Fisiologia: studio della natura
(Fysis, natura e Lógos, discorso )
STUDIO DEL NORMALE FUNZIONAMENTO DI UN ORGANISMO VIVENTE E DELLE
PARTI CHE LO COMPONGONO
Studio delle funzioni vitali del corpo umano..di tutti gli animali e piante
Anatomia (Struttura) e Fisiologia (Funzione) sono strettamente correlate
I campi di studio della Fisiologia
Funzioni - Approccio Teleologico
(télos, fine e lógos, discorso )
Processi - Approccio Meccanicistico
sovrapposizioni
Omeostasi
La tendenza degli organismi viventi a mantenere in uno
stato di equilibrio le proprie caratteristiche al variare delle
condizioni esterne.
Termine coniato nel 1929 dal fisiologo americano Walter
B. Cannon. Homeo- ( simile) –stasis (nel senso di
condizione).
Parametri soggetti a controllo omeostatico
Fattori Ambientali
(osmolarità, Temperatura pH)
Materiali per le necessità cellulari
(nutrienti, acqua, sodio, calcio,altri ioni inorganici, ossigeno)
Secrezioni interne
(es ormoni)
Facoltà di Scienze Biotecnologiche
Corso di Laurea in Biotecnologie per l Innovazione di Processi e Prodotti
Programma del corso di Fisiologia ed elementi di Biofisica
(7+1)
Docente: Dott. Grazia Paola Nicchia
Parte I - Fondamenti di Biologia cellulare e Biofisica della cellula
Dinamiche di membrana. Struttura della membrana plasmatica. Il modello a mosaico fluido. Le proteine, i lipidi e i carboidrati della membrana. Proteoliposomi.
Tecniche per lo studio delle proteine di membrana.
Il trasporto di membrana di piccole molecole. Equazione generale dei trasporti. Forze agenti: potenziale chimico ed elettrochimico. Trasporti passivi: la
diffusione. Trasporto mediato dalle proteine di membrana. Trasporti attivi primari e secondari. Trasportatori ABC e farmaco resistenza. Trasporti mediati da
vescicole.
Parte II - Elettrofisiologia
Potenziale elettrochimico. Potenziale di diffusione. Potenziale di membrana a riposo.
Parte III - Comunicazione Cellulare e Trasduzione Dei Segnali Cellulari
Segnali elettrici. Eccitabilità. Potenziale d'azione. Basi ioniche dell eccitabilità di membrana. Proprietà molecolari e funzionali dei canali ionici voltaggiodipendenti. Eccitabilità della fibra nervosa, muscolare scheletrica e cardiaca. Propagazione dell impulso.
Segnali chimici. Trasmissione sinaptica. Sinapsi elettriche. Sinapsi chimiche: struttura, funzionamento e regolazione. Inibizione presinaptica. Integrazione
sinaptica. Il calcio e il rilascio di neurotrasmettitori. Neurotrasmettitori: classificazione, natura chimica e farmacologia. Modulazione dell attività sinaptica.
Farmaci e droghe per lo studio delle sinapsi. La tecnologia del DNA ricombinante per lo studio di alcune sinapsi. Accoppiamento elettro-meccanico: la
contrazione del muscolo scheletrico e cardiaco. Scossa muscolare. Unità motoria. Controllo della contrazione.
Trasduzione del Segnale. Recettori di membrana, secondi messaggeri, Ormoni, Fattori di crescita. Meccanismi d azione ed applicazioni biotecnologiche.
Parte IV - Distribuzione dell'acqua e dei soluti nell'organismo: equilibrio elettrico, chimico ed osmotico, osmosi e osmolarità.
Sistemi tampone del sangue e della cellula.
Parte V - Il sistema immunitario
Risposte anticorpali e risposte mediate da cellule.
