La fotosintesi
Le trasformazioni, orientate alla produzione di materia
organica, subite dall'acqua e dall'anidride carbonica sono
processi che non avvengono spontaneamente, ma hanno
bisogno di un notevole apporto di energia dall'esterno per poter
essere svolti (si parla, in questo caso, di reazioni
"endoergoniche"). La luce del Sole, catturata dai pigmenti
fotosintetici, forniscono l'energia necessaria
ad alimentare l'intera serie di reazioni chimiche.
Influenza sull’ambiente
Il fatto che la superficie della Terra (a differenza di quel che
accade negli altri pianeti del Sistema Solare) sia un luogo tanto
adatto per il manifestarsi delle innumerevoli forme di vita che ci
circondano, si deve principalmente a due cause:
1. la presenza dell'acqua allo stato liquido,
2. la presenza dell'ossigeno nell'atmosfera.
La reazione complessiva della fotosintesi può
essere così riassunta:
Anidride carbonica
x CO2 + y H2O => Cx(H2O)y + x O2
acqua
zucchero
ossigeno
FASE LUMINOSA
FASE OSCURA (avviene sempre)
HA LUOGO NEI CLOROPLASTI
• Amido:
– Farine
– Patate
– Semi di cereali e altri
• Cellulosa
– Tutte le erbe giovani e quindi anche alberi
• Zuccheri ……dolci
– Saccarosio (il solito zucchero)
– Fruttosio
– Maltosio (zucchero per fare la birra…. Si trova nell’orzo)
– Zucchero di canna……
– Galattosio: lo zucchero del latte…
I carboidrati zuccheri vengono costituiti utilizzando l'anidride
carbonica dell'aria, che viene ridotta per mezzo dell’idrogeno estratto
dall’acqua. L’energia luminosa necessaria per alimentare il
processo è catturata dal pigmento verde clorofilla, presente
nelle foglie.
L’acqua è un donatore di elettroni di atomi di idrogeno piuttosto pove
(potenziale redox di 0,82 V contro un valore di -0,82 V del NADH).
Alla base di tutta la complessa serie di reazioni che portano ad
ottenere tale equazione, troviamo la scissione dell'acqua nei
suoi componenti.
2 H2O => O2 + 4 H+ + 4 eL'ossigeno viene liberato sotto forma di gas O2 (l'ossigeno
molecolare dell'aria, appunto), mentre l'idrogeno (sotto forma di
ioni H+ ed elettroni) è poi utilizzato nelle reazioni successive
per trasformare la CO2 in carboidrati.
L'acqua è una molecola stabile, quindi poco reattiva, e il
processo della sua scissione è una reazione fortemente
endoergonica, che per avvenire ha bisogno di essere
alimentata dall'energia solare (si parla infatti di "fotolisi", ossia
"scissione per mezzo della luce").
Per mezzo della fotolisi, l’energia radiante della luce solare viene
convertita in energia chimica, dato che le molecole di idrogeno e di
ossigeno contengono una quantità maggiore di energia chimica
rispetto alla molecola d’acqua da cui sono derivate.
La clorofilla, a normali temperature e con l’energia della luce
visibile, scinde le molecole d’acqua, svolgendo un lavoro che, in
condizioni ordinarie, richiede temperature di circa 2000°C o una
forte corrente elettrica.
Proprio la fotosintesi, svolta nel corso di centinaia di
milioni di anni da piante e batteri fotosintetici, sarebbe
responsabile delle trasformazioni che hanno portato
l'atmosfera del nostro pianeta alla sua attuale
composizione.
Gli organismi fotosintetici avrebbero dunque
trasformato radicalmente la nostra atmosfera, estraendo
l'ossigeno gassoso dall'acqua e riducendo notevolmente
la proporzione di anidride carbonica (oggi vicina allo
0,03%).
Strato di ozono
La radiazione luminosa è costituita da fotoni, in grado di
colpire i pigmenti fotosintetici e di eccitare gli
elettroni ad un livello energetico più alto (teoria quantistica).
La luce però ha anche propagazione di tipo ondulatorio ed è
dotata di una lunghezza d'onda caratteristica, dalla quale
dipende la quantità di energia trasportata (teoria ondulatoria).
RAGGI GAMMA
da 10-3 a 10-6 nm
RAGGI X
da 1 a 10-3 nm
ULTRAVIOLETTO
da 103 a 1 nm
LUCE VISIBILE
circa 103 nm
INFRAROSSO
da 103 a 106 nm
MICROONDE
da 106 a 109 nm
ONDE RADIO
> 109 nm (1 m)
Lo spettro dei colori della luce visibile va dalla zona del violetto
(a circa 400 nm) a quella del rosso (a circa 750 nm).
