani diversissimi, presentanti differenziazioni anche estreme di ordine fisico, fisiologico e psichico. Altro
notevole esempio di superficialità e di inconsistenza logica e scientifica è la concezione di Ruffie’ J., op. cit., pp. 199 e
368, che identifica l'estremo Oriente o il sud-est asiatico quale luogo di provenienza dei sapiens crô-magnon. Inoltre lo
stesso autore, mentre da una parte ammette, seguendo Coon, un'origine plurifocale di sapiens da erectus, dall'altra nega
invece pervicacemente l'esistenza attuale di gruppi umani provvisti di una propria individualità biologica, considerando
il concetto di «razza» unicamente quale mito storico-politico prodotto dai diversi contesti socio-culturali, in ispecie dei
contesti socio-culturali europeo ed occidentale dei secoli XIX e XX. Rilevando come in tal modo l'autore perda di vista
la prospettiva biologica del fenomeno «cultura», il curatore dell'edizione italiana nota invece criticamente: «La
concezione puramente biologica dell'uomo si va riaffermando oggi come l'unico punto di vista valido perché tutti i dati
raccolti dalle varie discipline possano essere coordinati ed interpretati all'interno di una teoria che ne garantisca la
scientificità».
30
I mongoloidi furono al contrario in grado di scambiare i propri geni soltanto con un'altra
sottospecie, l'australoide, che dalle originarie zone indo-indocino-malesi sotto la pressione della
sottospecie più evoluta si ritirò lentamente nella zona papuo-australo-oceanica, lasciandosi dietro
razze relitto quali i Vedda dell'India e i Negriti delle Andamane, della Malesia e delle Filippine.
I congoidi furono sicuramente in contatto con l'altra sottospecie africana, e forse videro pure una
limitata periferica acquisizione, e cessione, di geni caucasoidi. Transitati a sapiens qualche decina di
migliaia di anni fa, la loro espansione settentrionale trovò il suo limite nella zona tropicale occupata
da gruppi razziali caucasoidi e successivamente nell'ormai inaridita regione sahariana. In tempi
ormai storici giunsero poi all'Oceano Indiano e, mentre i primi europei (portoghesi ed olandesi) si
addentravano nell'Africa australe, iniziarono la loro discesa verso i territori che stavano
spopolandosi dei gruppi capoidi, fino a scontrarsi nuovamente con gruppi caucasoidi30. I capoidi,
originatisi sull'intera fascia nordafricana e successivamente migrati nell'Africa australe scavalcando
la primitiva zona di diffusione congoide, furono sicuramente in contatto con le sottospecie
caucasoide e congoide.
La situazione geografica fornì dunque ai mongoloidi l'isolamento indispensabile per la
conservazione delle loro peculiarità razziali più tipiche, ed essi sono, infatti, a tutt'oggi considerati
la sottospecie più omogenea (nei suoi grandi gruppi asiatico ed amerindo) e più lontana dalle altre.
Allo stesso tempo la geografia permise ai caucasoidi di trovarsi in un'area zoologica centrale (e,
come abbiamo visto, più favorevole delle altre alla differenziazione, a causa dei più numerosi
gradienti climatici ed ambientali) nella quale oltre a ricevere da almeno altre tre sottospecie un
limitato apporto genico, poi sottoposto alla dura selezione dei climi e delle culture, furono in grado
di cedere agli altri gruppi parte, sempre minima, del loro patrimonio genico.
Al centro delle masse continentali del Vecchio Mondo, essi furono la meno uniforme tra tutte le
sottospecie umane, e la testimonianza di ciò si trova nei diversissimi reperti fossili di quest'area,
dalle svariate stirpi neandertaliane, occidentali, centroeuropee, orientali e del Vicino Oriente; agli
Homo sapiens arcaici che li precedettero nell'Ultimo Interglaciale, come la differenziata
popolazione di Krapina in Croazia; ai successivi popoli del Paleolitico Superiore cui è stato dato
nome di Crô-Magnon31.
