Biosintesi dei lipidi
La strategia di sintesi degli acidi grassi?
La sintesi degli acidi grassi comporta una serie di reazioni che seguono una strategia
diversa in vari modi dal corrispondente processo di degradazione:
Gli intermedi della sintesi degli acidi grassi sono legati covalentemente a gruppi
sulfidrilici delle proteine trasportatrici di acili (dall’inglese acyl carrier protein, ACP)
La sintesi degli acidi grassi avviene nel citosol
Gli enzimi della sintesi degli acidi grassi sono componenti di una lunga catena peptidica
detta acido grasso sintasi
Il coenzima per le reazioni di ossidoriduzione della sintesi degli acidi grassi è il
NADP+/NADPH
Le cellule devono fornire AcetilCoA e NADPH
Come vengono sintetizzati gli acidi grassi?
La biosintesi dei lipidi passa attraverso reazioni endoergoniche e riduttive, che
necessitano di ATP ed equivalenti riducenti, di solito sotto forma di NADPH,
ed ha come precursore il malonil-CoA, che non partecipa alla via degradativa:
-OOC–CH –CO–SCoA
2
Il malonil-CoA si forma dall’acetil-CoA e dal bicarbonato, con una reazione
irreversibile catalizzata dall’acetil-CoA carbossilasi
L’enzima è costituito da tre catene polipeptidiche (nei batteri), da un singolo
polipeptide trifunzionale (negli animali) o da entrambe le versioni
(nelle piante) ed è sempre biotina-dipendente
Le tre regioni funzionali sono:
1)
biotina carbossilasi,
2) proteina trasportatrice della biotina
3) transcarbossilasi
La reazione dell’acetil-CoA carbossilasi
La cinetica a ping pong
dell’acetil-CoA carbossilasi
Meccanismi di regolazione dell’acetil-CoA
carbossilasi
La proteina trasportatrice di acili
La proteina trasportatrice di acili (ACP) è la “navetta” che mantiene unito il sistema
ed ha come gruppo prostetico la 4’-fosfopanteteina
Esistono due varianti dell’acido grasso sintasi (fatty acid synthase, FAS):
Il tipo I si trova nei vertebrati e nei funghi, mentre il tipo II si trova nelle
piante e nei batteri
La FAS I dei vertebrati è una singola catena polipeptidica multifunzionale, che nei
mammiferi ha 7 domini differenti, ognuno con un proprio sito attivo:
KS, β-chetoacil-CoA sintasi
MAT, malonil/acetil-CoA-ACP trasferasi
DH, β-idrossiacil-ACP deidratasi
ER, enoil-ACP reduttasi
KR, β-chetoacil-ACP reduttasi
ACP, proteina trasportatrice di acili
TE, tioesterasi
La FAS II è costituita da proteine separate, è capace di produrre anche acidi grassi insaturi,
ramificati ed idrossilati e si trova anche nei mitocondri dei vertebrati
In tutti gli organismi gli acidi grassi vengono sintetizzati mediante una sequenza di quattro
reazioni enzimatiche ripetute, catalizzate dal sistema multienzimatico
dell’acido grasso sintasi:
1) Condensazione (con perdita di CO2)
2) Riduzione (NADPH-dipendente)
3) Deidratazione
4) Riduzione (NADPH-dipendente)
In ogni passaggio attraverso questa via anabolica, la catena dell’acido grasso
si allunga di due atomi di carbonio, donati dal malonato attivato
Ciclo di allungamento
della catena lipidica
Le FAS legano gruppi acetilici e malonilici attivati, catalizzando le seguenti reazioni fino
alla sintesi di acido palmitico (C16:0):
1) Condensazione (di Claisen) dell’acetile e del malonile, con formazione di acetoacetilACP, catalizzata dalla β-chetoacil-CoA sintasi (KS)
2) Riduzione del gruppo carbonilico sul C-3, catalizzata dalla β-chetoacil-ACP reduttasi
(KR) a spese del NADPH + H+
3) Deidratazione degli atomi di carbonio C-2 e C-3 con formazione del transΔ2-butenoil-ACP, ad opera della β-idrossiacil-ACP deidratasi (DH)
4) Riduzione del doppio legame con formazione di butirril-ACP da parte della
enoil-ACP reduttasi (ER) a spese del NADPH + H+
La proteina malonil/acetil-CoA-ACP trasferasi (MAT) e la proteina trasportatrice
di acili (ACP) permettono agli enzimi di ricevere i substrati necessari
alle varie reazioni
Per ragioni ancora poco chiare, l’allungamento della catena termina in genere dopo 7 cicli
di reazioni di condensazione e di riduzione, con il rilascio di acido palmitico ad opera della
tioesterasi (TE).