Nell'intervallo compreso tra le zone del rosso e del viola
troviamo tutti gli altri colori, tra i quali anche quelli delle
radiazioni utilizzate dalle piante per la fotosintesi.
I fatto che la fotosintesi utilizzi la luce visibile piuttosto che le
altre zone dello spettro elettromagnetico è probabilmente legato
alla maggior abbondanza delle radiazioni luminose rispetto a tutte
le altre (la nostra atmosfera è trasparente alla luce visibile) per cui
gli organismi, come le piante, in grado di sfruttarle avrebbe
sarebbero stati favoriti da un punto di vista evolutivo.
Inoltre, le radiazioni a lunghezza d'onda più grande di quelle del
rosso (oltre 750 nm) hanno scarsa energia, che per la maggior
parte è assorbita dall'acqua come calore, mentre quelle a
lunghezza d'onda minore della luce viola (sotto i 400 nm) ne
hanno troppa e, se assorbite, degraderebbero rapidamente molte
molecole biologiche.
I pigmenti sono sostanze chimiche che appaiono colorate
perché, quando vengono illuminate dalla luce bianca, sono
in grado di assorbire solo alcune radiazioni dello spettro
luminoso, mentre si lasciano attraversare da tutte le altre.
La molecola di un pigmento che rientri tra le sostanze
organiche (categoria alla quale appartengono anche i pigmenti
fotosintetici), presenta caratteristiche chimiche ben precise.
I pigmenti organici contengono infatti (accanto a particolari
gruppi funzionali, detti "cromofori" se sono colorati di per sé, o
"auxocromi" se aumentano l'intensità del colore) e una
serie di "doppi legami coniugati“.
Un doppio legame isolato assorbe infatti radiazioni nella zona
dell'ultravioletto, ma se viene "coniugato" con altri doppi legami,
l'assorbimento si sposterà nel campo della luce visibile (e,
all'interno di questo, dalla zona del viola a quella del rosso). In
altre parole, quanto maggiore è il numero dei doppi legami
coniugati, tanto maggiore sarà la lunghezza d'onda assorbita
dal pigmento.
L'assorbimento di una radiazione luminosa da parte di un
pigmento è un fenomeno che riguarda l'attivazione di uno o più
elettroni periferici che fanno parte del sistema dei doppi legami
coniugati. Quando cioè un fotone di frequenza appropriata
(ossia un fotone che trasporta l'esatta quantità di energia per
quella certa transizione) colpisce una molecola di pigmento, gli
elettroni dei doppi legami passano dal loro normale livello
energetico (stato fondamentale) ad un livello energetico più
alto (stato energetico "eccitato").
Tre doppi legami coniugati assorbono a circa 260 nm.
La sostanza appare bianca perché tutte le frequenze del visibile
sono riflesse.
Cinque doppi legami coniugati assorbono a circa 330 nm.
La sostanza appare bianca perché tutte le frequenze del visibile
sono riflesse.
Sette doppi legami coniugati assorbono a circa 380 nm.
La sostanza appare verde perché tutte le frequenze del visibile
sono riflesse, ad eccezione di quella viola assorbita.
Nove doppi legami coniugati assorbono a circa 425 nm.
La sostanza appare giallo-verde perché tutte le frequenze del
visibile sono riflesse, ad eccezione di quella blu-viola
assorbita.
Undici doppi legami coniugati (es. carotenoidi) assorbono a
circa 450 nm. La sostanza appare giallo-arancione perché tutte
le frequenze del visibile sono riflesse, ad eccezione di quella
blu assorbita.
L'energia assorbita dal pigmento può essere poi riemessa in
modi molto diversi a seconda dei casi e dello stato eccitato
raggiunto.
1.Riemissione sotto forma di radiazione luminosa avente
minore energia e quindi lunghezza d'onda maggiore di
quella assorbita: questo fenomeno si definisce
fluorescenza. In pratica l'elettrone sale al livello eccitato
con un solo salto, mentre ridiscende allo stato fondamentale
con due o più salti (l'energia di ognuno dei quali è una
frazione di quella assorbita). E' il fenomeno con cui i
carotenoidi assorbono le radiazioni
blu-violette e riemettono le radiazioni rosse che possono
essere assorbite dalla clorofilla.
2.Riemissione lenta di radiazione luminosa. Questo
fenomeno viene definito fosforescenza.
