CAPITOLO 9
EREDITA’ ED EREDITABILITA’ DEI
CARATTERI QUANTITATIVI
LIGUORI EDITORE
1
• Gli studi genetici presentati hanno riguardato soprattutto
studi di sostituzione alleliche che causano differenze
qualitative a livello fenotipico.
• In realtà la variabilità tra organismi è, in generale, di
natura quantitativa piuttosto che qualitativa.
2
Caratteristiche quantitative
Molti caratteri dell’uomo e di altri organismi sono
geneticamente influenzati, ma non mostrano patterns di
ereditabilità a singolo gene (Mendelian).
Sono influenzati dalla combinazione di molti geni e sono
caratterizzati da variazione continua. Questi caratteri
sono chiamati poligenici.
I caratteri con variabilità continua sia poligenici che
influenzati da fattori ambientali sono chiamati
multifattoriali.
3
Passaggio dalla struttura genetica a quella fenotipica e la relazione che
esiste tra i due
Importanza per la comprensione dei processi evolutivi e per la gestione
delle specie rare
Studio di mutanti con effetti nel
fenotipo e varianti alleliche in un
singolo locus genico, confronto con il
selvatico e diretta relazione tra
genotipo e fenotipo (es. involucri dei
semi di pisello – gialli o grigi) =>
carattere discontinuo con pochi
fenotipi distinti
Caratteri che mostrano un’ampia
gamma di fenotipi possibili e che
presentano una distribuzione
continua di fenotipi => caratteri
continui
Relazione semplice tra genotipo e
fenotipo
4
Caratteri discontinui => pochi fenotipi distinti => descritti in termini qualitativi
Caratteri continui => molti fenotipi distinti => descritti in termini quantitativi
Effetto dell’ambiente
Relazione tra genotipo e fenotipo complessa perché:
- un genotipo può originare diversi fenotipi
- lo stesso fenotipo può essere originato da molti genotipi diversi
Come la variabilità tra individui per un dato carattere è
determinata durante lo sviluppo è il tema principale della
genetica quantitativa
5
Distinzioni tra caratteri qualitativi e
quantitativi
Caratteri “Qualitativi” :
gruppo sangigno (ABO)
colore del mantello del gatto
colore degli occhi
le differenze tra i fenotipi di
due individui possono essere
spiegati da differenze nel
genotipo ad un basso
numero di loci ( es. 1 o 2).
variabilità discontinua
rapporto mendeliano in F1
Caratteri Quantitativi :
altezza
peso
Il fenotipo è determinato,
con alcune eccezioni, dal
genotipo, ma differenze
fenotipiche tra gli individui
sono dovute a differenze
genotipiche a numerosi loci.
Variabilità continua
nessun rapporto
mendeliano in F1
6
I fenotipi multipli per un carattere possono insorgere
in più modi
1) Diversi fenotipi controllati da numerosi genotipi (all’aumentare del
numero di loci che controllano un carattere, aumenta il numero di genotipi).
Caratteri codificati da più loci sono detti caratteri poligenici
2) Quando i fattori ambientali esercitano un’influenza sul fenotipo, ogni
genotipo è in grado di produrre una gamma di fenotipi. Il fenotipo è
influenzato sia dai genotipi multipli che dai fattori ambientali => carattere
multifattoriale
7
L’altezza nell’uomo NON è un
carattere qualitativo
P
X
• L’altezza media non è il risultato
della segregazione di un singolo
gene. Molti pathway dello sviluppo
influenzano la statura.
• L’altezza è influenzata da fattori
ambientali. Le persone malnutrite
non sono tanto alte.
F1
8
Se si considerano due individui estremi
(es. una pianta di mais alta circa 2 metri e
una alta 90 cm) l’incrocio tra i due non
produrrà una progenie che soddisfa le
leggi di Mendel.
9
Tipi di tratti quantitativi
I caratteri quantitativi vengono di solito
descritti da misurazioni (quantità)
Ad esempio: quanto è alto?
Alcuni tratti quantitativi sono meristici (misurati in numeri
interi).
Un’altro tipo di carattere quantitativo è il carattere soglia
che può essere presente o assente in funzione di effetti
cumulativi di fattori additivi (malattie).
10
Fattori ambientali
Effetti sul fenotipo di
7 differenti genotipi di
Achillea a diversa
altezza.
11
Fattori ambientali
• Il numero di setole
addominali in
genotipi omozigotici
diversi di Drosophila
a tre temperature
diverse
12
Caratteri qualitativi
Attributi, che non possono essere
misurati, ma possono essere espressi
soltanto in modo qualitativo, ad esempio:
Caratteri a variabilità
discontinua
Classificazione degli
individui in due o
poche classi
chiaramente distinte
l’una dall’altra
13
Caratteri Quantitativi
Caratteri economicamente importanti
Multifattoriali (poligenici),
sono sotto il controllo di una pluralità di geni
Ruolo dell’ambiente sui caratteri quantitativi
Clima , terreno, pratiche colturali …
14
9.1
MISURA DEI CARATTERI QUANTITATIVI
I caratteri evidenti in una pianta hanno delle differenze tali da
essere distribuiti su una scala continua di valori
Caratteri quantitativi
Variano in modo continuo nella popolazione, non possono
essere classificati secondo classi discrete ma possono essere
misurati e descritti mediante parametri numerici
Caratteri poligenici con influenza dell’ambiente
La posizione occupata da questi geni sui cromosomi
corrispondono ai loci per i caratteri quantitativi e sono detti
QTL (Quantitative Trait Loci)
15
9.1
MISURA DEI CARATTERI QUANTITATIVI
Prime osservazioni caratteri qualitativi contrastanti (Mendel)
Successivamente caratteri quantitativi, misurabili (lunghezza della spiga)
Difficili da valutare perché la
variazione era continua, senza classi
continue ricollegabili a un gene
definito
Influenza dei fattori ambientali e
condizionamento del fenotipo
Due correnti di pensiero:
Mendeliani (Bateson, de Vries, Castle):
Biometristi (Galton e Person):
Caratteri a variabilità continua
(qualitativi) hanno una base genetica e
le differenze quantitative determinate
da unità discrete
Eredità per mescolamento, non
esistono le unità discrete alla base della
trasmissione ereditaria dei caratteri
16
9.1
MISURA DEI CARATTERI QUANTITATIVI
1) Esperimenti per dimostrare l’azione tra fattori genetici e ambientali nella
determinazione fenotipica dei caratteri quantitativi
2) Ipotesi multifattoriale dell’eredità quantitativa: i caratteri a variazione
continua sono controllati da molti geni con effetto additivo sulla
manifestazione fenotipica
Variabilità
fenotipica
Componente
genetica
Componente ambientale
(condizioni pedoclimatiche e
pratiche colturali)
17
Relazione tra genotipo e fenotipo
GENOTYPE
(GENES)
Developmental interections
Per comprendere le
regole base della
ereditabilità
bisogna mettere in
correlazione
genotipo e fenotipo
PHENOTYPE
18
Relazione tra genotipo e fenotipo
ENVIRONMENT
GENOTYPE
(GENES)
Developmental interections
PHENOTYPE
In altre parole l’ambiente e il genotipo
interagiscono per produrre il fenotipo.
19
Uno dei problemi principali della genetica quantitativa
consiste nel capire come distinguere quando la
variabilità per un carattere tra gli individui di una
popolazione è dovuta alla genetica, quando è di origine
ambientale
Vp = VG + VE
Come si misura la variabilità fenotipica?
