1
Le membrane cellulari sono componenti importanti di tutte le cellule. Ogni cellula è circondata
da una membrana plasmatica che separa il citoplasma dall’ambiente extracellulare. La
membrana plasmatica funge da barriera di permeabilità che consente alle cellule di mantenere
una composizione citoplasmatica assai diversa da quella dei liquidi extracellulari.
Chiaramente nelle cellule degli eucarioti le membrane racchiudono anche diversi organelli.
Queste membrane dividono le cellule in compartimenti separati all’interno dei quali si svolgono
particolari processi biochimici. Molti processi cellulari di vitale importanza hanno luogo
all’interno o sulla superficie delle membrane degli organelli, esempi noti sono i meccanismi di
trasporto di elettroni e la fosforilazione ossidativa che avvengono alla superficie, all’interno e
nello spessore della membrana mitocondriale interna.
2
Il corpo umano è costituito da miliardi di cellule che devono collaborare e comunicare l’una con
l’altra pur mantenendo distinta la loro identità. La membrana cellulare gioca un ruolo chiave in
questo processo poiché costituisce la barriera che separa il compartimento intracellulare dai
segnali provenienti dall’esterno.
Ogni cellula, infatti, è circondata da una membrana plasmatica che separa il citoplasma dal
liquido extracellulare. La membrana plasmatica funge da barriera di permeabilità che consente
alle cellule di mantenere una composizione citoplasmatica assai diversa da quella dei liquidi
extracellulari. Contiene enzimi, recettori e antigeni che svolgono un ruolo fondamentale
nell’interazione con le altre cellule, con ormoni e con altre sostanze regolatrici presenti nel
liquido extracellulare.
FUNZIONE CONTENITIVA racchiude la cellula, ne definisce i confini, mantiene le differenze
tra l’ambiente intracellulare e quello extracellulare
FUNZIONE BIOCHIMICA offre sulla sua superficie interna ed esterna siti di supporto per
enzimi che così possono lavorare in modo ordinato e sequenziale
FUNZIONE FISIOLOGICA DI BARRIERA SELETTIVA seleziona in vario modo i diversi
materiali in transito tra esterno ed interno e viceversa
FUNZIONE FISIOLOGICA DI COMUNICAZIONE presenta sulla sua superficie esterna
recettori per gli ormoni e per qualsiasi altro segnale chimico, oltre che marcatori per farsi
riconoscere dalle cellule vicine
FUNZIONE STRUTTURALE alcune proteine della membrana cellulare sono ancorate a
proteine del citoscheletro che mantiene la forma della cellula
3
Le membrane cellulari e tutte le membrane presenti all’interno delle cellule sono costituite da
una combinazione di lipidi e proteine con una piccola quantità di carboidrati.
Il rapporto tra lipidi e proteine varia largamente e dipende dall’origina della membrana.
Generalmente tanto più la membrana cellulare è metabolicamente attiva tanto maggiore è il
suo contenuto di proteine.
Per esempio la membrana mitocondriale interna, che contiene enzimi per la fosforilazione
ossidativa, è costituita per ¾ da proteine.
4
La struttura delle mp è stata argomento di enorme interesse sin dal momento in cui sono state
osservate per la prima volta con un microscopio. Già dalla fine del 1800 si era giunti alla
conclusione che la membrana più esterna delle cellule fosse costituita da uno strato sottile di
lipidi che agiva da barriera tra l’interno cellulare e l’ambiente esterno. Infatti nel 1899 Overton
aveva constatato che la velocità di penetrazione all’interno della cellula delle varie molecole
che andava saggiando aumentava all’aumentare del coefficiente di ripartizione olio/acqua: ciò
indicava che la superficie cellulare doveva essere protetta da una barriera idrofobica. Inoltre
successivamente si osservò che attaccando la cellula con solventi dei lipidi o con lecitinasi
(che idrolizzano i fosfolipidi) o con complessanti del colesterolo come la digitonina si aveva
come risultato la distruzione della cellula.