3.Dissipazione dell'energia sotto forma di calore, che riporta
l'elettrone del pigmento allo stato fondamentale o ad uno
eccitato a minore energia;
4.Trasferimento dell'elettrone eccitato ad una molecola
accettore, dopo di che l'elettrone perduto deve essere
rimpiazzato a spese di un'altra molecola che funge da
donatore di elettroni (ad esempio, la clorofilla cede
l'elettrone eccitato ad un accettore, sostituendolo, subito
dopo, con un altro elettrone proveniente dall'acqua, nei
cloroplasti).
In termini ossido-riduttivi, l'elettrone ceduto inizialmente
(riduzione dell'accettore) viene rapidamente sostituito da un
elettrone ceduto da una seconda molecola (ossidazione del
donatore). L'intero fenomeno è appunto definito
ossido-riduzione.
Clorofille e carotenoidi
Come tutti i pigmenti, anche i pigmenti fotosintetici sono
caratterizzati da uno "spettro di assorbimento".
Lo spettro di assorbimento è una curva, rappresentata in un
diagramma che riporta in ascisse la lunghezza d'onda ed in
ordinate l'entità dell'assorbimento ("Assorbanza", indicata con
"A" e definita come il logaritmo del rapporto tra l'intensità di
luce che raggiunge il pigmento, "luce incidente"= I0, e quella
che lo attraversa senza essere assorbita, "luce trasmessa"= I).
A = log (Io / I)
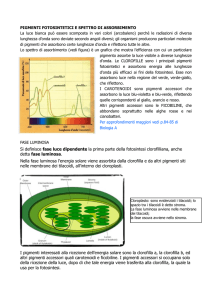
I pigmenti
Lo spettro di assorbimento ci dà la misura dell'intensità
luminosa che viene assorbita per ciascuna lunghezza d'onda,
quindi ci può fornire utili informazioni riguardanti la natura dei
pigmenti fotosintetici.
I principali pigmenti fotosintetici sono certamente le
clorofille: specialmente la clorofilla a, il pigmento che innesca le
reazioni della fase luminosa e la clorofilla b.
Esistono, tuttavia, altri tipi di pigmenti definiti “accessori” quali
i carotenoidi (β-carotene, xantofilla) e le ficobiline (ficoeritrina e
la ficocianina di cianobatteri e alghe rosse).
L'efficacia relativa delle varie lunghezze d'onda sulla prima
fase della fotosintesi è deducibile dallo "spettro d'azione
fotosintetico", un diagramma simile allo
spettro d'assorbimento, tranne che per il fatto di riportare in
ordinate la velocità della fotosintesi, anziché il semplice
assorbimento di luce.
Lo spettro d'azione fotosintetico mostra un andamento
parallelo allo spettro d'assorbimento della clorofilla a, con
piccole differenze (le bande di frequenze efficaci nel
promuovere la fotosintesi sono più ampie di quelle assorbite
dalla clorofilla) dovute alla presenza dei pigmenti accessori.
Questi sono, infatti, in grado di assorbire luce in zone diverse
dello spettro e di trasferire, poi, la loro energia di attivazione
alla clorofilla a.
L'immagine mostra lo spettro d'assorbimento della clorofilla a (in
alto) confrontato con lo spettro d'azione della fotosintesi (in
basso).Come si può osservare, i due spettri hanno un andamento
parallelo in molte zone (le differenze sono dovute all'influenza dei
pigmenti accessori sulla velocità della fotosintesi).
La clorofilla
La molecola della clorofilla a è
caratterizzata da un "nucleo
porfirinico" formato da
quattro anelli pirrolici, un atomo
di magnesio (Mg) e numerosi
doppi legami coniugati.
E' la parte evidenziata in
verde, in quanto è responsabile
dell'assorbimento di energia
luminosa e, quindi, della
colorazione verde della
clorofilla stessa.
Può essere paragonata a una
vera e propria antenna
ricevente.
.
Oltre al "nucleo porfirinico", la
molecola presenta una lunga
catena idrocarburica, detta
"fitolo".
Questa può essere
paragonata in qualche modo
all'asta che sostiene l'antenna.
La catena del fitolo è infatti la
porzione della molecola che
permette l'ancoraggio della
clorofilla allo strato lipidico
della membrana dei tilacoidi
I carotenoidi
La molecola del β-carotene è
caratterizzata da ben undici
doppi legami coniugati, che le
conferiscono la ben nota
colorazione giallo-arancione (è
la parte evidenziata, appunto,
in arancione).
