Principi di genetica - Robert J. Brooker
Copyright © 2010 – The McGraw-Hill Companies srl
SOLUZIONI AI PROBLEMI DEL CAPITOLO 25
Domande concettuali
C1.
I caratteri quantitativi sono descritti numericamente. Essi includono l'altezza, il peso, la velocità, e il tasso
metabolico. In una popolazione, a un carattere può essere attribuito un valore medio, e il grado di variabilità può essere
descritto dalla varianza e dalla deviazione standard.
C2.
A livello molecolare, i caratteri quantitativi di solito esibiscono una continuità nella variabilità fenotipica
perché sono influenzati da più geni che esistono in forme alleliche multiple. Anche una grande quantità di variabilità
ambientale aumenterà la sovrapposizione tra genotipi e fenotipi per i caratteri poligenici.
C3.
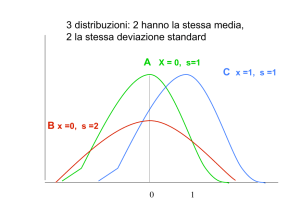
Una distribuzione normale varia in modo simmetrico attorno a un valore medio. Quando rappresentata in un
grafico, essa appare come una curva a campana. È comune che i caratteri quantitativi in una popolazione abbiano una
distribuzione normale attorno a un valore medio. Se una distribuzione è normale, la deviazione standard può predire la
percentuale degli individui che saranno compresi entro certi limiti. Per esempio, circa il 68% di tutti gli individui in una
popolazione cadono entro un'unità di deviazione standard (in direzione positiva oppure negativa) dalla media.
C4.
Un carattere discontinuo è un carattere che appartiene a categorie discrete. Esempi includono gli occhi
marroni e gli occhi blu negli esseri umani, e i fiori bianchi e porpora nelle piante di pisello. Un carattere continuo non
si distribuisce in categorie discrete. Esempi comprendono la statura degli esseri umani e il peso dei frutti nei pomodori.
La maggioranza dei caratteri quantitativi sono continui; i caratteri sono compresi in un intervallo di valori. La ragione
per cui i caratteri quantitativi sono continui è che essi sono solitamente poligenici e largamente influenzati
dall’ambiente. Come illustrato nella Figura 25.4b, questo tende a creare ambiguità tra genotipi e continuità fenotipica.
C5.
In genetica, una distribuzione di frequenza è un grafico che mette a confronto il numero degli individui e i loro
fenotipi per un determinato carattere. Questo è un modo tipico per descrivere una distribuzione di un carattere
quantitativo in una popolazione. Per costruire il grafico della distribuzione di frequenza per un carattere quantitativo
continuo, è necessario suddividere gli individui in gruppi arbitrari. Per esempio, se il carattere in considerazione è il
peso, gli individui potrebbero essere suddivisi in gruppi che siano multipli di 4,5 kg. Quindi si avranno gruppi di
individui nell'intervallo di 0 – 4,5 kg, 4,5 – 9 kg, 9 – 13,5 kg ecc. Per costruire una distribuzione di frequenza, si
sceglierà una popolazione di individui, e si determinerà il numero degli individui in ciascun gruppo. Le classi di peso
saranno riportate nel grafico in corrispondenza dell'ascissa, e il numero degli individui di ciascun gruppo sarà riportato
in ordinata.
C6.
Per ricadere nel 2,5% del bestiame più pesante ci si trova a circa due unità di deviazione standard. Se
calcoliamo la radice quadrata della varianza, la deviazione standard sarebbe 22 kg. Per essere nel 2,5% superiore un
animale dovrebbe pesare almeno 44 kg in più rispetto alla media, che equivale a 606 kg. Per essere tra lo 0,13% dei
bovini di peso inferiore, un animale dovrebbe essere tre deviazioni standard più leggero, che sarebbe almeno 66 kg
inferiore rispetto alla media, ossia 496 kg.
C7.
A.
Le differenze nella varianza possono essere spiegate con la variabilità con cui le patate sono influenzate
dalle variazioni ambientali, oppure potrebbero essere spiegate mediante differenze genetiche. La varietà con
varianza più elevata potrebbe avere una maggiore variabilità genetica.
B.
Da un lato, la varietà con la varianza più elevata potrebbe essere migliore perché il coltivatore potrebbe
voler selezionare individui quelle piante che producono patate più grandi. D'altro canto, se il coltivatore
volesse ottenere delle patate di dimensioni uniformi, la popolazione con una varianza più bassa sarebbe
migliore.