Come si scompone la variabilità fenotipica nelle sue componenti?
Chiedere aiuto alla statistica
«il ruolo della metodologia statistica è essenziale nell’applicazione
del metodo scientifico»
Ercole Ottaviano 1995
20
Un metodo per riassumere i fenomeni di carattere
quantitativo è rappresentato dalla distribuzione di
frequenza, ovvero dalla descrizione di una popolazione
in termini di proporzione di individui che possiedono
ciascun fenotipo
Si contano quanti individui ricadono all’interno
dell’intervallo del fenotipo in questione
21
9.1
MISURA DEI CARATTERI QUANTITATIVI
Dati relativi a caratteri quantitativi = entità numeriche rappresentati mediante una
distribuzione di frequenza (= dividere il carattere in classi fenotipiche di un preciso intervallo)
Istogrammi mettono in relazione valori fenotipici e frequenze relative
Distribuzione di frequenza = curva di distribuzione normale (gaussiana) in cui il
carattere varia in modo simmetrico secondo una curva a campana (teorica)
Carattere quantitativo -> calcolo del valore centrale del campione e deviazione
attorno alla media (standard)
In teoria misure e campioni
dovrebbero coincidere
In pratica la dimensione della
popolazione è molto più elevata dei
dati di campionamento
Il campione è solo rappresentativo (->
campionamento casuale
22
sufficientemente grande)
9.1
MISURA DEI CARATTERI QUANTITATIVI
Media statistica = somma delle osservazioni diviso numero delle osservazioni
x=
(x1 + x2 + …. + xn )
n
Raccogliere osservazioni in gruppi (intervalli di classe di ampiezza stabilita a
priori), inserirli in un grafico ed estrapolare una distribuzione di frequenza
Moda = valore più frequente nel campione
Mediana = valore centrale di una serie
Distribuzione di frequenza consente di descrivere graficamente un carattere
quantitativo -> “Normale” con andamento simmetrico
Valore centrale = media
Ma come variano i valori all’interno dei campioni stessi?
Varianza = quanto i valori si discostano dalla media (più grande è, più i valori sono
23
dispersi)
Le tre misure con tendenza centrale più comuni:
• Media indica la media aritmetica che viene calcolata
sommando un insieme di numeri e quindi dividendo la
somma per la quantità dei numeri sommati. La media
di 2, 3, 3, 5, 7 e 10, ad esempio, è data dalla divisione
di 30 per 6, ovvero 5.
• Mediana indica il numero che occupa la posizione
centrale in un insieme di numeri, ovvero una metà dei
numeri ha un valore superiore rispetto alla mediana,
mentre l'altra metà ha un valore inferiore. La mediana,
ad esempio, di 2, 3, 3, 5, 7 e 10 è 4.
• Moda indica il numero più ricorrente in un insieme di
numeri. La moda, ad esempio, di 2, 3, 3, 5, 7 e 10 è 3.
24
In una popolazione i fenotipi di
individui per un carattere
quantitativo tende ad avere una
distribuzione normale
25
Distribuzione normale
La media:
+/- 1s = 66% dei valori
+/- 2s = più del 95%
Standard
Deviation
Mean
(average center of
distribution)
-5
-4
-3
-2
-1
0
1
2
3
4
5
26
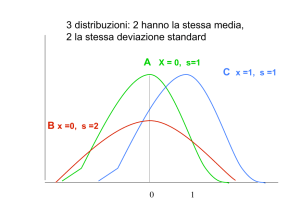
• Variabilità continua
µ
2
Frequenza (f)
Distribuzione normale
µ
Curve gaussiane aventi stessa media (µ)
ma diversa deviazione standard (σ).
-3σ
-2σ
-1σ
µ
+1σ
+2σ
+3σ
68,26%
95,45%
99,73%
27
9.1
MISURA DEI CARATTERI QUANTITATIVI
Varianza (s2 o 2) = rapporto tra le devianze (= somma delle differenze dei
valori osservati e della media, elevati al quadrato), diviso i gradi di libertà
Deviazione standard = si preferisce alla varianza ed è espressa nella stessa
unità di misura delle osservazioni (più grande è, più i valori delle osservazioni
si discostano dalla media, più la curva è bassa e larga)
s () =
s2
In una popolazione infinitamente grande, la distribuzione degli istogrammi è
rappresentata dalla caratteristica forma a campana
28
29
9.1
MISURA DEI CARATTERI QUANTITATIVI
Coefficiente di variabilità (CV) = rapporto percentuale tra la deviazione
standard e la media del campione
CV = s () 100
x
serve a comparare la quota di variabilità tra 2 campioni….
Altezza
(cm)
Peso (g)
x
53,59
23,28
s
10,85
8,50
CV
20,25
36,35
prendendo in considerazione solo le dev stand. sembrerebbe che i dati relativi
al carattere altezza siano più dispersi …. i CV indicano invece la distribuzione
del carattere peso a presentare una maggiore variabilità
30
Una difficoltà, quando di pensa in termini di fenotipo, è isolare i caratteri dal loro
contesto.
Studiarli singolarmente può essere artificiale perché gli organismi sono composti
da numerosissimi caratteri
Geni e fattori ambientali possono avere un effetto su diversi caratteri. Ad es. i
geni che controllano l’altezza possono essere pleiotropici sul peso e viceversa
Due o più caratteri sono spesso associati o
correlati
Se un carattere subisce un cambiamento, è
possibile che cambi anche l’altro
31
Covarianza (covariance) = misura se i valori di due set di campioni (genitori e
progenie) variano in maniera corrispondente (+), indipendente (0), o contrastante (-)
(xi – x) (yi – y)
Covarianza (cov xy) =
n-1
Il coefficiente di correlazione misura l’intensità dell’associazione tra due
variabili (x = lunghezza del corpo; y = larghezza del cranio)
Coefficiente di correlazione (r) =
Cov xy
s2x s2y
Può variare tra -1 e +1 e indica la direzione della correlazione
- Positiva (incremento di una variabile associata all’incremento dell’altra variabile)
32
- Negativa (incremento di una variabile associata al decremento dell’altra variabile)
(xi – x) (yi – y)
= 17979/19 = 946,26
Covarianza (cov xy) =
n-1
s2 (2) = (xi – x)/n-1 = 874/19 = 46,00
s2 (2) = (yi – y)/n-1 = 447148/19 = 23534,10
r=
946,26
= 946,26/1040 = 0,9095
46,00 x 23534,10
Tab. 8.10 – Valori significativi minimi del coeff. «r» per
diversi gradi di libertà e per livelli di P 1% e 5%
Gradi di
Libertà
P=0,05
P=0,01
18
0,4438
0,5614
50
0,2732
0,3541
100
0,1946
0,2540
33
Il coefficiente di correlazione indica l’importanza
dell’associazione tra due variabili e se la relazione è
direttamente o inversamente proporzionale, ma non dà
alcuna informazione sulla relazione tra le variabili. Questo
viene espresso dalla retta di regressione
Regressione (regression) = stima la misura in cui la varianza
fenotipica è dovuta a cause genetiche (attraverso la correlazione
tra generazione parentale e progenie): grossolanamente, è la
misura della diversità genetica ai caratteri quantitativi.