TESTI CONSIGLIATI
SILVERTHORN
M. BERNE, M. LEVY
B. ALBERTS
H. LODISH:
Fisiologia Umana
Fisiologia
"Biologia Molecolare della cellula"
Biologia Molecolare della cellula
ED. AMBROSIANA
ED. ABROSIANA
ED. ZANICHELLI
ED. ZANICHELLI
Compartimenti liquidi funzionali del corpo
Le membrane del corpo
Sottile strato di tessuto che
riveste una cavità o separa due
compartimenti
Invenzione dei microscopi nel XVI secolo
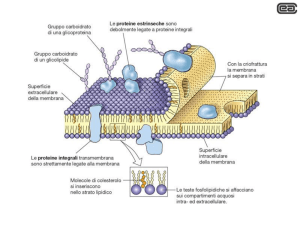
LA MEMBRANA PLASMATICA
Funzioni di una membrana cellulare
(membrana plasmatica o plasmalemma)
Funzione contenitiva
(Isolamento fisico)
Funzione di scambio
(Regolazione degli
scambi con l ambiente)
Funzione di riconoscimento
(comunicazione tra cellula
e ambiente)
Supporto strutturale
Struttura delle membrane
Film molto sottile di molecole lipidiche e proteiche tenute insieme da interazioni non covalenti
Figura 3.4
Le membrane cellulari sono strutture dinamiche e fluide
(Modello a mosaico fluido, Singer e Nicolson 1972)
Le molecole lipidiche sono disposte come un doppiostrato dello spessore di circa 5-8 nm
Il doppio strato lipidico forma la struttura base della membrana e funge da barriera selettiva
I LIPIDI
DELLA MEMBRANA
PLASMATICA
I lipidi costituiscono circa il 50% della massa delle membrane
FOSFOLIPIDI, COLESTEROLO e GLICOLIPIDI
I principali lipidi della membrana sono i
FOSFOLIPIDI (fosfogliceridi e sfingolipidi)
Colina
I FOSFOGLICERIDI
derivano da
Glicerolo
Abbiamo anche il Fosfatidil Inositolo coinvolto nella trasduzione del segnale
Gli SFINGOLIPIDI
derivano da
Serina
Variano in
dimensioni, forma
e carica
I principali lipidi della membrana sono i FOSFOLIPIDI o (fosfogliceridi)
Sono molecole anfipatiche e insature (legami cis)
Testa polare e due code idrocarburiche idrofobiche (14-24 atomi di C)
Possono avere uno o piu legami cis (sature o insature). Questo e la lunghezza degli acidi grassi
influenza la compatezza e quindi la fluidità
FOSFATIDILCOLINA
Comportamento dei fosfolipidi in soluzioni acquose
Le MICELLE
I LIPOSOMI
sono importanti nei
meccanismi di
digestione e
assorbimento dei
grassi nel TGI
possono contenere molecole
idrosolubili al loro interno.
Oggi sono usati per il
trasporto di farmaci
attraverso la cute.
La forma e la natura delle molecole lipidiche determinano il loro
comportamento in soluzione acquosa
Proprietà di autoriparazione
Fornita dalle stesse forze che spingono i fosfolipidi a formare doppi strati
Caratteristica fondamentale per per la creazione di una cellula vivente
Diffusione dei lipidi nel doppio strato (1970)
Preparazioni utili a questi studi
I liposomi
Le membrane nere
Si sono utilizzati lipidi marcati
Diffusione dei lipidi nel doppio strato
I lipidi non migrano da uno strato all altro. Questo processo (flip-flop) avviene solo durante
il processo di sintesi dei lipidi nel reticolo endoplasmatico.
I movimenti dei fosfolipidi all interno della membrana sono:
- diffusione laterale
- rotazione attorno al loro asse maggiore
- flessione
La fluidità del doppio strato lipidico
La fluidità di un doppio strato lipidico dipende:
1) dalla temperatura
2) dalla sua composizione
1) La membrana resta fluida a temperature più basse.
2) Più le catene lipidiche sono corte o hanno doppi legami e più le membrane sono fluide.
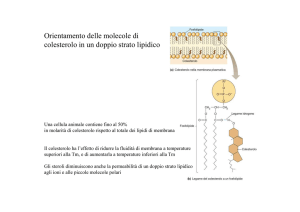
IL COLESTEROLO
Componente essenziale del doppio strato. Puo costituire fino al 50 % dei lipidi di membrana.
Serve a stabilizzare la membrana in condizioni di temperatura corporea normale
Il colesterolo rende i doppi strati lipidici meno fluidi
in quanto rende il doppio strato meno deformabile
in quelle regioni in cui esso interagisce con altri
fosfolipidi.
Fa così diminuire la permeabilità del doppio strato
a piccole molecole solubili in acqua
I GLICOLIPIDI
Rappresentano il 5% dei lipidi del monostrato esterno
Si trovano nella metà non citoplasmatica del doppi strato lipidico (distribuzione asimmetrica)
Funzioni
Nella membrana apicale degli epiteli possono avere
funzione protettiva (es. enzimi e pH).
I glicolipidi nelle cell animali derivano da serina
Glicolipidi carichi possono essere importanti per i loro
effetti elettrici.