C.
Userei la varietà con la varianza più elevata, sperando che una proporzione significativa della varianza sia
dovuta alla variabilità genetica.
C8.
C’è una correlazione positiva, ma potrebbe dipendere semplicemente dal caso. Secondo la Tabella 25.2, questo
valore potrebbe essersi verificato per un errore di campionamento. Dovresti eseguire ulteriori esperimenti per
determinare se esiste una correlazione significativa, per esempio esaminare un numero maggiore di coppie di individui.
Se N = 500, la correlazione sarebbe statisticamente significativa, e concluderesti che la correlazione non sarebbe effetto
del caso. Tuttavia, non potresti dedurne una relazione di causa-effetto.
C9.
Un coefficiente di correlazione negativo significa che due variabili sono in relazione tra loro in modo inverso.
Principi di genetica - Robert J. Brooker
Copyright © 2010 – The McGraw-Hill Companies srl
Per esempio, un'abbondante nevicata in inverno potrebbe essere correlata in modo negativo alla sopravvivenza delle
specie selvatiche.
C10.
Quando un coefficiente di correlazione è statisticamente significativo, questo significa che l’associazione è
probabilmente avvenuta per ragioni diverse dall’errore di campionamento. Essa può indicare una relazione di causaeffetto ma non necessariamente è questa la spiegazione. Per esempio, parentali grandi possono avere progenie grande
per cause genetiche. Tuttavia, la correlazione può dipendere dalla condivisione di ambienti simili piuttosto che da una
reale relazione di causa- effetto.
C11.
L’eredità poligenica si riferisce alla trasmissione dei caratteri che governano due o più geni diversi. Questi
caratteri sono difficili da studiare, perché potrebbero esserci più geni con alleli multipli, e gli effetti ambientali
potrebbero essere significativi. Perciò, i genotipi discreti non ricadono in classi fenotipiche discrete. Invece, la tendenza
è alle sovrapposizioni, che confondono l'analisi genetica. Gli approcci molecolari, come la mappatura dei QTL,
possono contribuire aiutare a chiarire in modo significativo la situazione.
C12.
I loci di caratteri quantitativi sono localizzazioni cromosomiche che contengono i geni che determinano un
carattere quantitativo. È possibile che un QTL contenga un gene, oppure esso può contenere due o più geni strettamente
associati. La mappatura dei QTL, che comporta l’associazione con marcatori molecolari noti, è usata comunemente per
determinare le localizzazioni dei QTL.
C13.
A.
La probabilità di ereditare due alleli leggeri da un parentale sarebbe 0,5 x 0,5 = 0,25. La probabilità di ereditare
due alleli leggeri da entrambi i parentali sarebbe 0,25 x 0,25 = 0,0625, ossia 6,25%.
B.
La probabilità di ereditare tre alleli leggeri da un parentale sarebbe 0,5 x 0,5 x 0,5 = 0,125. La probabilità di
ereditare tre alleli leggeri da entrambi i parentali sarebbe 0,125 x 0,125 x 0,125 = 0,0156, ossia 1,56%.
C.
La probabilità di ereditare quattro alleli leggeri da un parentale sarebbe 0,5 x 0,5 x 0,5 x 0,5 = 0,0625. La
probabilità di ereditare quattro alleli leggeri da entrambi i parentali sarebbe 0,0625 x 0,0625 = 0,0039, ossia
0,39%.
C14.
Se l’ereditabilità in senso stretto equivale a 1,0, ciò significa che tutta la variabilità nella popolazione è dovuta
alla variabilità genetica piuttosto che alla variabilità ambientale. Questo non significa che l’ambiente non sia
importante nella manifestazione del carattere. In altre condizioni ambientali, il carattere potrebbe comportarsi in modo
abbastanza diverso.
C15.
L'ipotesi della dominanza propone che i ceppi inincrociati siano omozigoti per gli alleli recessivi dannosi
mentre gli ibridi non lo siano. L'ipotesi della sovradominanza suggerisce che alcune combinazioni alleliche siano
particolarmente favorevoli nell'eterozigote. Nel complesso, è difficile giudicare quale ipotesi sia corretta, sebbene i
genetisti osservino solo raramente sovradominanza in geni singoli. Un'eccezione è l'allele che determina in omozigosi
l'anemia falciforme, che abbiamo descritto in questo libro di testo. D'altra parte, la formazione degli alleli recessivi
dannosi è un fenomeno diffuso. Queste osservazioni tendono a supportare l'ipotesi della dominanza, ma la situazione
non è ancora chiarita.