bxy = Covxy/VP
34
9.2 TAPPE FONDAMENTALI DELLA GENETICA
QUANTITATIVA
Studio dei caratteri quantitativi => Galton 1900
Johannsen 1909 -> influenza esercitata
dall’ambiente sull’espressione dei caratteri
quantitativi nelle linee pure di fagiolo
(autogama)
fattori
ambientali
fattori
genetici
Variabilità
Selezione
35
9.2 TAPPE FONDAMENTALI DELA GENETICA
QUANTITATIVA
Yule 1906 -> variabilità continua dei caratteri
quantitativi è legata a più coppie alleliche
segreganti insieme, senza alcuna spiegazione
Nilsson-Ehle 1908 -> influenza di fattori genetici
sulla variabilità di un carattere quantitativo in
frumento: più geni segreganti in modo
indipendente, ereditati in assenza di dominanza
con azione sul fenotipo nulla (geni “minus”) o
contributiva (geni “plus”) spiega i dati relativi al
grado di manifestazione di un carattere
quantitativo
Emerson e East 1913 -> variabilità di un carattere
quantitativo in linee inbred e ibridi di piante
allogame (mais) dipende solo da fattori ambientali
Variabilità fenotipica in F2 relativa alla variabilità
genetica conseguente alla segregazione e alla
36
ricombinazione
9.2 TAPPE FONDAMENTALI DELA GENETICA
QUANTITATIVA
East 1916 -> ipotesi multifattoriale dell’eredità
quantitativa in tabacco (autogama): caratteri
quantitativi sono controllati da molte coppie
alleliche a loci indipendenti e con azione uguale e
cumulativa (additiva) sul valore fenotipico
Analisi statistiche e citogenetiche dimostrano
come fattori genetici sono associati sugli stessi
cromosomi, manifestando inoltre effetti di
dominanza e interazione con altri fattori (epistasia)
Mather 1941 -> “poligeni” = fattori ereditari
coinvolti nel controllo genetico
“sistema poligenico” = insieme dei fattori che
controllano la variabilità continua dei caratteri
37
9.3 INFLUENZA DEI FATTORI AMBIENTALI SUI CARATTERI
QUANTITATIVI: Esperimenti di Johannsen
L’esperimento di
Johannsen nello
studio dei caratteri
quantitativi
38
Autofecondazione e omozigosi
Aa
Generazioni
segreganti
0,50
0,25
0,12
0,06
0,03
%
eterozigoti
%
omozigoti
1
1/2
1/2
2
1/4
3/4
3
1/8
7/8
4
1/16
15/16
5
1/32
31/32
m
1/2m
1-(1/2m)
0,50
0,75
0,88
0,94
0,97
Omozigosi
1-
1
2m
( )
n = N. di coppie
alleliche inizialmente
ibride
2m - 1
2m
(
2m - 1
2m
n
)
39
Percentuale degli omozigoti
1
n.
di coppie alleliche
inizialmente
ibride
0,9
0,8
0,7
1
0,6
2
0,5
5
0,4
10
0,3
100
0,2
0,1
0
1
2
3
4
5
6
7
8
9
10
n. di generazioni segreganti (m)
11
12
40
LINEA PURA
Insieme di individui
derivati per
autofecondazione
da un capostipite
Omozigote
41
Il risultato finale dell’autofecondazione degli
ibridi è una popolazione omozigote, ma non
omogenea perché costituita da famiglie
omozigoti per alleli diversi
Il numero possibile di famiglie omozigoti è
2n, dove “n” indica il numero delle coppie
alleliche inizialmente eterozigoti nell’ibrido
42
Lo studio dell’eredità dei
caratteri quantitativi è essenziale
per il miglioramento genetico
43
Wilhelm Ludvig Johannsen
(1857 - 1927)
• ha messo in evidenza l’azione
congiunta dei fattori genetici e dei fattori
ambientali nell’eredità dei caratteri
quantitativi.
Tra il 1903 e il 1909 egli realizzò
una serie di esperimenti allo
scopo di valutare l’eredità del
peso del seme in una specie
strettamente autogama come il
fagiolo (Phaseolus vulgaris)
44
Phaseolus vulgaris
45
Specie prev. autogama
9.3 INFLUENZA DEI FATTORI AMBIENTALI SUI CARATTERI
QUANTITATIVI: Esperimenti di Johannsen
Azione congiunta tra fattori genetici e fattori ambientali nell’eredità dei caratteri
quantitativi
Tipo di eredità del peso del seme nella pianta autogama di fagiolo (eterozigote a
tutti i loci)
Dimensioni diverse e relativo peso caratterizzato da variabilità continua
19 semi con peso compreso tra 64,2 cg e 35,1 cg
Autofecondazione -> linee pure
Semi prodotti da piante che provenivano da semi pesanti => peso medio alto
Semi prodotti da piante che provenivano da semi leggeri => peso medio basso
Differenze di natura
genetica tra le linee pure
della varietà
Variabilità entro ogni linea
dipendeva da fattori
ambientali
46
9.3 INFLUENZA DEI FATTORI AMBIENTALI SUI CARATTERI
QUANTITATIVI: Esperimenti di Johannsen
Dimostrazione (I esperimento) : i semi di ogni linea sono stati divisi in classi
di 10cg di ampiezza
piante
semi
Peso medio uguale a quello della linea pura da cui derivavano
Semi di grandezza diversa provenenti dalla stessa linea
pura danno origine a piante che producono semi con peso
medio caratteristico della linea di partenza
47
Princess
35,1 cg
64,2 cg
Linea 13
Peso medio
45,4 cg
selezione
dei
più pesanti
selezione
dei
più leggeri
selezione
dei
più pesanti
selezione
dei
più leggeri
Classi di peso
20-30-40-50
47,5 - 45,0 - 45,1 - 45,8
Entro una linea pura semi di grandezza diversa davano
progenie con peso caratteristico della linea
48
9.3 INFLUENZA DEI FATTORI AMBIENTALI SUI CARATTERI
QUANTITATIVI: Esperimenti di Johannsen
II esperimento: ogni linea pura moltiplicata per 6 generazioni (selezione e
moltiplicazione) ricorrendo ai semi più grandi e ai più piccoli
In ciascuna linea il peso medio dei semi della sottolinea leggera e pesante
sono risultati tra loro simili
Il peso medio dei semi in ogni linea rimane costante e la
selezione entro la linea pura è inefficace confermando che
la variabilità del carattere dipende solo da fattori ambientali
Coefficienti di correlazione tra pesi provenienti da ciascuna linea pura e
quelli prodotti dalle piante ottenute dai semi di ogni linea sono non
significativamente diversi da zero
49
Moltiplicazione entro ogni linea (fagioli grandi e
fagioli piccoli) per 6 generazioni
Linea 1 (64,3 cg)
Peso medio dei
semi delle progenie
Peso medio dei
semi dei genitori
Linea
leggera
Linea
pesante
Differenze
1902
60
70
+10
63,15
64,85
+1,70
1903
55
80
+25
75,19
70,88
-4,31
1904
50
87
+37
54,59
56,68
+2,09
1905
43
73
+30
63,55
63,64
+0,09
1906
46
84
+38
74,38
73,00
-1,38
1907
56
81
+25
69,07
67,66
-1,41
Anno
Linea
leggera
Linea
pesante
Differenze
Effetti della selezione continuata per sei generazioni entro la linea 1 della varietà
di fagiolo “Princess” (Johannsen, 1926)
50
Effetti della selezione continuata per sei generazioni
entro la linea 1 della varietà di fagiolo “Princess”
(Johannsen, 1926)
100
90
Genitori linea pesante
Peso in cg
80
Progenie linea leggera
70
Progenie
linea linea
pesante
GLL
Genitori
leggera
Genitori
linea pesante
GLP
Genitori
linea leggera
Progenie linea leggera
PLL
60
50
40
PLP
Progenie linea pesante
30
20
10
La selezione entro la linea
pura era inefficace
0
1
2
3
Anni
4
5
6
51
Coefficiente di correlazione
- Pesi dei semi scelti nell’ambito della cv. Princess
- Pesi medi dei semi prodotti dalle piante da essi ottenute
r = 0,34 (**) P 0,01
- Pesi dei semi presi all’interno della linea 13
- Pesi medi dei semi prodotti dalle piante da essi ottenute
r = 0,018 (ns)
• I semi della cv. Princess erano diversi (per cause ambientali e genetiche)
• Semi della linea 13 - geneticamente tutti uguali - il loro peso non influiva
sulla progenie
52
Princess
“Un insieme di
linee omozigoti
per alleli diversi”
la variabilità continua di un
carattere quantitativo è dovuta a
Cause genetiche
e
Cause ambientali
Rendono
la selezione
efficace
2P = 2G + 2E
Possono
ostacolare
la selezione
• Gli esperimenti di Johannsen dimostrarono
in modo inequivocabile l'effetto dei fattori
ambientali sui caratteri quantitativi.