Importanti nei processi di riconoscimento
(Ganglioside GM1, recettore di superficie per la tossina
colerica presente nelle cellule epiteliali dell intestino).
I glicolipidi piu complessi, i GANGLIOSIDI (circa una
quarantina), contengono oligosaccaridi con uno o piu
residui di acido sialico che conferisce carica netta
negativa. Nelle cell nervose rappresentano circa il
5-10 % dei lipidi.
(NANA= acido N-Acetil NeuroAminico)
Glicolipide Neutro
I lipid raft o zattere lipidiche
Piccole Aree Specializzate della Membrana (di circa 70 nm di diametri) in cui sono concentrati
Sfingolipidi, Colesterolo e Proteine
Gli sfingolipidi avendo catene
idrocarburiche lunghe creano delle forze
attrattive maggiori favorendo la
formazione di microdomini.
I lipid raft aiutano a concentrare proteine
(es per facilitare il trasporto in piccole
vescicole, o facilitando la trasmissione del
segnale)
Nei lipid raft i due monostrati sono
comunicanti tra loro
Asimmetria del doppio strato lipidico
(Membrana Plasmatica di globulo rosso)
EXT: Fosfatidilcolina e Sfingomielina
Il colesterolo si pensa sia
distribuito in modo uguale
nei monostrati
INT: Fosfatidiletanolamina e Fosfatidilserina
L asimmetria dei lipidi è funzionalmente importante: ex PKC richiede PS per la sua attività
Alcune funzioni dei fosfolipidi di membrana
PI = fosfatidilinositolo :
fosfolipide minore coinvolto nella
trasmissione del segnale
Legami fosfoesteri
Legami esteri
Fosfolipasi: Enzimi che tagliano i fosfolipidi
LE PROTEINE
DELLA MEMBRANA
PLASMATICA
Le proteine di membrana
Composizione chimica di alcune membrane (in % )
Membrana
Mielina
Eritrocita
Epatocita
Mitocondrialeinterna
Proteine
18
49
44
76
La quantità e i tipi di proteine in
una membrana sono altamente
variabili
Da un punto di vista anatomico si
dividono in
PROTEINE INTEGRALI
PROTEINE PERIFERICHE
PROTEINE ANCORATE AI LIPIDI
Lipidi
79
43
52
24
Carboidrati
3
8
4
0
Le proteine di membrana
Proteine Integrali di Membrana
(Transmembrana)
Proteine Periferiche di Membrana
Natura Anfipatica
Possono avere da 1 a 12 segmenti
Oligosaccaride legato al fosfatidilinositolo
Ancora di GPI (glicosilfosfatidilinositolo)
Rilasciate dalla fosfolipasi C
Alfa-elica anfipatica
Il legame covalente con
un lipide ne aumenta
l anfipaticità
Lipide con legame covalente.
Prodotte come proteine solubili
nel citosol
L organizzazione strutturale riflette la funzione
Interazioni non covalenti
Proteine Integrali di Membrana - Struttura α-elica
Struttura α-elica delle proteine di membrana
Le proteine transmembrana nella maggior parte attraversano la membrana in conformazione
α-elica). Poiché i legami peptidici sono polari e poiché l acqua è assente nel doppio strato, essi
formano tra loro legami idrogeno favoriti da tale conformazione. I legami idrogeno tra i legami
peptidici stabilizza la catena. L α-elica rappresenta il ripiegamento piu naturale che una catena
polipeptidica possa assumere.
Prozione Polipeptidica del Centro di reazione fotosintetico
Grafici di Idropatia
Difficili da cristallizzare e pertanto difficili da studiare per cristallografia ai raggi X
Grafici di idropatia misura della polarità di un residuo aminoacidico. Per localizzare potenziali segmenti
ad α-elica di una catena polipeptidica
Indice di idropatia: l energia necessaria per trasferire segmenti successivi di una catena polipeptidica
da un solvente NON polare all acqua. Il calcolo viene fatto per segmenti di 10-20 aa.
Valori positivi: è richiesta energia per il trasferimento all acqua – segmento idrofobico.
La Glicoforina A tipica proteina transmembrana ad attraversamento singolo
Piccola glicoproteina (30kDa) dei globuli rossi. Presente in forma dimerica. Abbondante
(106 copie per cellula).
La parte N-terminale è glicosilata. Funzione sconosciuta.
La Batteriorodopsina proteina transmembrana ad attraversamento multiplo
Pompa protonica attivata dalla luce – sette α-eliche.