C16.
Il vigore dell’ibrido è il fenomeno per cui un individuo della progenie prodotto da due ceppi inincrociati
presenta caratteristiche migliori rispetto ai corrispondenti parentali. I pomodori e il mais sono spesso ibridi.
C17.
Il principale vantaggio dell'incrocio selettivo è che esso permette di migliorare i caratteri quantitativi. Per
esempio, l'incrocio selettivo può ingrandire le dimensioni dei frutti, il numero dei frutti ecc. Uno svantaggio è che
l'incrocio selettivo diminuirà la variabilità genetica complessiva della popolazione, e questo può portare all'accidentale
prevalenza di caratteri indesiderati. Per esempio, quando un coltivatore di pomodori seleziona i pomodori più grandi e
succosi, egli involontariamente può ottenere una varietà con frutti più grandi e succosi, ma più suscettibile ai patogeni
rispetto al ceppo parentale originale (non selezionato).
I caratteri indesiderati, come la sensibilità ai patogeni, vengono spesso ereditati in modo recessivo. Questo è
dovuto ai rari alleli recessivi che diventano monomorfici per effetto casuale dell'incrocio selettivo. La creazione di
ibridi può permettere di superare gli effetti avversi dei rari alleli recessivi. Due diverse varietà che sono state sottoposte
all'incrocio selettivo in modo indipendente non saranno monomorfiche per gli stessi alleli recessivi. Perciò, quando due
varietà inincrociate vengono incrociate tra loro, gli ibridi diventano eterozigoti e portano per ogni gene una copia
dell'allele normale e una copia dell'allele recessivo deleterio. Se l'allele è recessivo, esso non manifesterà degli effetti
indesiderati sul fenotipo dell'organismo.
C18.
Quando una specie è sottoposta a incrocio selettivo, l’allevatore sta ponendo la sua attenzione sul
miglioramento di un particolare carattere. In questo caso, il coltivatore di rose si sta concentrando sulla dimensione e
qualità dei fiori. Siccome solitamente i coltivatori selezionano come parentali per la generazione successiva un piccolo
Principi di genetica - Robert J. Brooker
Copyright © 2010 – The McGraw-Hill Companies srl
numero di individui (per esempio quelli con i fiori migliori), questo può portare alla diminuzione della variabilità
allelica in altri geni. Per esempio, molti geni influenzano la profumazione nei fiori. In una popolazione non selezionata,
questi geni possono esistere nella forma allelica che determina la profumazione e in quella che non la determina. Dopo
molte generazioni di incrocio selettivo per i fiori grandi, gli alleli che rendono il fiore profumato possono venire persi
dalla popolazione, semplicemente per effetto del caso. Nell’incrocio selettivo questo è un problema comune . Quando
si seleziona per migliorare un carattere, è possibile diminuire inavvertitamente la qualità di un carattere non
selezionato.
Altri hanno suggerito che la mancanza di profumo possa essere collegata alla struttura e alla funzione del
fiore. Forse la quantità di risorse che un fiore usa per rendere i petali belli in qualche modo diminuisce la sua capacità
di produrre il profumo.
C19.
Quando si analizza un carattere in una popolazione, esso varia spesso tra i diversi individui. Talvolta la
variabilità è notevole, altre volte è limitata. La variabilità può essere causata dalle differenze genetiche tra i membri di
una popolazione, e essa può dipendere dal fatto che ciascun individuo della popolazione è esposto a un diverso
ambiente. La proporzione di variabilità che è determinata dalle differenze genetiche tra i membri della popolazione è
l'ereditabilità del carattere. Essa è applicabile solamente a una particolare popolazione che occupa un particolare
ambiente perché ogni popolazione presenta la propria quantità unica di variabilità genetica (sulla base dell'eredità
genetica dei suoi membri) e gli individui di una popolazione sono esposti a delle condizioni ambientali uniche.
C20.
L’ereditabilità in senso lato prende in considerazione tutti i fattori genetici che influiscono sulla variabilità
fenotipica di un carattere. L’ereditabilità in senso stretto considera solamente gli alleli che si comportano in modo
additivo. In molti casi, gli alleli che influenzano i caratteri quantitativi sembrano comportarsi in modo additivo.