I cloroplasti sono in grado di
assorbire una banda di
radiazioni molto ampia nella
zona del viola-blu-azzurro, per
cui riescono ad utilizzare
anche le frequenze non
assorbite dalla clorofilla.
Il loro ancoraggio, nella
membrana dei tilacoidi, è simile
a quello del fitolo.
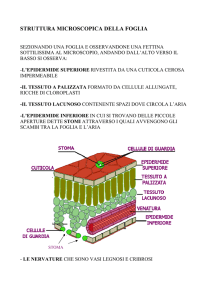
La foglia
Le uniche cellule
dell'epidermide ricche
di cloroplasti (e quindi
in grado di svolgere la
fotosintesi) sono
proprio le "cellule di
guardia" degli stomi.
Tutte le altre cellule
epidermiche sono
trasparenti alla luce,
che le attraversa per
andare a colpire i
tessuti sottostanti, i
clorenchimi,
innescando la prima
fase della
fotosintesi.
Gli stomi
I cloroplasti
Cloroplasti in una foglia di muschio
Doppia membrana esterna
Serie di membrane interne sovrapposte e
collegate fra loro in modo da ottenere il
massimo sviluppo di area superficiale: sono
le membrane fotosintetiche, quelle in cui si
trovano "ancorati" i pigmenti destinati a
catturare la luce solare.
Vescicole, o sacchetti (tilacoidi), alcuni dei
quali impilati gli uni sugli altri in strutture
simili ad ammassi (grana) I grana sono
collegati tra loro da altre membrane con
struttura lamellare (intergrana).
I tilacoidi sono immersi in una soluzione
(stroma),
caratterizzata da un alto contenuto di
proteine (specialmente
enzimi, tra i quali quelli destinati a formare
i carboidrati).
E' stato ipotizzato che tutti i tilacoidi siano, in qualche modo,
collegati tra di loro, in modo da formare un’unica cavità interna.
Questo spazio interno servirebbe (come vedremo, in seguito)
da sito di accumulo per gli ioni H+ liberati nella prima fase della
fotosintesi (il gradiente formato dai quali, ai due lati della
membrana dei tilacoidi, è assai importante per fornire energia
alla sintesi chemiosmotica di ATP).
I cloroplasti, nelle piante superiori, sono piccoli e numerosi ed hanno
sempre forma lenticolare.
Nelle alghe verdi, invece, i cloroplasti sono pochi e di grandi
dimensioni. La loro forma è molto variabile e caratteristica per
ciascuna specie.
All'interno di questi cloroplasti sono ben distinguibili, anche al
microscopio ottico, uno o più corpi tondeggianti detti
"pirenoidi", che sono i siti di accumulo dell'amido primario
appena formato.
In effetti, rispetto alla membrana che ricopre il cloroplasto, la
membrana dei tilacoidi presenta diverse peculiarità. In
particolare, nella membrana tilacoidale troviamo tre gruppi di
molecole importantissime.
1.I fotosistemi, di cui il PS II presente soprattutto sulla
membrana dei tilacoidi impilati in grana e il PS I più esterno per
essere accessibile al NADP+ nello stroma.
Per "fotosistema" si intende l'insieme dei pigmenti
fotosintetici, che comprende i pigmenti antenna
(accessori) e i pigmenti del centro di reazione (tra cui la
cosiddetta "coppia speciale" di molecole di clorofilla a, che
fa partire le reazioni della fase luminosa).
L'ampio sviluppo superficiale di cui è dotata, il rapporto al
piccolo volume racchiuso nello spazio interno dei tilacoidi,
permette alla membrana di sfruttare al meglio la luce
ricevuta, creando rapidamente un'alta concentrazione di
ioni H+.
2.Il sistema di trasporto elettronico del cloroplasto, in grado
di ricevere gli elettroni perduti dalla clorofilla, in seguito
all'assorbimento di luce.
3.I complessi proteici dell'ATP-sintetasi, in grado di sfruttare
il gradiente di ioni H+, creatosi tra lo spazio interno dei
tilacoidi e lo stroma, per produrre molecole di ATP con
meccanismo chemiosmotico.
Nella fase luminosa i pigmenti fotosintetici assorbono
l'energia radiante del sole e la trasformano in energia
chimica (sotto forma di legami fosfato nelle molecole di
ATP e come potere riducente nel NADPH). In questa fase
viene utilizzato l'idrogeno dell'acqua e rilasciato O2 come
sottoprodotto.