Però questa struttura semplice e uniforme non è tuttavia sufficiente per spiegare le proprietà
molto variabili delle membrane presenti nei vari tipi cellulari. Come mai l’acqua è capace di
attraversare la membrana cellulare dei GR ma non in alcune cellule del tubulo renale? La
spiegazione doveva risiedere nella struttura della membrana cellulare.
Per isolare e analizzare le membrane cellulari i ricercatori del primo decennio del ventesimo
secolo fecero omogenati di cellule. Scoprirono che le membrane cellulari e tutte le membrane
presenti all’interno delle cellule sono costituite da una combinazione di lipidi e proteine con una
piccola quantità di carboidrati. Il rapporto tra lipidi e proteine varia largamente e dipende
dall’origine della membrana. Generalmente tanto più la membrana cellulare è
metabolicamente attiva tanto maggiore è il suo contenuto di proteine. Per esempio la
membrana mitocondriale interna, che contiene enzimi per la fosforilazione ossidativa, è
costituita per ¾ da proteine. L’analisi della composizione chimica delle membrane non spiega
ancora come i lipidi e le proteine siano strutturalmente disposti in una membrana. Studi
compiuti negli anni 1920 hanno suggerito che in una data area di membrana c’erano
abbastanza lipidi da formare un doppio strato
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
Le molecole lipidiche che mostrano la > asimmetria di distribuzione sono i glicolipidi. Essi si
trovano nel foglietto esoplasmatico e si associano tra loro in microaggregati tramite legami
idrogeno. I glicolipidi costituiscono il 5 % dello monostrato lipidico esterno; quelli più complessi,
gangliosidi, contengono oligosaccaridi con uno o più residui di acido sialico (componente più
comune, acido N-acetilneuramminico (NANA), e sono più abbondanti nelle membrane
delle cellule nervose dove costituiscono sino al 10% della massa lipidica totale. Sono stati
identificati sino ad ora 40 gangliosidi diversi.
Ruolo fisiologico:
Data la loro localizzazione verso l’ambiente extracellulare essi possono intervenire nella:
protezione della membrana contro condizioni estreme; basso pH; enzimi degradativi; e ad
esempio protezione delle membrane dal congelamento; i glicolipidi carichi possono alterare il
campo elettrico e di conseguenza la concentrazione esterna di ioni come il calcio; possono
avere un ruolo nell’isolamento elettrico data la loro abbondanza nella membrana mielinica;
possono avere un ruolo nei processi di riconoscimento cellulare: ad esempio il ganglioside
GM1 agisce come recettore per la tossina colerica che si lega solo alle cellule che hanno GM1
sulla superficie cellulare. Possono avere funzione di recettori per normali molecole
extracellulari ed infine prendere contatto con la matrice extracellulare
22
Le interazioni idrofobiche tra le catene carboniose di glicolipidi e fosfolipidi creano una lamina
contenente i due strati di fosfolipidi le cui teste polari sono rivolte verso il mezzo acquoso
circostante e le catene degli acidi grassi formano uno strato continuo idrofobico interno dallo
spessore di circa 4 nm. Ogni strato fosfolipidico di questa struttura lamellare prende il nome di
foglietto. Sfingomieline, galattolipidi e tutti i fosfolipidi che costituiiscono i doppi strati
contengono 2 catene di acidi grassi di lunghezza media di 16 – 18 atomi di carbonio. Le
catene aciliche insature hanno di solito un solo doppio legame ma possono averne 2, 3, 4 o
additittura 6. Di solito i doppi legami si trovano in posizione cis e questo produce una
considerevole torsione della catena alifatica che come vedremo può avere degli effetti
considerevoli sulla fluidità della membrana. Un’altra importante differenza tra i fosfolipidi
riguarda le teste polari, che a pH neutro non possiedono carica elettrica fatta eccezione per la
fosfatidilserina la cui testa polare è negativa. Sfingomielina e glicolipidi pur essendo
chimicamente diversi dai fosfolipidi hanno una struttura simile e formano con loro doppi strati
misti.