Nondimeno, se un coltivatore assume che l’ereditabilità di un carattere sia dovuta agli effetti additivi degli alleli,
diventa possibile prevedere il risultato dell’incrocio selettivo. Questo viene anche definito ereditabilità realizzata.
C21.
A.
Falso. L'ambiente ha uno scarso impatto sulla variabilità del carattere. Tuttavia, l'ambiente contribuisce sempre
in modo considerevole alla manifestazione di ogni carattere. Non è possibile avere un organismo vivente senza
un ambiente.
B.
Vero.
C.
Probabilmente vero, ma non è possibile affermarlo con sicurezza. I caratteri quantitativi sono solitamente
poligenici.
D.
Falso. Non è possibile dire nulla circa l'ereditabilità del peso delle uova dei polli di un allevamento. Essa
dipende dalla quantità di variabilità genetica nella popolazione e dal tipo di ambiente nel quale sono stati
allevati i polli.
C22.
A.
A causa della loro buona nutrizione, puoi dedurre che essi diventeranno più alti.
B.
Se l’ambiente fosse abbastanza omogeneo, i valori di ereditabilità tenderebbero ad essere maggiori perché
l’ambiente contribuisce in modo minore alla variabilità del carattere. Perciò, nella comunità, l’ereditabilità può
essere maggiore, perché essi praticano in modo uniforme una buona condotta alimentare. D’altra parte, siccome
la comunità è di dimensione inferiore rispetto alla popolazione generale, la quantità di variabilità genetica può
essere minore, il che renderebbe inferiore l’ereditabilità. Tuttavia, siccome il problema afferma che la
popolazione della comunità è grande, potremmo assumere che la quantità di variabilità genetica sia simile a
quella della popolazione generale. Nell’insieme, la migliore previsione sarebbe che l’ereditabilità nella
popolazione della comunità è maggiore a causa degli standard nutritivi uniformi.
C.
Come affermato nella parte B, la quantità di variabilità sarebbe probabilmente simile, perché la comunità è
ampia. Come risposta generale, le popolazioni più grandi tendono ad avere maggiore variabilità genetica.
Perciò, la popolazione generale probabilmente ha un po’ più di variabilità.
C23.
In alcuni casi, gli alleli che si stanno selezionando diventeranno monomorfici nella popolazione, e l'ulteriore
selezione non produrrà alcun effetto. In alternativa, una seconda possibile spiegazione del limite di miglioramento è la
selezione naturale. Alcuni geni che rispondono alle selezione artificiale hanno un effetto negativo sulla fitness
complessiva di una popolazione. Si raggiunge un limite nel quale gli effetti desiderati sono bilanciati dagli effetti
negativi sulla fitness.
C24.
E’ probabile che una popolazione naturale abbia maggiore diversità genetica rispetto alla popolazione
addomesticata. Questo perché le popolazioni addomesticate sono state sottoposte per molte generazioni all’incrocio
Principi di genetica - Robert J. Brooker
Copyright © 2010 – The McGraw-Hill Companies srl
selettivo, che diminuisce la diversità genetica. Perciò, VG è probabilmente maggiore nelle popolazioni naturali. L’altro
punto è l’ambiente. È difficile affermare quale gruppo abbia un ambiente più omogeneo. In generale, le popolazioni
naturali tendono ad avere ambienti più eterogenei, ma non sempre. Se l’ambiente è più eterogeneo, questo tende a
causare una maggiore variabilità fenotipica che rende VE maggiore.
Ereditabilità
= VG/VT
= VG/(VG + VE)
Quando VG è elevata, l’ereditabilità aumenta. Quando VE è alta, l’ereditabilità diminuisce. Nella popolazione naturale
di lupi, ci aspettiamo che VG sia alto. Inoltre, possiamo prevedere che VE sia altrettanto alto (ma su questo c’è maggiore
incertezza). Tuttavia, se questo fosse vero, l’ereditabilità nelle popolazioni di lupi potrebbe essere simile alla
popolazione addomesticata. Questo perché VG nella popolazione di lupi sarebbe bilanciata dall’elevato valore di VE.
D’altra parte, se VE non fosse così elevata nella popolazione di lupi, o se fosse abbastanza alta nella popolazione
addomesticata, allora la popolazione di lupi avrebbe un’ereditabilità maggiore per questo carattere.
C25.
A.
Ipotesi della dominanza
B.
Entrambe le ipotesi
C.