Verso la fine del Paleolitico, dopo che tutte le cinque sottospecie erano divenute sapiens, e prima
che le due più settentrionali avessero portato a termine la loro espansione verso sud e verso est, per
mare e per terra, tutte comprendevano presumibilmente lo stesso numero di individui (si valuta il
loro ammontare complessivo ad una decina di milioni di unità). L'uso del termine espansione si
rivela in realtà più esatto di migrazione; gli spostamenti più importanti ebbero luogo in tempi
lunghissimi, con inizio forse centomila anni fa: non si trattò di improvvisi e volontari trapianti di
intere popolazioni, ma piuttosto di una costante deriva di piccoli gruppi di cacciatori che potevano
contare anche poche decine di individui, al seguito degli spostamenti della selvaggina o perché
sospinti da altri gruppi umani e dai mutamenti climatici.
Questo lungo periodo di movimento fu senza dubbio un periodo di grande ibridazione fra i gruppi;
ibridazione possibile certo a diversi livelli, ma che avvenne soprattutto fra gruppi tribali affini delle
diverse razze e sottospecie. Che le commistioni fra le principali suddivisioni dell'umanità siano state
del tutto sporadiche e senza esito rilevante di meticciato, lo possono testimoniare quei piccoli
gruppi di uomini isolati in popolazioni del tutto differenti che mediante un istintivo ostacolo
all'esogamia mantengono a tutt'oggi, anche dopo decine di migliaia di anni, la loro integrità
sottospecifica e razziale, quali i caucasoidi Ainu del Giappone, i superstiti avanzi mongoloidi del
Deccan, del Transvaal e della Tanzania, i gruppi negriti di ascendenza australoide sparsi dall'India
alle Filippine. In seguito all'introduzione, a partire dal Vicino e Medio Oriente, dell'agricoltura e
30
Il popolamento dell'Africa del Sud vide una precedenza di gruppi bianchi rispetto ai gruppi bantù che costituiscono
oggi la maggioranza delle popolazioni di quelle regioni, per cui è giocoforza riconoscere agli Afrikaaner attuali una più
«legittima» aborigenicità, rispetto ai discendenti dei congoidi (la qual cosa, in sé, non prova ovviamente nulla).
31
Certo tale maggiore variabilità delle popolazioni europee arcaiche potrebbe essere in parte addebitabile alla
documentazione fossile riscontrata in Europa più numerosa che in altri continenti.
31
dell'allevamento (oltre che sulla base di quei fattori morali di cui parla Gozzoli32, scientificamente
imponderabili eppure ben identificabili nel divenire storico dell'uomo), circa diecimila anni fa le
sottospecie caucasoide e mongoloide, delle quali come già accennato si può seguire a ritroso il
cammino verso le origini di sapiens per un tempo molto più remoto che per le altre, cominciarono a
superare numericamente in modo netto gli altri gruppi sottospecifici.
Duemila anni or sono il numero complessivo di individui umani era salito a trecento milioni; supera
oggi i quattro miliardi (per oltre il novanta per cento appartenenti ai gruppi caucasoidi e
mongoloidi), mentre i gruppi viventi di pura caccia al modo dei nostri lontani antenati prima della
rivoluzione neolitica ed agricola contano complessivamente circa 300.000 persone.
Gli australoidi sono oggi in declino ovunque, tranne che presso le tribù aborigene dell'India, isole in
un mare umano caucasoide; i capoidi con le loro popolazioni boscimane ed ottentotte, contano solo
poche decine di migliaia di individui; i congoidi hanno invece dimostrato una straordinaria vitalità e
versatilità nell'assimilare le nuove culture, sia nei loro territori di origine, sia in tutti quei luoghi
ove, nell'ultimo millennio, sono stati sottomessi da caucasoidi e mongoloidi come uomini di fatica.