Nella fase oscura l'ATP e il NADPH formati nella prima
fase riducono l'anidride carbonica, utilizzandola per
sintetizzare i carboidrati.
Lo schema seguente mostra le due semireazioni
(l'ossidazione dell'ossigeno dell'acqua e la riduzione
dell'anidride carbonica) in modo separato
2 H2O
===>
NADP+ + 3 ADP +3 Pi <=====>
[CH2O]
=====>
O2
2 NADPH + H+ +3 ATP
CO2
La cattura dell’energia luminosa
I fotoni che, attraversando gli strati
dell'epidermide ed entrando
nel cloroplasto, raggiungono la membrana dei
tilacoidi, e colpiscono i fotosistemi.
Ciascun fotosistema è composto da due
subunità complementari: il gruppo dei
pigmenti antenna ed il centro di reazione
fotochimico. In quest’ultimo avviene la
separazione di carica che dà inizio a una
serie di reazioni redox.
I fotosistemi
PS I con più chl A assorbe a
700 nm e il suo centro di
reazione prende il nome di
P700
PS II con chl A = chl B assorbe
a 680 nm e il suo centro di
reazione prende il nome di
P680 (fotosintesi ossigenica da
3 mld di anni).
In questo esempio la radiazione riemessa è identica a quella assorbita
In questo esempio vengono riemesse più radiazioni,
La somma delle cui energie è identica all'energia della radiazione assorbita.
Nel caso della clorofilla a in un centro di reazione, l'elettrone
"ridiscende" (in termini energetici) in più tappe, ma anziché
attraversare i livelli energetici della molecola di partenza fino
allo stato fondamentale, fa la sua discesa abbandonando la
clorofilla e trasferendosi agli orbitali di molecole vicine, posti a
livelli energetici via via più bassi.
In altri termini, l'elettrone della clorofilla, una volta raggiunto il
suo stato eccitato viene perduto dalla clorofilla stessa (che
resta carica positivamente) e ceduto ad un accettore di
elettroni che si trova nello stesso complesso del centro di
reazione, da dove, successivamente, continuerà la sua
"discesa energetica" passando alle altre molecole del sistema
di trasporto elettronico.
Le reazioni della fase luminosa
1) IL TRASPORTO DI ELETTRONI
L’energia luminosa, alimentando una reazione redox
endoergonica, viene trasformata in energia elettrica in forma di
cariche elettriche separate (cioè di differenza di potenziale):
avendo perso un elettrone, la clorofilla rimane carica
positivamente e la molecola di accettore che ha acquisito
l’elettrone porta una carica negativa.
Le reazioni del trasferimento elettronico possono essere
considerate come una serie di ossidoriduzioni, in cui ogni
molecola si comporta, di volta in volta, da accettore e donatore di
elettroni (riducendosi e acquistando energia e, poi, ossidandosi di
nuovo e tornando alla sua energia di partenza).
La figura rappresenta il cosiddetto "schema Z", che prende il
nome dal modo in cui viene generalmente rappresentata la
traccia lasciata dal movimento degli elettroni e che riassume i
passaggi fondamentali della fase luminosa della fotosintesi.
Come si vede lo schema è diviso in due segmenti, uno per
ogni fotosistema:
• il primo segmento (quello alimentato dal fotosistema II)
riguarda la fotolisi dell'acqua e la produzione di ATP (e
viene perciò detto "segmento ATP"),
• il secondo (alimentato dal fotosistema I) riguarda il destino
finale degli elettroni e la produzione di NADPH.