Ipotesi della sovradominanza
Domande sperimentali
S1.
A.
Dobbiamo prima calcolare le deviazioni standard per l'altezza e il peso, e poi la covarianza per entrambi i
caratteri.
Altezza: varianza= 140,02; deviazione standard = 11,8
Peso: varianza = 121,43; deviazione standard = 11,02
Covarianza = 123,2
r(X,Y) = CoV(X,Y) / DSXDSY
r(X,Y) = 123,3 / (11,8)(11,02) = 0,948
B.
Con 8 gradi di libertà, questo valore è statisticamente significativo. Questo significa che l'associazione tra
queste due variabili avviene più frequentemente rispetto a quanto atteso in base all'errore casuale. Essa non
implica necessariamente un rapporto di causa-effetto.
S2.
Per calcolare la media, sommiamo i valori e li dividiamo per il numero totale
Media = [1,9 + 2 (2,4) + 2 (2,1) +3 (2,0) + 2 (2,2) + 1,7 +1,8 + 2 (2,3) + 1,6] / 15
Media = 2,1
La varianza è la somma delle deviazioni quadratiche dalla media, divisa per N – 1. Il valore medio di 2,1 deve essere
sottratto da ciascun valore, e poi se ne ricava la radice quadrata. Questi 15 valori vengono sommati e poi divisi per 14
(N – 1).
Varianza = 0,85 / 14
= 0,061
La deviazione standard è la radice quadrata della varianza.
Deviazione standard = 0,25
S3.
n = D2 / 8VG = (514 – 621)2 / 8(382) = 3,7
Ci sarebbe un numero minimo di quattro geni che influenzano la variabilità del peso di queste due varietà
bovine.
S4.
I risultati sono in accordo con l’idea che sono presenti dei QTL per questo carattere sui cromosomi 2 e 3, ma
non sul cromosoma X.
Principi di genetica - Robert J. Brooker
Copyright © 2010 – The McGraw-Hill Companies srl
S5.
L'ereditabilità riflette la quantità di variabilità genetica che influenza un carattere. Nella varietà con sette QTL,
ci sono almeno sette geni diversi con due o più forme alleliche che influenzano la manifestazione del carattere. In
un'altra varietà, vi sono gli stessi tipi di geni, ma tre di essi sono monomorfici e perciò non contribuiscono alla
variabilità del carattere.
S6.
Quando affermiamo che un RFLP è associato con un carattere, intendiamo che un gene che influenza un
carattere è strettamente associato con un RFLP. A livello cromosomico, il gene di interesse è così strettamente associato
con un RFLP che non avviene quasi mai un crossing-over tra loro.
Nota: ogni pianta eredita quattro RFLP, ma potrebbe essere omozigote per uno oppure due di essi.
Piccolo: 2700 e 4000 (omozigote per entrambi)
Medio-piccolo: 2700 (omozigote), 3000 e 4000; oppure 2000, 2700, e 4000 (omozigote)
Medio: 2000 e 4000 (omozigote per entrambi); oppure 2700 e 3000 (omozigote per entrambi); oppure 2000, 2700,
3000, e 4000.
Medio-grande: 2000 (omozigote), 3000, e 4000; oppure 2000, 2700, e 3000 (omozigote)
Grande: 2000 e 3000 (omozigote per entrambi)
S7.
Questi risultati suggeriscono che vi sono (almeno) tre geni diversi che influenzano la dimensione dei suini.
Questa è una stima minima perché un QTL può avere due o più geni strettamente associati. Inoltre, è possibile che le
razze piccole e grandi presentino, nell'analisi degli RFLP, una stessa banda associata a uno o più geni che determinano
le dimensioni.
S8.
Assumiamo che sia disponibile una mappa dettagliata di marcatori molecolari per il genoma del riso.
Potremmo iniziare con due ceppi di riso, uno con una grande resa e una con bassa resa, che differiscano
considerevolmente rispetto ai marcatori molecolari che portano. Dovremmo eseguire un incrocio tra questi due ceppi
per avere una F1 ibrida. Successivamente dovremmo reincrociare gli ibridi F1 con i due ceppi parentali, ed esaminare
centinaia di individui della progenie rispetto alla resa di riso e ai marcatori molecolari. In questo caso, i nostri risultati
attesi indicherebbero che sei diversi marcatori molecolari nel ceppo ad alta produzione sarebbero correlati alla progenie
con resa elevata. Se alcuni di questi geni fossero strettamente associati tra loro e allo stesso marcatore, dovremmo
ottenere meno di sei bande. Dovremmo ugualmente avere meno di sei bande se i due ceppi parentali avessero lo stesso
marcatore associato con uno o più geni che influenzano la resa.