In ogni caso il flusso dei geni attraverso le zone di contatto tra le varie sottospecie e razze, nel corso
dell'ultimo mezzo milione di anni (periodo corrispondente a ventimila generazioni) sarebbe stato
sufficiente a rendere omogenea l'intera umanità, col risultato immediato di un’uniforme colorazione
cutanea color cachi chiaro e con lo sviluppo di similari strutture mentali e comportamentali, se tale
fosse stato lo schema evolutivo delle cose, e se non fosse stato vantaggioso per ognuna delle singole
sottospecie, oltre certo che facilitato dalle barriere geografiche, mantenere per la massima parte
intatti gli elementi adattativi del proprio status quo genetico.
Come le altre specie di esseri viventi, gli esseri umani sono, infatti, indirettamente e al profondo
guidati dai loro geni, e la propagazione di questi geni è stata da sempre la funzione ultima del
comportamento umano. Ad una visione disincantata, i diversi valori e le diverse culture risultano
quindi essere solo strumenti (ma strumenti capaci di riempire di significato un'esistenza) per
raggiungere questo obbiettivo. La maggior parte dei mutamenti che possono verificarsi in una
cultura già consolidata, come in campo biologico la maggior parte delle mutazioni genetiche, deve
essere dunque considerata deleteria, quando non indifferente o superflua. Così ci assicurano tanto i
nostri sentimenti che la nostra ragione.
I sistemi culturali che hanno dato prova di saper funzionare, e che trovano in sé stessi ampie e
sufficienti motivazioni di svolgimento biologico e storico, possono essere alterati dall'esterno solo
correndo rischi talora mortali.
Come ci si può allora sorprendere se i nostri antenati, meno innovatori di noi, furono caratterizzati,
per un'epoca che copre oltre il 99 per cento del divenire umano, da un sano conservatorismo
culturale e biologico; conservatorismo che solo noi, presuntuosi uomini della modernità, ci
permettiamo di definire sterile quando non immorale? Ed è questa una seconda conclusione che
possiamo trarre dallo studio integrato dell'evoluzione delle specie e dell'uomo fossile: che cioè, ci
piaccia o non ci piaccia, il meticciato intraspecifico ed interrazziale è stato sempre respinto a livello
di gruppo33 quando gli ambienti umani possedevano caratteristiche peculiari e naturali; che il fatto
di essere perfettamente adattata al proprio ambiente, senza possibilità di valido ricambio alternativo,
finisce per significare alla lunga la rovina sicura della specie; che la massima variabilità all'interno
di una specie non può quindi essere che un bene da tutelare con estrema attenzione e con estrema
prudenza, considerato che la variabilità ha sempre garantito le specie viventi dai mutamenti
32
Gozzoli S., Le radici e il seme, op. cit.
Ho cancellato dalle varie sottospecie con l'assorbimento dei caratteri in quel momento più deboli e con la
disgregazione dei sistemi di geni propri dei gruppi soccombenti. Per il concetto di sistema di geni cfr. von Bertalanffy,
op. cit., pp. 122. Ulteriore sottolineatura della complessa fragilità e dell'importanza della preservazione delle
caratteristiche di un patrimonio genico comune ad un «gruppo» (aperto a tutti i suoi membri, non cioè chiuso e
frazionato in caste, ma chiuso verso l'esterno), si ha in Dobzhansky, op. cit., pp. 250-251.
33
32
accidentali degli ecosistemi e che per quanto ci concerne l'umanità è senza dubbio ancora in
evoluzione, anche se non sappiamo verso cosa essa è diretta34.