23
24
25
Oltre alla temperatura, altri fattori possono determinare la fluidità del doppio strato lipidico. In
generale i lipidi con catene aciliche corte raggiungono lo stato di transizione a temperature più
basse di lipidi con catene lunghe. Le catene corte infatti presentano una superficie inferiore per
formare interazioni di van der Waals con altre catene vicine e ciò aumenta lo stato di fluidità
della membrana. Un aumento di fluidità è determinato anche dall’aumento delle insaturazioni
degli acidi grassi. Questo è il risultato di un aumento del volume occupato da ogni singola coda
idrofobica che si ripercuote su un minore grado di impaccamento dei fosfolipidi
La presenza di insaturazioni (doppi legami) di tipo cis, aumenta il volume libero della catena di
acido grasso, aumentandone la fluidità, ancora una volta per riduzione delle forze di van der
Waals. L’aumento dell’insaturazione degli acidi grassi è una strategia che cellule batteriche o
organismi ectotermi, la cui temperatura fluttua in relazione a quella ambientale, mettono in atto
per mantenere un adeguata fluidità del doppio strato lipidico di membrana anche a bassa
temperatura. Organismi ectotermi come i teleostei Antartici adattati evolutivamente alla
sopravvivenza a temperature inferiori agli 0°C, presentano tale adattamento omeoviscoso del
doppio strato lipidico per evitare l’irrigidimento delle membrane cellulari. Un altro elemento che
influenza la fluidità del doppio strato è il colesterolo. Questo lipide è troppo idrofobico per
costituire una struttura a foglietto, ma si intercala tra i fosfolipidi. Il suo gruppo idrossilico polare
è in contatto con la soluzione acquosa vicino alle teste polari dei fosfolipidi; gli anelli steroidei
interagisce con le catene aciliche. Il colesterolo limita il movimento casuale della regione
fosfolipidica che si trova sulla superficie esterna dei foglietti, ma separa e allontana tra loro le
code delle catene aciliche inducendo una maggiore fluidità nelle regioni più interne del doppio
strato. A temperature di 37 °C il colesterolo rende le membrane complessivamente meno
fluide. A basse temperature il colesterolo ha un effetto opposto in quanto impedendo le
interazioni tra catene di acidi grassi adiacenti mantiene la membrana allo stato fluido ed
impedisce la cristallizzazione.
26
Infine anche una maggiore quantità di proteine ha generalmente una diminuzione della fluidità
delle membrane cellulari
26
27
Come si può osservare in figura la struttura dei diversi fosfolipidi è abbastanza variegata. A pH
neutro solo la fosfatidil-serina ha una carica negativa mentre gli altri fosfolipidi sono
elettricamente neutri a pH fisiologico. Ci si può chiedere allora quali sono le ragioni per le
quali le membrane delle cellule eucariotiche contengano una tale varietà di fosfolipidi con teste
polari che differiscono per dimensioni, carica e forma. Le ragioni sono da ricercare nella
funzione dei lipidi di agire come solvente bidimensionale per le proteine di membrana così
come l’acqua è un solvente tridimensionale per le proteine. Alcune proteine di membrana
possono funzionare solo in presenza di specifiche teste di fosfolipidi, proprio come molti enzimi
in soluzione acquosa richiedono per la loro attività uno ione particolare.
Oltre al fatto che i vari fosfolipidi sono diversamente rappresentati nelle membrane cellulari,
esistono enormi differenze tre i due foglietti del doppio strato differiscono in termini di
composizione fosfolipidica. In particolare i glicolipidi, la sfingomielina e la maggior parte della
fosfatidilcolina, con le loro teste cariche positivamente sono localizzate sul foglietto
esoplasmatico. I lipidi con teste polari cariche negativamente o neutre, come la
fosfatidiletanolammina e la fosfatidilserina sono localizzati prevalentemente sul foglietto
citoplasmatico. I glicolipidi sono assolutamente asimmetrici in quanto sono localizzati sul
foglietto esoplasmatico della membrana plasmatica ed in quello luminale del reticolo
endoplasmatico. Il colesterolo è distribuito uniformemente tra i due foglietti.