S9.
Questa risposta assume che un topo sia omozigote per tutti gli alleli di suscettibilità affinché esso sia
suscettibile al virus della leucemia. Chiamiamo i geni per la resistenza virale V1, V2, V3, eccetera. V1 è un allele per la
resistenza, e v1 è un allele dello stesso gene che conferisce la suscettibilità. V2 è un allele di un gene diverso rispetto a
V1 e V2 conferisce la resistenza, mentre v2 conferisce la suscettibilità. Con queste premesse, calcoliamo la probabilità
che un topo F2 sia omozigote per tutti gli alleli di suscettibilità.
Se sono coinvolti due geni, l'incrocio parentale è
V1V1V2V2 x v1v1v2v2
Questo incrocio produce individui della progenie F1 sono V1v1V2v2.
Se costruisci un quadrato di Punnett, vi è una probabilità pari a 1/16 che un individuo F2 sia v1v1v2v2. Oppure
possiamo utilizzare la regola del prodotto. La probabilità che un parentale F1 trasmetta alla progenie un allele recessivo
di ciascun gene è ½. La probabilità che un topo erediti tutti i quattro alleli recessivi è ½ x ½ x ½ x ½ = 1/16.
Numero atteso di individui della progenie F2 suscettibile al virus = 1/16 x 120 = 7,5
Possiamo seguire lo stesso approccio per determinare la probabilità che la progenie sia omozigote per tutti gli
alleli recessivi se sono coinvolti tre o più geni.
Per tre geni: ½ x ½ x ½ x ½ x ½ x ½ = 1/64
Numero atteso di individui della progenie F2 suscettibile al virus = 1/64 x 120 = 1,9
Per quattro geni: ½ x ½ x ½ x ½ x ½ x ½ x ½ x ½= 1/256
Numero atteso di individui della progenie F2 suscettibile al virus = 1/256 x 120 = 0,5
Per una progenie di 120 individui, ci aspettiamo che circa due topi siano suscettibili al virus se sono coinvolti
tre geni. Perciò, tre geni tre geni sembrerebbe essere il numero di geni più probabile. Se non avessi ottenuto alcun gene
suscettibile, concluderemmo che siano coinvolti quattro o più geni. Sarebbe improbabile non ottenere topi suscettibili
Principi di genetica - Robert J. Brooker
Copyright © 2010 – The McGraw-Hill Companies srl
al virus se fosse coinvolto un numero pari o inferiore a tre geni.
S10.
A.
Se assumiamo che il ceppo altamente inincrociato non abbia varianza genetica,
VG (per il ceppo selvatico) = 3,2 g2 – 2,2 g2 = 1,0 g2
B.
hB2 = 1,0 g2 / 3,2 g2 = 0,31
C.
È lo stesso di hB2, quindi equivale a 0,31.
S11.
hN2 = R / S
R = X0 – X
S = XP – X
In questo problema:
X equivale a 1,01 (come indicato nel problema)
X0 equivale a 1,11 (calcolando il valore medio per i parentali)
XP equivale a 1,09 (calcolando il valore medio per la progenie)
R = 1,09 – 1,01 = 0,08
S = 1,11 – 1,01 = 0,10
hN2 = 0,08 / 0,10 = 0,8 (che è un valore di ereditabilità piuttosto elevato)
S12.
A.
hN2 = (X0 – X) / (XP – X)
0,21 = (26,5 g – 25 g)/ (27 g – 25 g)
X0 – 25 g = 2 g (0,21)
X0 = 25,42 g
B.
0,21 = (26,5 g – 25 g)/ (XP – 25 g)
(XP – 25 g) (0,21) = 1,5 g
XP = 32,14 g parentali
Tuttavia, siccome questo valore è molto lontano dalla media, ci potrebbero non essere dei parentali di 32,14 g nella
popolazione di topi che hai a disposizione.
S13.
In questo problema dobbiamo costruire i quadrati di Punnett sulla base dei genotipi.
A.