• Effetto congiunto dell’eredità e dell’ambiente sul
carattere peso del seme:
La componente ambientale della variabilità (2E) era
legata a piccole differenze nelle condizioni interne ed
esterne della pianta che agivano sul seme
La componente genetica della variabilità (2G) era
legata alla OMOZIGOSI per alleli diversi nelle linee.
Princess
54
• Gli esperimenti di Johannsen non
dettero alcuna giustificazione della
variabilità continua di tali caratteri e
della conseguente impossibilità di
studiarli con il classico procedimento
basato sull'osservazione delle
segregazioni mendeliane.
55
9.3 INFLUENZA DEI FATTORI AMBIENTALI SUI CARATTERI
QUANTITATIVI: Esperimenti di Johannsen
Principi generali (base della variabilità genetica ancora poco chiara):
i) La variabilità fenotipica di un carattere quantitativo può avere una
componente genetica e una ambientale
ii) La selezione è efficace solo in presenza di variabilità genetica (linee pure
omozigoti per alleli diversi)
iii) La variabilità all’interno di linee pure è dovuta all’ambiente
iv) La selezione entro linee pure è inefficace
56
9.5 EREDITA’ DEI CARATTERI QUANTITATIVI: Esperimenti
di NILSSON-EHLE della cariosside in frumento
1908: l’osservazione del colore della cariosside in frumento fornì il primo
esempio naturale per spiegare l’eredità dei caratteri quantitativi
P
F1
F2
Linea pura a cariossidi
colorate (rosse)
×
Linea pura a cariossidi non
colorate (bianche)
cariossidi mediamente
colorate (rosse)
cariossidi colorate di
intensità variabile
×
cariossidi non
colorate (bianche)
57
Variabilità continua con tre loci
segreganti per due alleli
ABC ABc AbC aBC Abc aBc abC abc
ABC ABc AbC aBC Abc aBc abC abc
P1
Rosso
AA BB CC
X
Bianco
aa bb cc
F1
Colore intermedio
Aa Bb Cc
×
Rosso
Bianco
Fenotipo
Numero di alleli che producono colore 6 : 5 : 4 : 3 : 2 : 1 : 0
Rapporto
1 : 6 :15:20:15: 6 : 1
58
Nilsson-Ehle’s Model
Genotipo
Conteggio del
“rossore”
Colore
AABBCC
6
Rosso scuro
aabbcc
0
bianco
AaBbCc
3
Rosso medio
AABbcc
3
Rosso medio
59
P
F1
Le differenze tra:
• l’ereditabilità dei geni che influenzano i
caratteri quantitativi
• l’ereditabilità dei geni che influenzano i
caratteri qualitativi
sta nel numero di loci che determina il
carattere.
F2
- Sia i caratteri quantitativi (continui)
e quelli qualitativi (discontinui) sono
in accordo con le leggi di Mendel.
60
Geni e genotipi
Geni (n) Genotipi (3n)
1
2
3
(AA, Aa, aa)
9
3
27
5
243
10
59,049
Per n geni con due alleli
segreganti A e a, etc., il
numero totale di genotipi
dall’incrocio di n hybrid è 3n
Così per 5 geni segreganti ci
sono 243 genotipi
61
Cosi come il numero dei loci
influenzano gli aumenti del
tratto, il numero delle categorie
fenotipiche aumentano
N categorie fenotipiche =
[n(locus)× 2] +1
Collegando i punti delle
frequenze di distribuzione si
crea una curva a campana
chiamata distribuzione normale
62
9.5 EREDITA’ DEI CARATTERI QUANTITATIVI: Esperimenti
di NILSSON-EHLE della cariosside in frumento
3 diverse coppie alleliche e 3 loci responsabili del colore della cariosside:
A’A/B’B/C’C
A’, B’, C’
=> geni per il colore “rosso”
A, B, C
=> geni per il colore “bianco”
Geni “plus” (additivi)
-> contribuiscono alla manifestazione fenotipica
Geni “minus” (non additivi) -> non hanno effetti sulla colorazione della cariosside
Rispetto delle leggi di Mendel da parte di ciascuna coppia
63
9.5 EREDITA’ DEI CARATTERI QUANTITATIVI: Esperimenti
di NILSSON-EHLE della cariosside in frumento
Genotipo
A’A’
A’A
AA
Fenotipo
Rapporto fenotipico
Cariosside rossa
3
Cariosside bianca
1
Cariosside rossa intensità variabile
15
Cariosside bianca
1
Cariosside rossa intensità variabile
63
A’-B’A’-BB
AAB’AABB
A’-B’-C’…
…
Il fenotipo rosso era controllato da genotipi diversi e
l’intensità del colore era determinata dal numero di geni
“plus” determinando il risultato di un effetto cumulativo
64
9.5 EREDITA’ DEI CARATTERI QUANTITATIVI: Esperimenti
di NILSSON-EHLE della cariosside in frumento
Più coppie alleliche segreganti in modo indipendente (senza
dominanza e con azione uguale e additiva sul fenotipo),
potevano spiegare i risultati relativi al grado di espressione
del carattere nella generazione F2
Ogni allele per il rosso
aggiunge un certo grado di
colorazione alla cariosside
=> diversi genotipi a
seconda del numero di
loci in condizione
eterozigote in F1
65
9.5 EREDITA’ DEI CARATTERI QUANTITATIVI: Esperimenti
di NILSSON-EHLE della cariosside in frumento
Azione di due coppie alleliche a loci
indipendenti segregano in assenza di
dominanza e coinvolgono alleli plus con effetto
uguale e cumulativo sulla colorazione.
L’intensità del colore può essere derivata in base al numero di alleli
66
A’ e B’ presenti nel genotipo delle diverse piante
Calcolo della quota di un dato
genotipi in F2
• Quale proporzione della generazione F2 di un incrocio tra una pianta di
frumento a seme rosso ed una a seme bianco avrà due alleli dominanti?
• Assumendo 2 loci, si considerano i seguenti fenotipi:
– AaBb (1/2 x 1/2 = 1/4 or 4/16)
– AAbb (1/4 x 1/4 = 1/16)
– aaBB (1/4 x 1/4 = 1/16)
Considerando ogni gene si determina la proporzione di ogni classe
(legge del prodotto), quindi si sommano i tre insieme (legge della
somma).