Una terza conclusione, ma a questo punto il discorso può essere meglio ampliato con l'ausilio delle
discipline etologiche e sociobiologiche (e con una sana visione della storia che distrugga la
morfologia universalista ed il teleologismo informe delle speculazioni illuministe e marxiste)35, può
essere vista nel riconoscimento che le ideologie che vanno troppo contro le strutture profonde e
complesse della natura umana sono alla lunga destinate al fallimento, anche se purtroppo solo dopo
avere provocato danni talora irreparabili per il gruppo e sempre in ogni caso per l'individuo; e che
l'evoluzione culturale dell'uomo può progredire con successo e dignità solo se si trova in sintonia
con i suoi bisogni più reali fondati sull'infrastruttura biologica e temperati da un'autocoscienza
spoglia di veli ideologici e da una volontà conscia dei suoi limiti operativi.
È ben vero che l'uomo è l'unico animale che in grado sempre maggiore sta forgiando con le sue
mani il suo destino; ma è altrettanto vero che solo un'ignorante presunzione da padrone
dell'universo mondo può fargli supporre di essersi svincolato dalle norme più elementari che hanno
regolato per miliardi di anni la crescita e l'affermarsi della vita sulla Terra e di cui lo sviluppo
storico delle diverse società umane ci offre infiniti riscontri applicati. (G. Valli, 2005)
Marcello Andriola Dipartimento di Biologia Animale e Genetica “Leo Pardi”, Laboratori di Antropologia ed Etnologia Gruppo di Antropologia Cognitiva, Università di Firenze Via del Proconsolo 12 50122 Firenze (Italia) e‐mail:[email protected] 34
Profonde suggestioni nietzscheane ci fanno balenare alla mente (impropriamente per un saggio a sfondo biologico?)
la predicazione folgorante di Zarathustra: «L'uomo è un cavo teso tra l'animale e il superuomo, un cavo al di sopra di
un abisso. Un passaggio periglioso, un periglioso essere in cammino, un periglioso guardarsi indietro e un periglioso
rabbrividire e fermarsi. La grandezza dell'uomo è di essere un ponte e non uno scopo: nell'uomo si può amare che egli
sia una transizione e un tramonto». Nietzsche F., Così parlò Zarathustra, Adelphi, 1973, (2° ed.), p. 8.
35
von Bertalanffy, op. cit., pp. 305 e segg.: «La storia non è un processo che si sviluppa entro un'umanità amorfa,
nell'àmbito di un Homo sapiens inteso come specie zoologica». Con espressioni più radicali, ribadisce Darlington (in
Dobzhansky , op. cit., p. 55): «I caratteri innati ci fanno vivere in mondi diversi, anche se siamo fianco a fianco;
vediamo il mondo con occhi diversi, anche la parte che ne guardiamo insieme... I materiali ereditari dei cromosomi
costituiscono la sostanza solida che, in ultima analisi, determina il corso della storia».
33
Bibliografia
Anati E., 1992. Le radici della cultura, Jaca Book, Milano.
Andriola M., 2003. I principi antropologici della creatività, Pontecorboli, Firenze;
2004. Motivazione al successo, Pontecorboli, Firenze;
2006. Le origini neurobiologiche dell’ansia, Pontecorboli, Firenze;
2008. L’istinto costruttivo: Come l’uomo ha modificato la natura e se stesso, Pontecorboli, Firenze.
Ardrey R., 1968. L'istinto di uccidere, Feltrinelli, Milano.
Asimov I., 1983. Esplorando la terra e il cosmo, Mondadori, Milano.
Attenborough D., 1979. La vita sulla Terra, Rizzoli, Milano.
Balbi, R., 1993. Ebrei, razzismo e antisemitismo, Theoria, Roma.
Bastide, R., 1990. Noi e gli altri: I luoghi di incontro e di separazione culturali e razziali, Jaca Book, Milano.
Bateson G., 1995. Verso un’ecologia della mente, Adelphi, Milano;
1984. Mente e natura, Adelphi, Milano.
Beck U., 1999. Che cos’è la globalizzazione, Carocci, Roma;
2000. Il lavoro nell’epoca della fine del lavoro, Einaudi, Torino.
Campbell J., 1992. Mitologia creativa: le maschere di Dio, Mondadori, Milano.
