CIRCUITI CORTICALI
Cell. stellata
Cell. piramidale
Le afferenze di tipo associativo (tramite
fibre callosali) innervano invece
principalmente neuroni degli strati più
superficiali. Interneuroni inibitori (cellule
a canestro) permettono di delimitare
funzionalmente i moduli.
Le cellule della corteccia sono
organizzate in moduli. All’interno di
ogni modulo esistono dei circuiti che
permettono la trasmissione dalle
cellule stellate dello strato IV (che
ricevono le afferenze talamiche nel
caso di corteccie sensoriali) ai neuroni
piramidali dello strato III e V .
ORIGINE DEL SEGNALE EEG
L’ampiezza del segnale EEG é data dal prodotto tra I (corrente sinaptica) e
Re (resistenza extracellulare). Il segnale generato é quindi di ampiezza
ridotta rispetto al segnale sinaptico il cui valore é dato dal prodotto tra I e
Rm (resistenza di membrana).
La propagazione del segnale sinaptico genera in ogni cellula un campo
elettrico, che essendo descritto da un vettore ha intensità, direzione e
verso. Solo quando i vettori hanno la stessa direzione si sommeranno per
generare un segnale risultante non nullo.
La direzione del campo dipende dall’orientamento dell’albero
dendritico. Nel caso delle cellule piramidali i vettori sono paralleli e
quindi si sommano. Nel caso delle cellule stellate, data la loro
simmetria radiale il loro campo elettrico risultante è nullo.
IL POTENZIALE GENERATO DA UN DIPOLO
Solo segnali sincroni si sommano ed il
segnale EEG risulta dalla somma di tanti
campi elettrici generati in maniera
sincrona. I segnali sincroni sono quelli
sinaptici (lenti) e non i potenziali d’azione
(veloci).
Una eccezione a questo principio si ha nel caso di scariche sincrone di potenziali
d’azione (si verificano nel caso di attacchi epilettici).
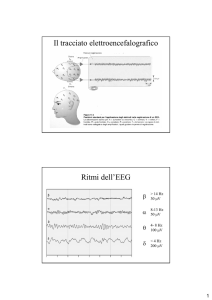
<3.5 Hz;
150 µV
4-7 Hz;
100 µV
8-15 Hz;
40-50 µV
16-30 Hz;
10-15 µV
SONNO
PROFONDO
SONNO
SUPERFICIALE
(nei bambini
anche
durante la
veglia)
VEGLIA
RILASSATA
VEGLIA
VIGILE
IL TRONCO DELL’ENCEFALO CONTROLLA L’ATTIVITA’ DEI NEURONI CORTICALI
ENCEFALO ISOLATO: NORMALE
RITMO SONNO-VEGLIA
PTB
SEZIONE MEDIO
PONTINA:
CERVELLO ISOLATO: SONNO
CONTINUO
IL RITMO SONNO VEGLIA DIPENDE DALL’ATTIVITA’ DI 2 CENTRI NERVOSI
CENTRO BULBARE DEL
SONNO
-
(INIBISCE IL
CENTRO
PONTINO)
CENTRO PONTINO DELLA
VEGLIA
LA PERFUSIONE SELETTIVA DEL BULBO (CENTRO DEL SONNO)
CON SOLUZIONE SALINA CONTENENTE UN BARBITURICO
CAUSA IL RISVEGLIO DELL’ANIMALE ADDORMENTATO. QUESTO
INDICA CHE L’ATTIVITA’ DEL CENTRO BULBARE E’ NECESSARIA
PER INDURRE IL SONNO. QUINDI IL SONNO E’ A TUTTI GLI
EFFETTI UN FENOMENO ATTIVO E NON UN FENOMENO
PASSIVO GENERATO DALL’ASSENZA DI STIMOLI SENSORIALI.
EFFETTO DEI BARBITURICI SULLA DURATA E SULLA STRUTTURA DEL SONNO
DIDASCALIA. Le cellule gangliari fotosensibili della retina (blu) rilevano le
variazioni cicliche dell’intensità luminosa e i loro assoni formano
connessioni con il nucleo soprachiasmatico (SCN), il principale orologio
biologico interno dell’organismo. SCN influenza l’attività del ganglio
cervicale superiore (SCG), il quale controlla la produzione ciclica, secondo
un ritmo di circa 24 ore (circadiano), della melatonina da parte della
ghiandola pineale (P). Gli assoni delle cellule gangliari fotosensibili
mandano collaterali anche al nucleo olivare del pretetto (OPN) e tramite la
proiezioni dal OPN al nucleo di Edinger-Westhphal (EW). E’ questa la
ragione per cui un certo grado di controllo del diametro pupillare è
presente anche quando i fotorecettori sono degenerati.