Denominiamo gli alleli A1 e A2. La F1 avrà un genotipo A1A2. I risultati della F2 saranno 1 A1A1 : 2 A1A2 : 1
A2A2. Dato che le piante A1A1 e A2A2 producono frutti di 0,45 kg e A1A2 producono frutti di 0,9 kg, ciò
produrrà un rapporto fenotipico pari a 50% 0,9 kg : 50% 0,45 kg.
B.
Denominiamo gli alleli A e a e B e b e assumiamo che essi contribuiscano in modo additivo. La progenie F1 sarà
AaBb. Il rapporto della F2 sarà
1 AABB (0,9 kg) : 2 AABb (0,9 kg) : 1 AAbb (0,67 kg) :
2 AaBB (0,9 kg) : 2 Aabb (0,67 kg) : 4 AaBb (0,9 kg) :
1 aaBB (0,67 kg) : 2 aaBb (0,67 kg) : 1 aabb (0,45 kg)
Il rapporto fenotipico sarà 9 (0,9 kg) : 6 (0,67 kg) : 1 (0,45 kg).
C.
Denominiamo gli alleli A1 e A2 e B1 e B2 e assumiamo che essi contribuiscano in modo additivo. La progenie 1
sarà A1A2B1B2. Il rapporto F2 sarà
1 A1A1B1B1 (0,45 kg) : 2 A1A1B1B2 (0,67 kg) : 1 A1A1B2B2 (0,45 kg) : 2 A1A2B1B1 (0,67 kg) : 2 A1A2B2B2 (0,67
kg) : 4 A1A2B1B2 (0,9 kg) : 1 A2A2B1B1 (0,45 kg) : 2 A2A2B1B2 (0,67 kg) : 1 A2A2B2B2 (0,45 kg)
Il rapporto fenotipico sarà 4 (0,45 kg) : 8 (0,67 kg) : 4 (0,9 kg).
Principi di genetica - Robert J. Brooker
Copyright © 2010 – The McGraw-Hill Companies srl
D.
Non è possibile calcolare un rapporto preciso. All'aumentare del numero dei geni, diventa più improbabile
essere eterozigote in tutti i loci quindi diventa meno probabile produrre frutti di 0,9 kg. Con un numero molto
grande di geni, la maggior parte degli individui F2 sarà produrrà frutti di un peso di valore intermedio (0,67 kg).
S14.
Dobbiamo prima calcolare i valori di a e b. In questo calcolo, X rappresenta la statura dei padri, e Y quella dei
figli maschi.
b = 92 / 72 = 1,28
a = 175 – (1,28) (173) = – 46,4
Per un padre alto 178 cm,
Y = (1,28) (178) + (– 46,4) = 181,4
L’altezza più probabile del figlio sarebbe 181,4 cm.
S15.
I gemelli monozigoti e dizigotici, che probabilmente condividono gli stessi ambienti, ma che si differenziamo
per la quantità di materiale genetico che condividono, sono un forte argomento a sfavore di una diversità ambientale.
Le differenze nelle correlazioni osservate (0,49 verso 0,99) sono in accordo con le differenze nelle correlazioni attese
(0,5 contro 1,0)
S16.
A.
Dopo sei o sette generazioni l’incrocio selettivo sembra avere raggiunto un plateau. Questo suggerisce che le
piante di pomodoro siano diventate monomorfiche per gli alleli che influenzano il peso dei pomodori.
B.