Capra F., 2003. Il punto di svolta, Feltrinelli. Milano
Cavalli-Sforza Luigi Luca, 1996. Geni, popoli e lingue, traduzione di E. Stubel, Adelphi, Milano.
Chiarelli B., 1981. L'origine dell'uomo, Laterza, Bari.
1993. Bioetica globale, Pontecorboli, Firenze;
2003. Dalla natura alla cultura, Piccin, Padova;
2010. Systema Naturae, Vol. 10, Altravista, Pavia.
Colbert E.H., 1978. Animali e continenti alla deriva, Mondadori, Milano.
Coon C.S., 1970. L'origine delle razze, Bompiani, Milano.
Coppens Yves, 1987. I più antichi fossili ominidi, in L’evoluzione dei primati, Jaca Book, Milano.
Darlington C.D., 1973. L'evoluzione dell'uomo e della società, Longanesi, Milano.
Dawkins R., 1979. Il gene egoista, Zanichelli, Bologna.
Desmond A., 1979. L'enigma dei dinosauri, Newton Compton, Roma.
Dobzhansky Th., 1979. L'evoluzione della specie umana, Einaudi, Torino.
Eibl-Eibesfeldt I., 1976. I fondamenti dell'etologia, Adelphi, Milano;
1980. Amore e odio, Bollati Boringhieri, Torino;
2001. Etologia umana, Bollati Boringhieri, Torino.
Eiseley L., 1975. Il secolo di Darwin, Feltrinelli, Milano.
Reene G.J.C., 1971. La morte di Adamo, Feltrinelli, Milano.
Eliade M., 1997. Immagini e simboli, Editori Associati, TEA, Milano.
Eysenck H.J., Kamin L., 1981. Intelligenti si nasce o si diventa?, Laterza, Bari.
Larmat J., 1976. La genetica dell'intelligenza, Armando, Roma.
Paul C., 1982. Storia naturale dei fossili, Etas libri, Milano.
Facchini F., 1995. Antropologia. Evoluzione, uomo, ambiente, Utet, Torino.
Geertz C., 1987. Interpretazioni di culture, Il Mulino, Bologna.
Giddens A. 1999. Identità e società moderna, Ipermedium, Napoli;
2000. Il mondo che cambia, Il Mulino, Bologna.
Gliozzi, G., 1990. Teorie della razza nell'età moderna, Loescher, Milano.
Gozzoli S., 1981. Le radici e il seme, Edizioni dell'Uomo libero, Milano.
Grassè P.P., 1979. L'evoluzione del vivente, Adelphi, Milano.
Johanson D.C., Edey M.A., 1981. Lucy, le origini dell'umanità, Mondadori,
Kant E. 1910. Critica della ragion pura, Laterza, Bari.
Kluckhohn C., Kroeber A. L., 1982. Il concetto di cultura, Il Mulino, Bologna.
Kroeber A.L., 1983. Antropologia, Feltrinelli, Milano.
Lovelock J., 1981. Gaia: nuove idee sull’ecologia, Bollati Boringhieri, Torino;
1992. Gaia: manuale di medicina planetaria, Zanichelli, Bologna.
Mayr E., 1970. L'evoluzione delle specie animali, Einaudi, Torino.
Montagu, M.F.A., 1966. La razza. Analisi di un mito, Einaudi, Torino.
Morin E., 1974. Il paradigma perduto, Feltrinelli, Milano.
2001. I sette saperi necessari all’educazione del futuro, R. Cortina, Milano
Nietzsche F., 1973. Così parlò Zarathustra, Adelphi, Milano.
Pilbeam D., 1987. Evoluzione degli ominidi e origine degli ominidi, in L’evoluzione dei primati, Jaca Book, Milano.
Raven P. H., Johnson G. B., 1993, Biologia, Edises, Napoli.
Reeves H., 1982. L'evoluzione cosmica, Feltrinelli, Milano.