28
29
Da quanto detto la composizione del doppio strato lipidico può influenzare enormemente le
caratteristiche fisiche delle membrane. Mediante tecniche di frazionamento cellulare e analisi
chimiche come la gas-cromatografia e la cromatografia su strato sottile è stato possibile
evidenziare che le membrane cellulari hanno differente composizione. Tutte le membrane,
indipendentemente dalla loro origine contengono proteine e lipidi. Il rapporto proteine-lipidi può
variare notevolmente; ad esempio la le membrane interne dei mitocondri sono costituite per il
76 % da proteine mentre la mielina ne ha solo il 18%, poiché ha funzione di isolare
elettricamente la cellula nervosa dal suo ambiente. Anche la composizione lipidica è molto
variabile. Tutte le membrane contengono una certa quantità di fosfolipidi. Alcuni fosfolipidi
come gli inositolo-fosfolipidi sono presenti in quantità minori rispetto a fosfatidil-colina e
fostatidil-etanolammina ma sono funzionalmente importanti ad esempio nella trasduzione del
segnale La sfingomielina, derivato dall’ammino alcool sfingosina è anfipatica come i fosfolipidi
ed è contenuto nelle membrane plasmatiche in alta percentuale. Le membrane delle cellule
eucariotiche contengono colesterolo in percentuale molto elevata (30-50%), mentre questo
lipide è praticamente assente nella maggior parte dei batteri. Alcuni lipidi come i glicolipidi
sono presenti in un gruppo ristretto di membrane, ad esempio nella mielina. Il fosfolipide
cardiolipina è abbondantemente presente nella membrana interna mitocondriale. I carboidrati
rappresentano un componente importante di molte membrane. Essi sono legati sia alle
proteine per formare glicoproteine che ai lipidi per formare glicolpidi, costituenti molto
rappresentati nelle membrane plasmatiche delle cellule eucariotiche mentre sono assenti nelle
membrane mitocondriali interne. Il legame con i carboidrati aumenta il carattere idrofilico di
lipidi e proteine e stabilizza molte strutture proteiche di membrana. Nei mammiferi alcuni
glicolipidi formano gli antigeni dei gruppi sanguigni.
30
Una delle vie più comuni di segnalazione intracellulare si basa sull’uso di secondi messaggeri derivati dal
fosfolipide di membrana fosfatidilinositolo 4,5 bifosfato. PiP2. Agli inizi degli anni 80 quando si dimostrò che
l’aumento del calcio citosolico è preceduto dall’idrolisi di questo fosfolipide di membrana, in risposta ai segnali
cellulari. Questo fosfolipide appartiene ad una famiglia di inositolo fosfolipidi, con inositolo fosforilato (con 1 o 2
fosfati). Sebbene tutti e tre gli inositolo fosfolipidi possono essere degradati in risposta al segnale, la demolizione
critica è quella del PIP2, anche se è il meno abbondante costituendo il 10% di tutti gli inositolo fosfolipidi e meno
dell’1% di tutti i fosfolipidi. L’idrolisi di questo fosfolipide operata dalla fosfolipasi C porta a 2 prodotti: 1,2
diacil-glicerolo che resta nella membrana e l’inositolo 1,4,5 trifosfato (IP3) idrosolubile. Una proteina G accoppia i
recettori ormonali con la fosfolipasi C; il trattamento con la tossina della pertosse inattiva G e abolisce
l’attivazione della fosfolipasi C. L’IP3 diffonde nel citosol, si lega a suoi recettori sul reticolo endoplasmico e
promuove il rilascio del calcio. Il diacil-glicerolo rimane nella membrana e aiuta ad attivare la protein chinasi C.