– Risultato = 6/16
67
9.5 EREDITA’ DEI CARATTERI QUANTITATIVI: Esperimenti
di NILSSON-EHLE della cariosside in frumento
I fenotipi estremi sono quelli
più rari e i fenotipi intermedi
sono i più diffusi
P
F1
Distribuzione delle classi,
discontinua
Dimensioni di ciascuna
classe, ridotte
F2
Aumento del numero degli
alleli
Distribuzione normale
68
9.5 EREDITA’ DEI CARATTERI QUANTITATIVI: Esperimenti
di NILSSON-EHLE della cariosside in frumento
Quando un carattere quantitativo è controllato da molti geni è
impossibile distinguere le varie classi fenotipiche, perché l’effetto di
ciascun gene è troppo piccolo per essere discriminato
n
= coppie di alleli
2n+1
= numero di fenotipi in F2
(a+b)2n
= rapporti fenotipici attesi
L’effetto che l’ambiente esercita sulla
manifestazione del carattere quantitativo può
essere così importante da modificarne il valore
fenotipico.
69
9.5 EREDITA’ DEI CARATTERI QUANTITATIVI: Esperimenti
di NILSSON-EHLE della cariosside in frumento
P
F1
F2
Rosso intermedio
AAB’B’CC
Rosso intermedio
A’A’BBC’C’
X
Rosso intermedio rispetto ai parentali
A’AB’BC’C
×
Fenotipi di colore più estremi dei parentali
AA’B’B’C’C’
AABBCC
Quando si trovano nella generazione filiale piante con fenotipi più estremi
rispetto ai parentali, si parla di segregazione trasgressiva
La quota di variazione trasgressiva aumenta con il numero di geni coinvolti,
mentre la frequenza diminuisce all’aumentare della complessità dell’ibrido.
70
9.5 EREDITA’ DEI CARATTERI QUANTITATIVI: Esperimenti
di NILSSON-EHLE della cariosside in frumento
Ipotesi poligenica = l’eredità dei caratteri quantitativi dipende dalla
segregazione delle numerose coppie alleliche che possiedono effetti additivi
identici o quasi sul fenotipo e che non manifestano dominanza completa
i) In nessun locus uno degli alleli presenta dominanza sull’altro, ma ci saranno una
serie di alleli con effetti additivi (“plus”) e non additivi (“minus”)
ii) Ogni allele “plus” agisce in maniera cumulativa e ha uguale effetto sul fenotipo
iii) Gli alleli “minus” non contribuiscono a incrementare il fenotipo
iv) Non esiste interazione interallelica (epistasia)
v) I loci non sono associati, segregano in modo indipendente
vi) Non esiste variazione ambientale
i) -> iv)
v) NO => molti geni
vi) NO
71
9.4 INFLUENZA DELA COMPONENTE GENETICA SULLA
VARIABILITA’ DEI CARATTERI QUANTITATIVI:
Esperimenti di Emerson e East
Mais -> piante allogame. Misure della lunghezza della spiga raggruppabile in
classi discrete in funzione della lunghezza
Valutazione della variabilità del carattere “lunghezza della spiga” in piante:
- F1 ottenute dall’incrocio tra due linee inbred antagoniste
- F2 ottenute dall’inter-incrocio tra ibridi (<- effetti della segregazione e
ricombinazione)
Linee inbred parentali
- spiga corta
- spiga lunga
Lunghezza media di ibridi F1
compresa tra le lunghezze dei
parentali
Lunghezza media di ibridi F2 simile a F1,
ma con una variabilità fenotipica attorno
alla media più alta rispetto a F1
72
9.4 INFLUENZA DELA COMPONENTE GENETICA SULLA
VARIABILITA’ DEI CARATTERI QUANTITATIVI:
Esperimenti di Emerson e East
F2 variabilità fenotipica maggiore rispetto a F1 (valori estremi in F2 si discostano
maggiormente rispetto a F1)
MA la deviazione standard non può essere attribuita all’effetto della componente
ambientale
Aumento della variabilità fenotipica attorno alla media in F2 risiede
nella variabilità genetica dovuta agli effetti di segregazione e
ricombinazione
73
9.4 INFLUENZA DELA COMPONENTE GENETICA SULLA
VARIABILITA’ DEI CARATTERI QUANTITATIVI:
Esperimenti di Emerson e East
Principi generali (ambiente esercita la stessa influenza indipendentemente
dalla costituzione genotipica):
i) Incrociando due linee inbred antagoniste, il valore fenotipico medio in F1 è
compreso tra le medie fenotipiche dei parentali
ii) In F2 si osserva variabilità continua e i valori fenotipici non sono raggruppabili
secondo classi discrete
iii) Valore fenotipico medio in F2 è simile a F1
iv) F2 presenta una variabilità fenotipica attorno alla media del carattere
maggiore rispetto a entrambe le linee parentali ed F1
v) Valori fenotipici estremi di F2 si estendono verso le estremità della
distribuzione più di F1
vi) Variabilità nelle linee inbred e F1 dipende solo da fattori ambientali,
mentre l’aumento di variabilità in F2 è dovuta alla variabilità genetica
74
9.7 EFFETTO DELLA DOMINANZA SULL’EREDITA’ DEI
CARATTERI QUANTITATIVI
75
9.7 EFFETTO DELLA DOMINANZA SULL’EREDITA’ DEI
CARATTERI QUANTITATIVI
76
9.7 EFFETTO DELLA DOMINANZA SULL’EREDITA’ DEI
CARATTERI QUANTITATIVI
Quando le coppie alleliche che controllano un carattere quantitativo
manifestano dominanza (direzionale), la distribuzione della F1 avrà un valore
fenotipico spostato verso il parentale omozigote per gli alleli dominanti.
Nella F2 seguirà lo stesso andamento ed essendoci sempre meno eterozigoti,
tenderà a spostarsi i una posizione intermedia rispetto ai parentali omozigoti
iniziali.
77
9.7 EFFETTO DELLA DOMINANZA SULL’EREDITA’ DEI
CARATTERI QUANTITATIVI
All’aumentare del numero di coppie alleliche, la distribuzione diventa sempre
meno asimmetrica. Con più di 10 loci è difficile distinguere una distribuzione
influenzata da dominanza da quella ottenibile in presenza di effetti additivi
Per alcuni caratteri quantitativi la
variabilità fenotipica è dovuta a
differenze genetiche, per altri è
dovuta a cause ambientali
Effetto diverso nella F2
78
EREDITABILITA’ DI UN CARATTERE QUANTITATIVO
In una popolazione segregante o
variabile per un carattere,
lasciate che si incrocino individui
delle due code della curva di
distribuzione, mettete le loro
progenie in un ambiente comune
e controllato ed osservate :
79
EREDITABILITA’ DI UN CARATTERE QUANTITATIVO
In una popolazione segregante o
variabile per un carattere,
lasciate che si incrocino individui
delle due code della curva di
distribuzione, mettete le loro
progenie in un ambiente comune
e controllato ed osservate :
- Se le progenie seguono una
distribuzione che devia da quella
osservate nelle progenie
parentali andando oltre la fine
della curva dei loro parentali,
allora il carattere è ereditabile.
80
EREDITABILITA’ DI UN CARATTERE QUANTITATIVO
In una popolazione segregante o
variabile per un carattere,
lasciate che si incrocino individui
delle due code della curva di
distribuzione, mettete le loro
progenie in un ambiente comune
e controllato ed osservate :
- Se le progenie seguono una
distribuzione che devia da quella
osservate nelle progenie
parentali andando oltre la fine
della curva dei loro parentali,
allora il carattere è ereditabile.