La biosintesi dell’acido palmitico
Nella prima fase si ha la formazione di 7 molecole di malonil-CoA:
7 Acetil-CoA + 7CO2 + 7ATP → 7Malonil-CoA + 7ADP + 7Pi
Nella seconda fase si hanno 7 cicli di condensazione e riduzione:
Acetil-CoA + 7Malonil-CoA + 14NADPH + 14H+ → Palmitato + 7CO2 + 8CoA
+ 14NADP+ + 6H2O
(Si noti che una molecola di H2O è utilizzata per idrolizzare il legame tioestere
del prodotto finale, il palmitato)
La reazione complessiva è quindi:
8 Acetil-CoA + 7ATP + 14NADPH + 14H+ → Palmitato + 8CoA + 7ADP + 7Pi
+ 14NADP+ + 6H2O
In molti organismi la sintesi degli acidi grassi avviene nel citosol,
mentre nelle piante avviene nei cloroplasti
Gli acidi grassi possono subire allungamento e
insaturazione
Gli acidi grassi saturi a catena lunga sono sintetizzati dal
palmitato, mediante l’aggiunta di unità acetiliche ad
opera del sistema di allungamento degli acidi grassi,
presente nel reticolo endoplasmatico liscio e nei
mitocondri
Introduzione di un singolo doppio legame cis nei batteri
Batteri: via O2-indipendente,
Eucarioti: via O2-dipendente
Introduzione di un singolo doppio legame cis
negli eucarioti
Vie di sintesi di altri acidi grassi
Le piante ed i batteri devono
sintetizzare acidi grassi poli-insaturi, per
garantire fluidità alle membrane alle
basse temperature
L’azione delle desaturasi
delle piante si esplica sugli acidi grassi
di un fosfolipide, non su quelli liberi
Il controllo regolatorio del metabolismo degli acidi grassi
Meccanismi di regolazione dell’acetil-CoA
carbossilasi
Il controllo ormonale del metabolismo degli acidi grassi
La sintesi dei glicerolipidi
Diacilglicerolo
e CDPdiacilglicerolo
sono i
precursori
principali dei
glicerolipidi
negli eucarioti
La sintesi di fosfatidilserina
La sintesi dei
trigliceridi
nell’intestino
Sintesi della
cardiolipina e del
fosfatidilinositolo
ATP sintasi
La biosintesi dei
plasmalogeni
La via “ciclica” dall’arachidonato (C20:4)
alle prostaglandine ed ai trombossani
La prostaglandina endoperossido H sintetasi
La via “lineare” dall’arachidonato ai leucotrieni
Il colesterolo
Il colesterolo è sintetizzato dall’acetil-CoA in quattro tappe:
1) Condensazione di tre unità di acetato, per generare mevalonato;
2) Conversione del mevalonato in unità isopreniche attivate;
3) Polimerizzazione di unità isopreniche (a 5 atomi di C), fino a dare lo
squalene (composto lineare a 30 atomi di C);
4) Ciclizzazione dello squalene per formare il nucleo a quattro anelli (A-D)
del ciclopentanoperiidrofenantrene
La biosintesi del colesterolo
La via di biosintesi del
colesterolo inizia nel
citosol con la sintesi del
mevalonato
La modulazione dell’attività della
HMG-CoA reduttasi
Dal mevalonato allo squalene
La
conversione
dello
squalene in
lanosterolo
porta alla
sintesi del
colesterolo
Il colesterolo è un componente essenziale delle lipoproteine
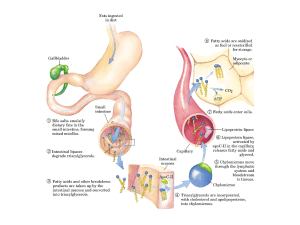
Lipoproteine e trasporto lipidico
Come viene trasportato il colesterolo
nel sangue?
Come viene trasportato il colesterolo
nel sangue?
Le lipoproteine
Assunzione del colesterolo attraverso il processo di endocitosi
mediata da recettore colesterolo nel sangue?
La biosintesi dei sali biliari
L’attività ossidasica a funzione mista
della 7α-idrolasi
Gli ormoni steroidei derivano dal colesterolo
La sintesi degli ormoni steroidei
Androgeni
Mineralcorticoidi
Estrogeni
Glucocorticoidi
Panoramica della biosintesi degli isoprenoidi