Vi sono tre classi di fosfolipasi C (, , ) la classe beta viene attivata da recettori legati a proteine G. La classe
gamma contiene domini SH2 che ne mediano l’attivazione da parte di recettori tirosin chinasici. Quindi questo è
una seconda via di segnalazione che utilizza ’inositolo fosfolipide senza una proteina G intermediaria. Dopo un
secondo dalla sua formazione l’IP 3 viene idrolizzato a inositolo 1,4-difosfato, molecola che non determina rilascio
di altri ioni calcio. In pochi minuti gli store vengono esauriti ed il mantenimento di elevati livelli di calcio
citosolico richiede l’ingresso dall’ambiente extracellulare. Nelle cellule pancreatico tale ingresso è mediato
dall’inositolo 1,3,4,5 tetrafosfato (IP4) derivante dalla fosforilazione di IP3.
31
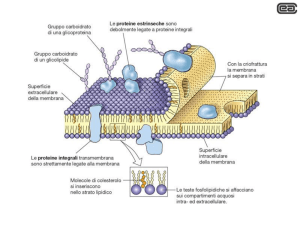
La componente proteica della membrana è altamente variabile sia in funzione del tipo di cellula e del tipo di
membrana cellulare. Le proteine della membrana mitocondriale sono estremamente diverse dalle proteine della
membrana plasmatica; anche i componenti della membrana plasmatica di una cellula epatica o intestinale sono
differenti. Tali proteine possono avere strutture e funzioni diverse. Alcune posseggono funzione di trasporto di
nutrienti o ioni come quella rappresentata in Figura, che corrisponde alla struttura tridimensionale di un sistema di
scambio Na/H in un batterio, il controtrasporto NHaA; altre hanno funzione di canali ionici come quello riportato
in figura corrispondente al canale per il potassio. Altre ancora hanno funzione recettoriale come il recettore a sette
eliche correlato alle proteine G nella trasduzione del segnale; altre ancora come la glicoforina intervengono nelle
interazioni tra citoscheletro e la membrana cellulare.
Come sono integrate queste proteine con il doppio strato lipidico; alcune sono legate solo superficialmente alla
membrana e sono denominate per questo motivo estrinseche, altre presentano una regione immersa nella membrana
e domini su 1 o ambedue i lati di essa e sono denominate proteine intrinseche di membrana.
32
Le proteine di membrana svolgono un ruolo importantissimo nel mantenimento della struttura e
dell’attività cellulare e si stima che rappresentino circa un terzo di tutte le proteine codificate
dal DNA. Ogni cellula esprime da 10 a 50 tipi diversi di proteine di membrana che posso
essere raggruppate in tre grandi classi.
Le proteine integrali interagiscono fortemente con il doppio strato lipidico e possono essere
rimosse soltanto dopo la distruzione della membrana con l’uso di detergenti. Alle proteine
integrali appartengono le proteine transmembrana composte da uno o più segmenti che
attraversano l’intero spessore della membrana e le rendono accessibile da entrambi i lati.
Le proteine estrinseche interagiscono debolmente con le regioni polari del doppio strato
lipidico e con le proteine integrali di membrana. Sono accessibili da un solo lato (interno o
esterno) e possono essere rimosse facilmente senza distruggere l’integrità di membrana. Ad
esse appartengono vari enzimi di membrana (es. enzimi del lume dell’intestino tenue,
l’adenilato ciclasi, la fosfolipasi C, l’anidrasi carbonica, ecc.) e alcune proteine strutturali che
ancorano il citoscheletro alla membrana cellulare.
33
La > parte delle proteine integrali sono trattenute nella membrana dalle catene laterali aminoacidiche non polari.