- Se la progenie mostra la stessa
distribuzione della precedente
generazione, allora il tratto non è
ereditabile.
81
9.8 EREDITABILITA’ DEI CARATTERI QUANTITATIVI:
componenti della varianza fenotipica e genetica
È difficile riconoscere le classi
genotipiche a livello fenotipico quando
un carattere quantitativo è sotto l’effetto
di molti geni
Al contributo dei geni, bisogna
aggiungere anche l’effetto dell’influenza
ambientale
82
9.8 EREDITABILITA’ DEI CARATTERI QUANTITATIVI:
componenti della varianza fenotipica e genetica
Caratteri quantitativi sono sotto
l’effetto:
- Numero di geni
Distinzione mediante l’uso di
popolazioni sperimentali appropriate
- variazione ambientale tra individui
Caratteri qualitativi (monogenici) e quantitativi (poligenici con effetti ambientali)
Ereditabilità (h2) è la misura della variabilità fenotipica che risente del
contributo dei fattori genetici e ambientali.
E’ la frazione ereditabile della variazione osservabile per un carattere
quantitativo
E’ la proporzione di variabilità fenotipica di una popolazione attribuibile
a fattori genetici
83
9.8 EREDITABILITA’ DEI CARATTERI QUANTITATIVI:
componenti della varianza fenotipica e genetica
2P = 2G + 2E + 2GE
s2P = s2G + s2E + s2GE
Il contributo di ciascuno dipende da:
- struttura genetica della popolazione
- ambiente di coltivazione
- interazione tra i geni e l’ambiente
s2 G = s 2 D + s 2 H + s 2 I
La varianza genetica dipende da:
- effetto additivo
(più alleli a loci diversi con effetto cumulativo sul fenotipo)
- dominanza
(un allele può mascherare l’espressione dell’altro allele sullo stesso locus)
- epistasia
(interazione di alleli a loci diversi)
84
s2
P=
(s2
D
+
s2
H
+
s2 I) +
s2
E
+
s2
GE
IL CONTRIBUTO DELLA VARIABILITA’ GENETICA
AI CARATTERI QUANTITATIVI
la diversità genetica per i caratteri quantitativi può essere scomposta
nelle componenti che riflettono il potenziale evolutivo (VA), la
suscettibilità alla depressione da inbreeding (VD) e gli effetti
dell’outbreeding (VI)
VG = VA + VD + VI
VA: riflette il potenziale adattativo della popolazione per un determinato carattere
VD: riflette la dominanza tra alleli ai loci (quindi la possibile depressione da inbreeding)
VI: riflette le interazioni epistatiche tra alleli, quindi gli effetti dell’outbreeding (benefici o deleteri)
85
9.8 EREDITABILITA’ DEI CARATTERI QUANTITATIVI:
componenti della varianza fenotipica e genetica
2P = 2G + 2E + 2GE
s2P = s2G + s2E + s2GE
Il contributo di ciascuno dipende da:
- struttura genetica della popolazione
- ambiente di coltivazione
- interazione tra i geni e l’ambiente
s2E = s2Eg + s2Es + s2Em
La varianza ambientale dipende da:
- effetti ambientali generali
- effetti ambientali speciali
(colore della pelle dopo l’esposizione al sole)
- effetti materni
(dimensione alla nascita)
s2
P=
(s2
D
+
s2
H
+
s 2 I) +
(s2
Eg
+
s2
Es
+
s2
Em)
+
s2
86
GE
9.8 EREDITABILITA’ DEI CARATTERI QUANTITATIVI:
componenti della varianza fenotipica e genetica
0<
h2 =
s2 G
=
s2
h2 = 0
s2 G
P
>1
s2 G + s 2 E
variabilità fenotipica non è dovuta a differenze genetiche
h2 = 0,5 variabilità fenotipica dovuta per metà a differenze genetiche
per l’altra metà a cause ambientali
h2 = 1
variabilità fenotipica dovuta interamente a differenze genetiche
87
9.8 EREDITABILITA’ DEI CARATTERI QUANTITATIVI:
componenti della varianza fenotipica e genetica
Il ruolo dei geni e dell’ambiente varia da caso a caso
Quando tutta la variabilità è dovuta a differenze genetiche, il fenotipo è determinato
dal genotipo e ogni frequenza fenotipica può essere rappresentata da un
istogramma
Quando tutta la variabilità è dovuta anche a cause ambientali, le frequenze
fenotipiche sono rappresentate da una curva
Importanza dello studio dell’ereditabilità dei caratteri quantitativi nel miglioramento
genetico perché consente di formulare previsioni sul progresso conseguibile con la
selezione (quando esistono solo differenze genetiche e i fattori ambientali agiscono
in modo limitato)
88
9.9 EREDITABILITA’ IN SENSO LARGO E STRETTO
Scomporre la variabilità fenotipica per stabilire fino a che punto le variazioni tra
individui risultino da differenze genetiche
Ereditabilità in senso largo (h2B) considera la variabilità genetica dovuta a
qualsiasi effetto genico (additività, dominanza, epistasia)
h2B =
s2 G
s2 P
Prevede il confronto tra popolazioni geneticamente uniformi (linee pure, ibridi F1) e
variabili (F2 o interincrocio)
Stima della varianza ambientale
Stima in maniera cumulativa la
varianza fenotipica totale
h2B =
s2 G
=
s2 P
s2 P - s 2 E
s2 P
89
9.9 CALCOLO DELL’EREDITABILITA’
Basic generations
La varianza genetica può essere scomposta in
due quote: una attribuibile alle differenze tra i
valori riproduttivi (varianza genetica additiva ) e
l’altra alle deviazioni di dominanza (varianza
genetica non additiva)
La varianza fenotipica della F2 risulta perciò
scomposta in tre componenti
σ2
P
=
σ2 + σ2 + σ2
A
D
E
90
9.9 CALCOLO DELL’EREDITABILITA’
Basic generations
Due linee inbred: P1 e P2
Generazione dal loro incroci: F1, F2, B1 e B2
Calcolare la varianza e il valore medio
La varianza tra individui appartenenti alla stessa
linea inbred σ 2
σ2
P1
P2
e quella della F1 sarà:
σ2
F1
Sono stime della varianza attribuibile a cause ambientali per cui:
σ2
P1
+ σ2 + σ2 /3=
P2
F1
σ2
E
91
9.9 CALCOLO DELL’EREDITABILITA’
Basic generations
La varianza tra individui della F2 comprende quote
genetiche e quote ambientali
σ2
F2
2 + σ 2 +
σ
= A
D
σ2
E
La composizione della varianza dei due reincroci sarà:
σ2 + σ2=
B1
B2
σ 2 + 2 σ 2 + 2σ 2
A
D
E
92
9.9 CALCOLO DELL’EREDITABILITA’
Basic generations
La soluzione del sistema seguente consente la
stima dei parametri genetici (le incognite)
σ 2 = σP12 + σ 2 + σ 2 / 3
E
σ 2 = σA2 +
P2
σ2
D
σ2
= σ
+
σ2 +
σ2
σ2
= σ
+
σ2 +
σ2
F2
B1
B2
2
A
2
A
F1
+
D
D
σ2
E
E
E
93
9.9 CALCOLO DELL’EREDITABILITA’
Basic generations
Moltiplicando per 2 l’equazione relativa alla F2 e
sottraendo l’equazione 1) alla equazione 2) si
ottiene la stima della varianza additiva
σ2
F2
=
2
2σ =
F2
σ2
A
2σ 2 +
A
σ2 + σ2=
B1
+
B2
σ2 +
σ2
D
E
2σ 2 +
2σ 2
D
σ2
A
+2
E
σ2
D
+ 2
σ2
E
94
Ereditabilità
(in senso largo)
Porzione della varianza totale di una
popolazione dovuta alla varianza genetica
Indipendente
ambiente
GENI
P=G+E
Valore
carattere
Phenotype = Genotype + Environment
95
Come si quantifica l’ereditabilità?