34
Ruffiè J., 1978. Dalla biologia alla cultura, Armando, Roma.
Ruse M., 1981. Sociobiologia, Il Mulino, Bologna.
Sheldrake R., 1993. La rinascita della natura, Corbaccio, Milano.
Tobias Phillip V., 1987. Evoluzione degli ominidi, Jaca Book, Milano.
Towsend C. R., Harper J. L., Begon M., 2001, L’essenziale di ecologia, Zanichelli Bologna.
Von Bertalanffy L. 1976. Teoria generale dei sistemi, Isedi, Milano.
35
Come pubblicare su Neuroscienze.net
Neuroscienze è una rivista on-line di informazione scientifica che tratta tematiche di
Neuroscienze, Psicologia e Scienze Cognitive.
Chi può collaborare?
Se sei un medico, un neurologo, uno psichiatra, uno psicologo, o se hai conoscenze
specifiche di neuroscienze, psicologia o scienze cognitive in genere, Neuroscienze ti offre
la possibilità di collaborare inviando i tuoi lavori.
Che percorso farà il tuo articolo?
Gli articoli ricevuti verranno considerati per la pubblicazione dall’Editorial Board e
successivamente inviati ai referee per la valutazione.
Come devono essere gli articoli?
Per poter essere pubblicato su Neuroscienze, il tuo lavoro deve rispettare le prescrizioni
contenute nella pagina “LINEE GUIDA PER GLI AUTORI“.
Come inviare il tuo articolo?
Per inviare il tuo articolo a Neuroscienze devi essere registrato al portale ed aver
effettuato l’accesso con username e password forniti al momento dell’iscrizione. A quel
punto potrai accedere direttamente alla zona riservata ed inserire autonomamente il tuo
articolo.
Linee Guida per gli Autori
Tutti i manoscritti sono soggetti a revisione redazionale. La presentazione di un articolo
per la pubblicazione NON implica il trasferimento del diritto d’autore da parte dell’autore
all’editore. Tutti i documenti sono pubblicati sotto Licenza Creative Commmons. E’
responsabilià dell’autore ottenere il permesso di riprodurre immagini, tabelle, ecc da altre
pubblicazioni.
Requisiti
Titolo, Autore e sottottotitolo: titolo, nome dell’autore e un sottotitolo sono necessari.
Parole chiave (keywords): per motivi di indicizzazione, un elenco di 3-10 parole chiave è essenziale.
Abstract: Ogni articolo deve essere accompagnato da un Abstract di un massimo di 10 righe.
Note: Evitare le note a piè di pagina. Quando necessario, numerarle consecutivamente e riportare le diciture
appropriate a piè di pagina.
Bibliografia: nel testo segnalare i riferimenti degli autori (cognomi ed anno di pubblicazione) tra parentesi.
L’elenco dei riferimenti deve essere in ordine alfabetico secondo il cognome del primo autore di ogni
riferimento. Il cognome di ogni autore è seguito dalle iniziali del nome. Si prega di citare tutti gli autori: ‘et
al.’ non è sufficiente. A questi devono seguire: l’anno tra parentesi, titolo, rivista, volume e numero delle
pagine.
Esempi:
Articoli pubblicati su Giornale: Gillberg, C. (1990). Autism and pervasive developmental
disorders. Journal of Child Psychology and Psychiatry, 31, 99–119.
Libri: Atkinson, J. (2000). The developing visual brain. Oxford: Oxford University Press Oxford Psychology
Series.
Contributi a Libri: Rojahn, J, e Sisson, L. A. (1990). Stereotyped behavior. In J. L. Matson
(Ed.), Handbook of behavior modification with the mentally retarded (2nd ed.). New York: PlenumPress.
Pubblica un Articolo
Dopo esserti registrato al portale, invia il tuo articolo dalla pagina:
http://www.neuroscienze.net/?page_id=1054