Sembra ci sia eterosi, perché la prima generazione ha un peso di 0,77 kg, maggiore sia rispetto ai pomodori di
Mary che a quelli di Hector. Questo spiega in parte perché Martin abbia ottenuto pomodori più pesanti di 0,68
kg. Tuttavia, l’eterosi non è l’unica spiegazione; essa non spiega perché Martin abbia ottenuto pomodori di 0,91
kg. Anche se i pomodori di Mary e Hector sono stati selezionati per il peso elevato, essi possono non avere tutti
gli alleli “pesanti” per ogni gene che controlla il peso. Per esempio, supponiamo ci siano 20 geni che
influenzano il peso, e ciascun gene esista nelle forme alleliche leggera e pesante. Durante le fasi iniziali di
incrocio selettivo, quando Mary e Hector hanno raccolto le loro 10 piante come produttrici di semi per la
prossima generazione, per effetto del caso alcune di queste piante potevano essere omozigoti per gli alleli
leggeri in alcuni dei 20 geni che controllano il peso. Perciò, per il solo effetto del caso essi probabilmente hanno
perduto alcuni degli alleli che determinano il peso elevato. Perciò, dopo 12 generazioni di incroci, questi
pomodori possiedono prevalentemente alleli pesanti, ma hanno anche gli alleli leggeri di alcuni geni. Se
rappresentiamo con la lettera maiuscola gli alleli pesanti, e gli alleli leggeri con la lettera minuscola, i ceppi di
Mary e Hector potrebbero essere:
Ceppo di Mary: AA BB cc DD EE FF gg hh II JJ KK LL mm NN OO PP QQ RR ss TT
Ceppo di Hector: AA bb CC DD EE ff GG HH II jj kk LL MM NN oo PP QQ RR SS TT
Come possiamo vedere, il ceppo di Mary è omozigote per gli allele pesanti a livello di 15 geni, ma presenta le
forme alleliche leggere per altri 5 geni. In modo simile, il ceppo di Hector è omozigote per 15 alleli pesanti e
porta l’allele leggero di altri 5. E’ importante notare, tuttavia, che gli alleli leggeri nei ceppi di Mary e Hector
non corrispondono agli stessi geni. Perciò, quando Martin li incrocia tra loro, egli inizialmente ottiene:
Progenie F1 di Martin: AA Bb Cc DD EE Ff Gg Hh II Jj Kk LL Mm NN Oo PP QQ RR Ss TT
Se gli alleli sono additivi e contribuiscono con identico peso al carattere, ci aspetteremmo circa lo stesso peso
(0,68 kg) perché questo ibrido ha un totale di 10 alleli leggeri. Tuttavia, se l’eterosi gioca un ruolo, i geni (che
erano omozigoti recessivi nei ceppi di Mary e Hector) diventeranno eterozigoti nella progenie F1, e questo può
rendere le piante più sane e determinarne un peso maggiore. Se il ceppo F1 di Martin viene sottoposto
all’incrocio selettivo, i 10 geni che sono in eterozigosi nella progenie F1 possono infine diventare omozigoti per
l’allele pesante. Questo spiegherebbe perché i pomodori di Martin hanno raggiunto un peso di 0,91 kg dopo
cinque generazioni di incrocio selettivo.
S17.
hN2 = ross / ratt
Il valore ratt deriva dalle relazioni genetiche note:
Madre/figlia: ross = 0,36
ratt = 0,5
hN2 = 0,72
Madre/nipote: ross = 0,17 ratt = 0,25
hN2 = 0,68
Sorella/sorella: ross = 0,39 ratt = 0,5
hN2 = 0,78
Principi di genetica - Robert J. Brooker
Copyright © 2010 – The McGraw-Hill Companies srl
Sorella/sorella (gemelli dizigotici): ross = 0,40
ratt = 0,5
Sorella/sorella (gemelli monozigotici): ross = 0,77 ratt = 1,0
hN2 = 0,80
hN2 = 0,77
L’ereditabilità media è 0,75.
S18.
A.
hN2 = (X0 – X) / (XP – X)
hN2 = (122 – 115) / (127 – 115) = 0,58
B.
0,58= (125 – 115) / (XP – 115)
XP = 132,2 kg
S19.
Questi dati suggeriscono che ci potrebbe essere una componente genetica nella pressione sanguigna, perché i
consanguinei dellepersone con la pressione sanguigna elevata sembrano avere un'alta pressione sanguigna.
Ovviamente, dovranno essere condotti sudi più approfonditi per determinare il ruolo dell'ambiente. Per calcolare
l'ereditabilità, la prima cosa da fare è calcolare il coefficiente di correlazione tra i gli inidividui consaguinei per vedere
se esso è statisticamente significativo. Se è così, allora potresti seguire l'approccio descritto nell'esperimento della
Figura EG25.2.2 (Esperimento di genetica 25.2 sul sito internet). Dovresti quindi determinare i coefficienti di
correlazione tra individui consanguinei per stabilire l'ereditabilità di un carattere. In questo approccio, l'ereditabilità
equivale a ross / ratt. Sarebbe importante includere coppie consanguinee che sono cresciute separatamente (per esempio
zii e nipoti) per vedere se essi hanno un valore di ereditabilità simile rispetto a coppie di individui geneticamente
correlati ma cresciuti nello stesso ambiente (per esempio fratelli e sorelle). Se i loro valori sono simili, questo ti
suggerirebbe che il valore di ereditabilità è determinato dalla genetica e non dovuto al fatto che parenti gli individui
consanguinei spesso condividono ambienti simili.