Trasduttore di energia solare in energia utile alla cellula
Archeo Halobacterium Salinarum. Batteri
fotosintetici
Chiazze di membrana purpurea contenenti
molecole di batteriorodopsina
Il cromoforo in grado di assorbire
fotoni luminosi. In seguito al
legame con un singolo fotone si
eccita, determina un cambiamento
conformazionale della proteina
che trasporta H+ al di fuori della
cellula.
Ciò crea un gradiente di protoni a
cavallo della membrana
plasmatica che fornisce energia
alla cellula.
E identico al cromoforo nella
rodopsina delle cellule
fotorecettrici dell occhio dei
vertebrati.
Proteine Integrali di Membrana - Struttura a barili-β
Struttura a barili-β
• Modo alternativo per i legami peptidici nel doppio strato lipidico di soddisfare le loro richieste di formare legami
idrogeno.
• Formano strutture molto rigide e meno flessibili delle α-eliche e per questo sono abbondanti nella membrana
esterna dei mitocondri, dei batteri e e dei cloroplasti ma poco nelle cellule eucariotiche. Perché sono difficili i
cambiamenti conformazionali.
• Alcuni barili-β formano dei grossi canali transmembrana (le Porine)
Sito di legame
per il Ferro
Recettore per
un virus
batterico
Lipasi
Porina batterica
Proteina trasp ioni Ferro
Le Porine
Sono proteine transmembrana che formano pori e attraversano il doppio strato lipidico come un barile β
• Si trovano sulla membrana esterna dei batteri e permettono ai soluti idrofilici fino a 600 Da di
diffondere attraverso il doppio strato esterno - Il canale all interno è rivestito da catene aa polari.
• Anse che sporgono nel lume hanno la funzione di restringerlo in modo da renderlo selettivo solo
per alcuni soluti
• Ciascun monomero consiste di un barile β composto da 16 filamenti antiparalleli. Tutte le porine
sono proteine transmembrana trimericheI grafici di idropatia non possono identificare i segmenti
che attraversano la membrana come β-barili perché di solito meno di 10 aa sono sufficienti ad
attraversare il doppio strato lipidico.
• Tuttavia si fanno cristallizzare facilmente (struttura raggi X)
Proteine Periferiche Ancoraggio mediante gruppi idrocarburici
legati covalentemente
L attacco covalente tramite diversi tipi di lipidi serve alla localizzazione di
una proteina nella membrana dopo la sua sintesi nel citosol
Molte proteine sono ancorate agli sfingolipidi dei lipid raft
Attacco delle proteine alla membrana tramite lipidi
ANCORE GPI (Glicosil Fosfatidil Inositolo)
Molti proteoglicani
Carboidrati (numero variabile)
ANCORE ACILICHE
Es: v-Src (forma mutante di TK )
Gruppo acilico: miristato (C14) o palmitato (C16)
ANCORE PRENILICHE
Es: prot Ras e Rab
(famiglia di GTPasi)
Gruppo prenilico:
farnesile (C15) o geranilgeranile (C20)
Ancoraggio di proteine periferiche attraverso proteine integrali di membrana
NOTE:
La glicosilazione è presente sempre
sul lato non citoplasmatico della membrana.
L ambiente citoplasmatico riducente
non consente la formazione di legami
disolfuro.
I legami disolfuro dal lato extracellulare hanno un
ruolo importante nello stabilizzare la struttura ripiegata
della catena o la sua associazione con altre catene
polipeptidiche
Il Glicocalice
La superficie della cellula è ricoperta di zuccheri
che si trovano
- come catene oligosaccaridiche legate
covalentemente alle proteine e ai lipidi di
membrana
- come proteoglicani (lunghe catene
polisaccaridiche unite covalentemente ad un
nucleo proteico).
Ruolo del glicocalice :
Rivestimento Cellulare di un Linfocita colorato con Rosso Rutenio
1 - protezione contro il danneggiamento
meccanico e chimico
2 - mediare processi temporanei di
adesione cellula-cellula
Il glicocalice si visualizza con vari
coloranti come
il ROSSO RUTENIO o con LECTINE
marcate.
Le lectine sono proteine affini ai
carboidrati
Schema semplificato del glicocalice
Studio delle Proteine di Membrana
Approcci sperimentali
La membrana plasmatica dei globuli rossi
Vantaggi
Grande disponibiltà
Non hanno nucleo o organelli
(non c è contaminazione da altri tipi di
membrane)
Membrana facile da preparare:
I fantasmi dei globuli rossi:
Proteine della membrana plasmatica dei globuli rossi
Spettrina, glicoforina e banda 3 costituiscono più del 60% in peso delle proteine totali di
membrana dei globuli rossi.