se P = G + E
H2 = G/(G + E) = G/P
(ereditabilità in senso lato)
1 > H2 > 0
L’ereditabilità è specifica per una popolazione e l’ambiente;
l’ereditabilità in un contesto non è la stessa rispetto ad
un altro contesto (popolazione + ambiente)
96
9.9 EREDITABILITA’ IN SENSO LARGO E STRETTO
Scomporre la variabilità fenotipica per stabilire fino a che punto le variazioni tra
individui risultino da differenze genetiche
Ereditabilità in senso largo (h2B) considera la variabilità genetica dovuta a
qualsiasi effetto genico (additività, dominanza, epistasia)
h2B =
s2 G
s2 P
=0
=> nessuna parte della variabilità fenotipica risulta da differenze genetiche
= 0.5
=> il 50% della variabilità fenotipica deriva da differenze genetiche
=1
=> tutta la variabilità fenotipica è dovuta a differenze genetiche
Ereditabilità in senso largo ignora il fatto che la varianza genetica possa essere
additiva, dominante o epistatica.
L’interazione genotipo-ambiente non è importante 97
9.9 EREDITABILITA’ IN SENSO LARGO E STRETTO
Ereditabilità in senso stretto (h2N) considera solo le differenze genetiche
attribuibili alle azioni geniche additive, ovvero le uniche a essere fissate con la
selezione perché rimangono inalterate nelle generazioni successive (le uniche
legate all’effetto medio dei geni)
Non viene considerata la dominanza e l’epistasia perché non sono fissate con la
selezione perché risentono dell’effetto della ricombinazione e della segregazione e
quindi variare da una generazione all’altra.
2D
h2 N =
2P
98
9.9 EREDITABILITA’ IN SENSO LARGO E STRETTO
Il calcolo dell’ereditabilità prevede il confronto tra:
A) popolazioni non imparentate (individui parentali e quelli della relativa discendenza
– parents-offspring)
B) popolazioni imparentate (individui della stessa progenie con uno o entrambi i
parentali in comune - half-sibs, full-sibs)
Base sperimentale:
- costituzione genotipica => individui imparentati devono essere anche
fenotipicamente simili, perché condividono molti geni
- costituzione ambientale => individui imparentati e geneticamente simili non
dovrebbero manifestare fenotipi più simili dei non imparentati
Gli individui imparentati studiati non devono condividere anche un ambiente
omogeneo, altrimenti risulterà difficoltoso dividere gli effetti genetici da quelli
ambientali
99
9.9 EREDITABILITA’ IN SENSO LARGO E STRETTO
Punti casuali
=> nessuna relazione tra i fenotipi dei parentali e della progenie
(effetti genici additivi molto bassi)
=> ereditabilità BASSA
=> h2N = 0
differenze fenotipiche attribuibili a effetti di dominanza e o
epistasi o a un forte potere ambientale
100
9.9 EREDITABILITA’ IN SENSO LARGO E STRETTO
=> Relazione tra i fenotipi dei parentali e della progenie
(effetti genici additivi importanti)
=> ereditabilità ALTA in senso stretto
=> h2N = b -> h2 = 1
=> nessun fattore ambientale comune a genitori e progenie
influenza il carattere
fenotipo della progenie è intermedio rispetto ai parentali
101
9.9 EREDITABILITA’ IN SENSO LARGO E STRETTO
0 < h2 < 1 fenotipo della progenie è dovuto a geni con effetto additivo e a fattori con
effetti non additivi (dominanza, epistasia, fattori ambientali)
102
9.9 EREDITABILITA’ IN SENSO LARGO E STRETTO
Nonostante l’utilità pratica, l’ereditabilità presenta alcuni limiti:
i) Non equivale a quanto l’espressione di un carattere dipende da fattori genetici,
ma esprime la proporzione della varianza fenotipica attribuibile a differenze
genetiche
ii) È una caratteristica della popolazione e non del singolo individuo
iii) Non è fissa ma dipende dalla composizione di un gruppo di individui in un
ambiente preciso
iv) Non può essere usata per trarre conclusioni riguardo la natura di differenze
genetiche tra popolazioni
v) Caratteri condivisi da membri della stessa famiglia non possiedono
necessariamente un’elevata ereditabilità. Quando succede, il carattere è familiare.
Ma l’ereditabilità non è la familiarità
103
Familiarità ed ereditabilità
Si dice che vi è Familiarità per un dato carattere se i membri della
stessa famiglia lo condividono, per qualsiasi ragione.
– I caratteri sono familiari per cause genetiche ed ambientali (per
esempio la religione è familiare ma non genetica)
Si parla di Ereditabilità di un carattere solo se la somiglianza deriva
dal condividere lo stesso genotipo
- L’altezza, il colore dei capelli, il colore della pelle sono caratteri
ereditabili.
104
9.9 EREDITABILITA’ IN SENSO LARGO E STRETTO
Nel miglioramento genetico, la variabilità genetica può portare a miglioramenti nella
resa, resistenza, dimensioni, qualità agronomicamente importanti
Evoluzione = variazione genetica nel tempo entro gruppi di organismi (importanza
nel poter prevedere la velocità e la grandezza della variazione genetica che
avverrà)
Importanza della selezione artificiale = sono gli uomini che selezionano gli individui
che dovranno sopravvivere e riprodursi. Se i caratteri selezionati avranno una base
genetica, anch’essi varieranno e si evolveranno
Selezione = mezzo per produrre un rapido cambiamento evolutivo (addomesticazione)
105
9.9 EREDITABILITA’ IN SENSO LARGO E STRETTO
Selezione naturale (Darwin) e artificiale (praticata da allevatori) dipendono dalla
presenza di variabilità genetica.
Il tipo e la quantità di variabilità genetica sono importanti nel determinare a quale
velocità avverrà l’evoluzione
Quando su un fenotipo si applica selezione, il fenotipo cambierà da una
generazione all’altra, a patto che esista nella popolazione variabilità genetica per il
carattere.
Risposta alla selezione =
quantità di variazione per il
fenotipo in una generazione
Risposta alla selezione =
Differenziale di selezione =
differenza tra fenotipo medio dei
genitori selezionati e il fenotipo medio
della popolazione non selezionata
ereditabilità in senso stretto
× differenziale di selezione
106
Per aumentare la dimensione corporea della D. melanogaster si avvia un
programma di selezione:
A) si esaminano i moscerini da una coltura geneticamente eterogenea e si
misurano le dimensioni dei moscerini non selezionati (es. 1,3 mg)
B) Si selezionano i moscerini di grandi dimensioni (es. 3 mg)
C) Si concentrano in una bottiglia e si lasciare fecondare
D) Si misura la grandezza corporea media (es. 2 mg), confrontando con quelli dei
non selezionati
Se alla base della variabilità per la dimensione corporea c’è variabilità genetica, la
progenie selezionata assomiglierà ai genitori
ereditabilità in senso stretto
Risposta alla selezione =
× differenziale di selezione
(2 mg – 1,3 mg) = 0,7 mg
ereditabilità in senso stretto
× (3 mg – 1,3 mg) = 1,7 mg
ereditabilità in senso stretto
(h2
107
N)=
0,7 mg / 1,7 mg = 0,41
9.9 EREDITABILITA’ IN SENSO LARGO E STRETTO
Considerando che con la selezione possono essere fissate solo le differenze
genetiche attribuibili alle azioni geniche additive, i genetisti considerano solo la
proporzione della varianza fenotipica ascrivibile alla componente additiva della
varianza genetica
R = S h2
R = risposta alla selezione
S = differenziale di selezione
h2 = R/S
L’analisi genetica della variabilità fenotipica permette di
stimare l’incidenza relativa delle cause genetiche e ambientali
nella determinazione di un carattere quantitativo
108
9.9a RISPOSTE ALLA SELEZIONE
Gli esperimenti di selezione per un carattere quantitativo sono
fondamentalmente di tre tipi:
A) Selezione in una direzione senza alcuna popolazione di
controllo …..