Feofitina (no Mg2+)
2 H2O O2 + 4 H+ + 4 e(sono richiesti 4 fotoni)
Fotosistema II
P680 + 1 hν P680*
P680* + Ph (feofitina) P680+ + Ph2 Ph- + QA 2 Ph + QA2QA2- + 2 H+ (dallo stroma) QBH2
---------------------------4 P680 + 4 H+ + 2 QB + 4 hν 4 P680+ + 2 QBH2
Complesso che scinde l’acqua
4 P680+ + 4 Tyr (pro D1 - PSII) 4 P680 + 4 Tyr+
4 Tyr+ + [complesso Mn]0 4 Tyr + [complesso Mn]4+
[complesso Mn]4+ + 2H2O [complesso Mn]0 + 4 H+ + O2
---------------------------2 H2O + 4 fotoni 4 H+ (nel lume) + 4 e- (al PSII) + O2 (gas)
TOTALE = 2 H2O + 2QB + 4 hv O2 + 2 QBH2
Trasferimento dal PS II al PS I
QBH2 (mobile) cyt bf (cyt b563 2 eme, FeS, cyt c552) plastocianina (Cu+/++)
Fotosistema I
P700* Ao A1 (fillochinone) Fe-S Fd (ferredossina 2Fe-2S)
Ferredossina-NADP+ ossidoreduttasi (Fp)
2Fd2+red + 2H+ + NADP+ (nello stroma) 2 Fd3+ox + NADPH + H+
Reazione totale della fase luminosa
2 H2O + 2 NADP+ + 8 hv O2 + 2 NADPH + 2 H+
4 fotoni assorbiti in in ogni PS
6 H+ nel lume (2 dallo stroma e 4 dall’acqua)
pH 8 nello stroma e pH 4,5 nel lume
Riassumendo, il flusso degli elettroni nelle membrane dei tilacoidi
ha convertito l’energia luminosa in due forme:
1. Il potenziale ridotto del NADPH, che sarà successivamente usato
nella fissazione del carbonio.
2. Il gradiente di H+ attraverso la membrana dei tilacoidi. La
generazione di questa riserva di H+ e la sua utilizzazione per
alimentare la sintesi chemiosmotica di ATP, insieme vengono
chiamate fosforilazione fotosintetica (o fotofosforilazione).
Gli elettroni a volte seguono un diverso itinerario di trasporto
elettronico. Questa via è chiamata flusso di elettroni ciclico
perché la clorofilla P700, nel fotosistema I, funge
contemporaneamente da donatore e da accettore finale di
elettroni (a colori, nella figura seguente).
Ciò si verifica quando la maggior parte del NADP+ è stato
ridotto, e ne rimangono poche molecole disponibili ad
accettare elettroni. Gli elettroni allora cedono la loro energia al
segmento-ATP del sistema di trasporto elettronico, dove
vanno ad incrementare la riserva di H+ nello spazio interno dei
tilacoidi (con produzione di ulteriori molecole di ATP).
ADP + Pi + (luce) ATP + H2O
Le reazioni della fase luminosa
2) LA SINTESI CHEMIOSMOTICA DI ATP
Man mano che il trasporto degli elettroni procede, la
concentrazione di H+ nel lume interno dei tilacoidi va sempre
più aumentando, mentre nello stroma rimane bassa. Si crea
così un notevole gradiente di concentrazione (la
concentrazione degli ioni idrogeno all'interno dei tilacoidi arriva
ad essere anche 3000 volte superiore a quella dello stroma)
associato ad un'energia potenziale che è in grado di compiere
un lavoro.
La membrana dei tilacoidi è impenetrabile a questi ioni,
tranne che in corrispondenza di particolari canali proteici
associati ai complessi dell'ATP-sintetasi: la rapida diffusione di
H+ nello stroma cede l'energia potenziale associata al
gradiente all'ATP-sintetasi, che la utilizza immediatamente per
La sintesi di nuove molecole di ATP.
Le frecce in rosso indicano il passaggio di protoni, quelle in blu il passaggio di
elettroni
Il meccanismo con il quale la fosforilazione di ATP è accoppiata
alla diffusione di ioni idrogeno secondo il gradiente di
concentrazione è detto chemiosmosi (o fotofosforilazione).
Un meccanismo simile si incontra anche nei processi respiratori
che avvengono nei mitocondri.
Complesso CF0CF1 = ATP sintetasi
CF0 (transmembrana) = 2 a, b, b’, 6-12 c
CF1 (periferica) = 3 a, 3 b, g, d, e
Stechiometria ancora incerta, ma
1 o 2 ATP per coppia di protoni
Resa della fotosintesi
6 CO2 + 6 H2O + (luce) C6H12O6 + 6 O2
DE libera standard = 2840 kJ/mole
Nella fotosintesi sono spesi, per mole di
glucosio prodotta, da 8160 a 14400 kJ (a
seconda della lunghezza d’onda dei fotoni)
Resa = dal 20% al 34%
Le reazioni della fase oscura
LA FISSAZIONE DEL CARBONIO
Granuli di amido secondario di fagiolo
Nelle reazioni della fissazione del carbonio (definita anche, a
volte, come organicazione del carbonio) l'anidride carbonica
viene legata ad una preesistente molecola di carboidrato e
ridotta a formare un nuovo carboidrato (con un atomo di
carbonio in più), grazie all'energia dall'ATP e l'idrogeno dal
NADPH, entrambi prodotti dalle reazioni della cattura
energetica.