Purificazione delle proteine
Purificazione delle
proteine globulari idrosolubili
Purificazione delle
proteine di membrana
Quando sono separate dalle
membrane queste proteine
mantengono la loro
conformazione nativa in un
mezzo acquoso poiché
presentano molti gruppi
idrofilici esposti
Quando sono separate dalle
membrane
le regioni idrofobiche di queste
proteine interagiscono
causandone l aggregazione in
un mezzo acquoso. Necessitano
di essere solubilizzate nei
DETERGENTI
Purificazione delle proteine di membrana – I DETERGENTI
I detergenti
Non ionici
Le moleclole di detergente sono
anfipatiche e tendono a formare
micelle in acqua
Idrofilica
Idrofobica
Ionici
CMC: Concentrazione micellare critica.
La CMC alla quale si formano le micelle è una
caratteristica di ogni detergente dipendente dalla
sua struttura
Agiscono in modi differenti a seconda delle diverse concentrazioni
Si legano alle regioni idrofobiche esposte delle prot di membrana e al nucleo idrofobico delle proteine
idrosolubili. A causa della carica questi detergenti distruggono anche i legami ionici e i legami idrogeno.
Denaturano le proteine
Solubilizzazione di proteine integrali di membrana
con detergenti NON ionici
Purificazione e ricostituzione della pompa Na+-K+
Occorre utilizzare
detergenti blandi!!
ELETTROFORESI
SDS PAGE e Western Blot (Immunoblotting)
SDS - PAGE
Western Blot (Immunoblotting)
Microscopia elettronica di crio-frattura
Freeze Fracture Electron Microscopy (FFEM)
Consente di studiare la morfologia della
membrana, la localizzazione e la
disposizione delle proteine di membrana
Le cellule sono congelate in azoto liquido e il blocco
congelato viene fratturato con un coltello
Microscopia elettronica di crio-frattura
Freeze Fracture Electron Microscopy (FFEM)
FFEM - Banda 3 e Glicoforina
La proteina tende a rimanere con il monostrato che contiene la massa principale della proteina
Le molecole di banda 3 restano in genere con la faccia P di frattura
Le molecole di glicoforina rimangono in genere con la faccia esterna (E) di frattura
- FFEM Visualizzazione di aggregati della proteina canale per l acqua Acquaporina-4
(AQP4)
E
P
Diffusione delle proteine
Le proteine di membrana non passano attraverso il doppio strato (flipflop) ma possono ruotare (diffusione rotazionale) o muoversi
lateralmente nella membrana (diffusione laterale) o flettersi (flessione).
Esperimento che dimostra che le proteine di membrana diffondono lateralmente
TECNICHE DI FOTOSBIANCAMENTO
MISURAZIONE DELLA VELOCITA DI DIFFUSIONE LATERALE DI UNA PROTEINA DI MEMBRANA
La proteina di interesse deve essere marcata con un anticorpo fluorescente o essere espressa come proteina
di fusione con una proteina fluorescente (es. GFP).
Si ricava il coefficiente di diffusione laterale della proteina che puo variare a seconda del tipo cellulare in cui
la proteina è espressa
FRAP
FRAP (Fluorescence Recovery After Photobleaching):
recupero della fluorescenza dopo fotosbiancamento
FLIP
FLIP (Fluorescence Loss In Photobleaching:
perdita di fluorescenza nel fotosbiancamento
- FRAP Velocità di diffusione laterale della proteina canale per l acqua Acquaporina-4
(AQP4)
Le cellule possono confinare proteine e lipidi in domini specifici della membrana
TIGHT JUNCTIONS NELLE CELLULE EPITELIALI
Nelle cellule epiteliali questa distribuzione asimmetrica è spesso essenziale per la
funzione dell epitelio stesso
Tre domini diversi nella stessa cellula spermatica
In questo caso esistono domini diversi senza la presenza di tight junctions.
Quattro modi in cui la diffusione laterale delle proteine di membrana può essere
ristretta
Le proteine si autoassemblano in grossi aggregati
Membrana purpurea dell halobacterium
Le proteine possono essere legate da
interazioni con complessi di
macromolecole all esterno (B) o
all interno (C) della cellula
Le proteine possono interagire con
proteine sulla superficie di un altra cellula