109
9.9a RISPOSTE ALLA SELEZIONE
B) Selezione in una direzione con una popolazione di controllo
…..
110
9.9a RISPOSTE ALLA SELEZIONE
C) Selezione nelle due direzioni opposte (selezione divergente)
con una popolazione di controllo
111
9.9a RISPOSTE ALLA SELEZIONE
Da un punto di vista pratico, le risposte più comuni che si
ottengono con il lavoro di selezione sono 5:
1) Risposta immediata che si esaurisce rapidamente
Nota
Questa risposta per caratteri
quantitativi controllati da loci in
assenza di dominanza, quindi
al primo ciclo di selezione si
possono distinguere i genotipi
AA, Aa e aa; mentre in presenza di dominanza ci vogliono
due cicli di selezione
112
9.9a RISPOSTE ALLA SELEZIONE
2) Risposta iniziale rapida e successivamente lenta
Nota
Risposte di questo tipo
fanno pensare che il
carattere quantitativo
oggetto di selezione sia
controllato da pochi
geni con effetti vistosi
(major genes) e da
molti geni minor (che
determinano piccoli
effetti sul fenotipo)
113
9.9a RISPOSTE ALLA SELEZIONE
3) Risposta non molto evidente ma costante nel tempo
Nota
Il tipo di risposta osservata
fa pensare con tutta evidenza che il carattere oggetto di
selezione sia sotto il controllo di moltissimi geni con piccolissimi effetti genetici di
tipo additivo
114
9.9a RISPOSTE ALLA SELEZIONE
3) Risposta non molto evidente ma costante nel tempo
Nota:
Nel 1896 G.H. Hopkins, presso la Stazione Sperimentale Agraria
dell’Università dell’Illinois, iniziò uno studio volto a determinare se
la composizione chimica della cariosside di mais poteva essere
cambiata con la selezione. Questa attività è stata già condotta per
oltre 100 anni e prosegue ancora oggi …..
115
9.9a RISPOSTE ALLA SELEZIONE
3) Risposta non molto evidente ma costante nel tempo
Nota:
116
9.9a RISPOSTE ALLA SELEZIONE
4) Risposte positive alternate a risposte nulle
Nota
Questo tipo di risposta tiene
conto di un modello genetico
che tiene conto della distinzione tra variabilità libera
(AA e aa) e variabilità potenziale (Aa). Considerando
più loci esiste una larga quota di variabilità potenziale
omozigote (AAbb) che deve essere prima convertita in
variabilità potenziale
eterozigote
117
9.9a RISPOSTE ALLA SELEZIONE
5) Risposta limitata o nulla
Nota
Questo tipo di risposta si ottiene quando il carattere oggetto di selezione presenta
una scarsa variabilità genetica di tipo additivo, quindi
una bassa ereditabilità,
rendendo inefficace il lavoro
di selezione
118
9.10 GENI MODIFICATORI, PENETRANZA ED ESPRESSIVITA’
Dal punto di vista sperimentale, il modo per studiare i caratteri quantitativi e i geni
che li controllano (QTL) è basato su:
- uso di marcatori genetici
- il mappaggio sui cromosomi
- popolazioni sperimentali appositamente costituite
- procedure statistiche articolate
Marcatori genetici sono geni che controllano un carattere quantitativo con
manifestazione fenotipica localizzazione cromosomica nota
La relazione tra i geni che controllano i marcatori morfologici e quelli che
controllano i caratteri quantitativi sono limitati (es. gene che controlla il colore dei
fagioli è associato con il gene che controlla il peso del fagiolo)
119
9.10 GENI MODIFICATORI, PENETRANZA ED ESPRESSIVITA’
I caratteri quantitativi sono condizionati da geni modificatori
L’espressione di un carattere può dipendere dall’interazione con altri geni
e da fattori ambientali.
Geni “minor” sono geni che modificano l’effetto visibile di altri geni e
alterano la manifestazione fenotipica di un carattere solo quando è
presente il gene principale (“major”)
L’effetto dei geni modificatori non sempre è facilmente distinguibile.
Talvolta si confonde con l’effetto ambientale
120
9.10 GENI MODIFICATORI, PENETRANZA ED ESPRESSIVITA’
Penetranza = capacità di un allele dominante di manifestarsi
fenotipicamente
Proporzione degli individui che manifestano il carattere rispetto al totale che
portano quell’allele. Si valuta sulla popolazione e non sul singolo individuo
Penetranza incompleta = la non espressione del carattere negli individui che
portano il relativo allele
Un carattere che si manifesta nell’80% della popolazione vuol dire che una
penetranza dell’80%
121
9.10 GENI MODIFICATORI, PENETRANZA ED ESPRESSIVITA’
Espressività = intensità con cui un gene si manifesta a livello
fenotipico e può variare da elevata a nulla (in assenza di
penetranza del gene)
A livello del singolo individuo il concetto di espressività si intende in termini
quantitativi
Quando un carattere ha molta differenza nel suo grado di manifestazione vuol dire
che è ad espressività variabile
122
9.10 GENI MODIFICATORI, PENETRANZA ED ESPRESSIVITA’
Penetranza 10% anche allo stato omozigote
Condizioni ambientali => 100% - 0%
Espressività variabile nel senso della quantità delle zone interessata
Il variare della penetranza e della espressività viene spiegata con l’azione
dei geni modificatori o con l’influenza dei fattori ambientali, anche se le
123
cause sono difficili da distinguere
Quantitative Trait Loci
I geni, che controllano i caratteri quantitativi
(quantitative trait loci or QTLs), di solito non
possono essere identificati individualmente
È possibile tuttavia localizzare le regioni del
genoma in cui questi geni si trovano (linkage
mapping) e stimare quanta parte della variabilità
totale si spiega con la variabilità dei QTL in ogni
regione.
124
Quantitative Trait Loci
• Si inincrociano due linee che differiscono marcatamente
per il carattere quantitativo (alto vs basso) e, inoltre,
rispetto ad alleli a loci conosciuti (geni marcatori, RFLPs,
VNTRs) che permettono di distinguere i diversi genotipi
• La F1 prodotta si incrocia con se stessa in modo da
ottenere una F2 segregante.
• Se vi sono QTL strettamente associati con un gene
marcatore, allora nella generazione segregante i
genotipi marcatori portaranno anche gli alleli QTL.
125
Analisi del linkage
• Questa tecnica è
stata utilizzata per
localizzare segmenti
cromosomici associati
con caratteri quali il
peso dei frutti in
pomodoro e il numero
di setole in Drosophila
126