Si dice che il carbonio viene "fissato", perché si ha
l'incorporazione di un gas (la CO2) in una molecola "fissa",
solida. Si parla invece di organicazione perché l'anidride
carbonica, che è inorganica, viene trasformata nella materia
organica dei carboidrati.
Gli enzimi che catalizzano queste reazioni si trovano nello
stroma dei cloroplasti.
CICLO DI CALVIN-BENSON (1940-1950)
o CICLO C-3
Che bisogno hanno di arrivare fino alla formazione dei carboidrati?
I motivi sono, fondamentalmente, due.
1. ATP e NADPH hanno entrambi una vita breve. Il
loro stesso alto contenuto energetico le rende
estremamente instabili. Le molecole di ATP sono
state giustamente paragonate alle monete in circolazione:
un discorso analogo può essere fatto per il NADPH.
Entrambe le molecole devono essere "spese"
rapidamente, subito dopo essere state "guadagnate"
(con la differenza che il NADPH può essere "speso"
solo nelle reazioni di riduzione).
2.
I carboidrati accumulati, oltre ad essere molecole
stabili (che possono durare nel tempo o, secondo
le necessità, essere rapidamente convertiti in
energia) costituiscono gli scheletri carboniosi di
base su cui la pianta (attraverso opportune
modifihe) costruirà tutte le molecole organiche
necessarie al suo metabolismo.
La fissazione del carbonio inizia con l'attacco dell'anidride
carbonica sul ribulosio bifosfato. La risultante struttura a
sei atomi di carbonio viene immediatamente idrolizzata a due
molecole di fosfoglicerato (PGA) a tre atomi di carbonio
ciascuna.
3
a) Fissazione
+ 3 ATP
Ribulosio-1,5-bisfosfato carbossilasi
3
3 Ribulosio-5-fosfato
Ribulosio-1,5-bisfosfato carbossilasi
6
2 Pi
1,3-fosfoglicerato chinasi
6
b) Riduzione
(isoenzimi glicolitici che
funzionano con il NADPH)
c) Rigenerazione
dell’accettore
6
Gliceraldeide-3-fosfato deidrogenasi
6
6 Pi
5
1 (netto)
6
1) GLICOLISI
2) SINTESI ESOSI
E AMIDO
RUBISCO
L'enzima che catalizza questa reazione è la
ribulosio bifosfato carbossilasi (Rubisco).
Dal momento che le reazioni catalizzate dalla Rubisco
rimangono ancora piuttosto lente, le piante producono
quantità enormi di questo enzima, che, da solo, raggiunge
circa il 25% di tutto il materiale proteico presente nei cloroplasti
ed i 50% di quello dello stroma.
Di conseguenza, è stato osservato
che la Rubisco è la più abbondante
proteina presente sulla Terra!
M= 660000. Nello STROMA
8 subunità grandi (sito attivo) = 56000
8 subunità piccole (???) = 14000
I composti intermedi della reazione della Rubisco
IL DESTINO DEL
3-FOSFOGLICERATO…
• Rigenerazione del ribulosio-1,5bisfosfato.
• Si combina con il didrossiacetone
fosfato (aldolasi e fruttosio-1,6bisfosfatasi) a formare fruttosio-6fosfato e glucosio-1-fosfato, un
precursore dell’amido.
• Convertito in diidrossiacetone fosfato
(trioso fosfato isomerasi), poi trasportato
dal cloroplasto nel citosol (antiporto Ptrioso fosfato), e infine degradato nella
glicolisi (sintesi acidi grassi e
aminoacidi) o utilizzato per la sintesi di
fruttosio-6-fosfato e quindi di saccarosio
(linfa elaborata) o per il trasporto netto
di NADPH/NADH e ATP nel citosol.
• Convertito in cellulosa o in altri
costituenti saccaridici della parete
cellulare.
Trioso
fosfato
isomer
asi
Trioso
fosfato
isomerasi
Aldolasi
Via inversa della via ossidativa
del pentosio-fosfato, che
converte gli esosi fosfato in
pentosi fosfato e viceversa.
Fruttosio-1,6bisfosfatasi
Transche
tolasi
Aldolasi
Sedoeptulosio
-1,7-bisfosfatasi
Transchetolasi
Ribulosio-5-fosfato chinasi
RIGENERAZIONE DEL
RIBULOSIO-1,5
BISFOSFATO
La transchetolasi trasferisce un
gruppo a due atomi di C di un
chetosio donatore al gruppo
prostetico TPP dell’enzima e poi ad
un aldosio accettore. La TPP lega un
gruppo chetolico CH2OH-CO-
3 RuBP + 3 CO2 + 3 H2O + 6 NADPH + 6 H+ + 9 ATP ==>
3 RuBP + 6 NADP+ + 8 Pi + 9 ADP + 1 gliceraldeide-3-P
Per rigenerare 9 ATP (con soli 8 Pi) c’è bisogno di importare dal citosol
nello stroma un gruppo fosfato (ANTIPORTO Pi-trioso fosfato
(DHAP)) sulla membrana interna dei cloroplasti, impermeabile agli altri
composti.
L'ADP, il Pi e il NADP+ ottenuti dal ciclo C3 sono di nuovo disponibili
per ricevere energia nelle reazioni della fase luminosa e vengono quindi
riciclati per formare nuovi ATP e NADPH.
Per la mancanza di Rubisco e ribulosio-5-P-chinasi gli animali non
possono convertire la CO2 in glucosio.
Piante contro Animali
Le piante hanno sequenze di reazioni uniche per ridurre la CO2 a
triosi fosfato, associate anche alla via riduttiva del pentosio fosfato.
Negli animali invece, la sintesi di carboidrati necessita sempre di
precursori con almeno tre atomi di C e con uno stato di ossidazione
più basso della CO2.
Le piante possono utilizzare CO2 come unica fonte di carbonio
(autotrofi) per la biosintesi non solo di carboidrati, ma anche di lipidi
e proteine. Negli animali invece, non possono ridurre la CO2 per
formare glucosio e le poche reazioni di fissazione diretta della CO2
vedono la CO2 immediatamente persa nelle reazioni succesive
(piruvato carbossilasi nella gluconeogenesi con CO2 fissato
nell’OAA; acetil-CoA carbossilasi nella sintesi degli AG; carbamil
fosfato sintetasi I nel ciclo dell’urea).
Regolazione della fotosintesi
Quando la [CO2] è troppo elevata, il gruppo ε-NH2 di una Lys della rubisco si lega
spontaneamente a CO2 (non quella fissata) e il carbamato che si forma lega uno ione
Mg2+ (necessario per la catalisi). L’enzima viene così attivato.
A [CO2] ambientali, il blocco della rubisco è superato grazie alla carbamilazione ad
opera della rubisco attivasi, a spese di 1 ATP.
La rubisco è inibita dal 2-carbossiarabinitolo-1-fosfato, un analogo del β-chetoacido
intermedio della reazione della rubisco. Questo composto è anche chiamato “inibitore
notturno” perché sintetizzato in alcune piante durante periodi di buio per ridurre la
fissazione della CO2.
• Alcuni enzimi stromali (rubisco
attivasi, fruttosio-1,6bisfosfatasi) necessitano di
ambienti alcalini e con la
presenza di Mg2+, due
condizioni che si verificano solo
in presenza di luce, quando H+ è
importato nei tilacoidi e Mg2+
esportato nel citosol.
• Ribulosio-5-P chinasi, fruttosio1,6-bisfosfatasi e la
sedoeptulosio-1,7-bisfosfatasi
sono regolati dalla condizione
dei loro residui Cys (ossidati
inattivi, ridotti attivi). PS I
ferredoxina tioredoxina (S-S
2 –SH) enzimi regolabili
(S-S 2 –SH).
“Alternative” alla clorofilla
BATTERI CHEMIOSINTETICI
scoperti nel 1880, ricavano energia senza l’uso della clorofilla:
assorbono andride carbonica al buio e non liberano ossigeno.
Ricavano energia ossidando composti di zolfo o di ferro.
BATTERIOCLOROFILLE
composti simili alla clorofilla, presenti in alcuni batteri, che
permettono di di convertire la CO2 in composti organici sfruttando
l’energia luminosa e a volte anche l’infrarosso, cosa che la normale
clorofilla non riesce a fare. Tuttavia le batterioclorofille non riescono
a catalizzare la fotolisi e quindi a favorire l’accumulo della grande
quantità di energia prodotta da questa reazione.
Conclusioni
Tutti i metodi per ricavare energia da fonti primarie, escluso quello
che utilizza la luce solare tramite la clorofilla, esistono solo in rare
e particolari condizioni e sono essenzialmente un “binario morto”,
in quanto nessun organismo più evoluto di un batterio ha utilizzato
questi metodi con successo.
Per quasi tutte le forme di vita, la clorofilla e la
fotosintesi, direttamente o indirettamente, sono la base
della vita.