Esse contengono domini che si estendono nel mezzo acquoso su ogni lato del doppio strato. Nella > parte dei casi i
segmenti che attraversano il doppio strato formano un alfa-elica. Questo configurazion è energeticamente favorita
dall’assenza di acqua nel doppio strato lipidico; di conseguenza tutti i legami peptidici compresi nel doppio strato
tendono a formare legami idrogeno tra loro e non con molecole di acqua. La necessità di formare legami idrogeno a
distanze regolari è anche la ragione per cui una catena polipeptidica attraverserà sempre integralmente il doppio
strato lipidico. Le proteine integrali sono tenute incastrate nella membrana da due principali interazioni: le
interazioni ioniche con i gruppi polari dei lipidi e quelle idrofobiche con i lipidi interni di membrana. Ambedue i
tipi di interazione favoriscono l’ancoraggio della glicoforina, proteina della membrana eritrocitaria, al doppio strato
lipidico. La parte della glicoforina che prende contatto con la membrana consiste in 34 residui (62-95) compresa
una sequenza di 23 aacidi altamente idrofobica comprendente, fenilalanina, leucina, isoleucina, valina, triptofano,
tirosina. Il grafico di idropatia della glicoforina è riportato in figura; in questo grafico il segno positivo indica che è
necessaria energia libera per il trasferimento del segmento in questione all’acqua e quindi si tratta di un segmento
idrofobico, viceversa se l’indice è negativo si tratta di regioni idrofiliche. Nel caso della glicoforina la regione
positiva cade proprio nell’intervallo compreso tra 62- 95 aacidi che viene di conseguenza essere reputato il
segmento della proteina che attraversa il doppio strato lipidico.
Poiché è possibile fare un grafico del genere per regioni superiori a 10 aacidi (un’alfa elica è generalmente di 2030 aacidi idrofobici), l’inserimento di proteine integrali che spannano il doppio strato tramite foglietti beta che sono
in genere costituiti da meno di 10 aacidi, non può essere analizzato tramite studi di idropatia.
34
Il canale del potassio di s. l. è una proteina integrale tetramerica con sequenza omologa a tutti i canali del potassio
conosciuti, particolarmente nella regione del poro. L’analisi a raggi X con risoluzione di 3.2 angstroms ha
dimostrato che 4 identiche sub-unità creano un cono, che contiene il filtro di selettività del poro verso il lato
extracellulare. Sia l’ingresso intra- che quello extracellulare sono carichi negativamente per la presenza di aacidi
acidi, questo comporta un aumento della concentrazione locale dei cationi ed una diminuzione della concentrazione
locale degli anioni. La lunghezza complessiva del poro è di circa 45 angstrom, nel cui interno vi è lLo stretto filtro
di selettività, lungo solo 12 angstrom, mentre la parte restante del poro è ampia e delimitata da aminoacidi
idrofobici probabilmente inerti. Un grande cavità riempita di acqua e dipoli ad elica sono posizionate in modo da
impedire la destabilizzazione elettrostatica di uno ione in un poro al centro del doppio strato lipidico a basso
capacità dielettrica. Infatti gli ioni potassio permeano nel poro interno rimanendo idratati. Al contrario il filtro di
selettività è così stretto che gli ioni devono perdere il mantello acquoso per entrarvi. La composizione chimica della
parete delimitante la parte interna del poro è in maniera predominante idrofobica. Al contrario il filtro di selettività
è limitato esclusivamente da catene polari che appartengono alla caratteristica sequenza dei canali per il potassio.
Gli atomi di ossigeno della catena principale delimitano il filtro di selettività, che è tenuto aperto da costrizioni
strutturali, in modo da coordinare ioni potassio anziché ioni Na di dimensione minore. Il filtro di selettività
contiene 2 ioni potassio a distanza di circa 7.5 angstroms. Tale configurazione permette la conduzione degli ioni
sfruttando le forze elettrostatiche repulsive per superare le forze attrattive tra gli ioni potassio ed il filtro di
selettività. L’architettura del poro stabilisce i principi fisici che sono alla base della selettiva conducibilità del
potassio.
L’architettura del poro probabilmente riflette quella posseduta da altri canali del sodio e del calcio.
35
La mappa 3D di NhaA a risoluzione di 7 vista normale al piano della membrana. Si possono vedere i 12 profili di
densita con forma a bacchetta corrispondenti alle eliche trans-membrana, con una densità per ciascuna delle eliche
non continua attraverso il bilayer. Due possibilità per la collocazione del patway di traslocazione sono evidenziate
con gli asterischi. Nell’inset, sezioni orizzontali prese dal centro della mappa 3D con la proteina nello stesso
orientamento e le eliche arbitrariamente numerate da 1 a 12 (non corrispondenti all’assegnazione). La vista dal lato
delle eliche mostrata in Fig.3 corrispondono alla prospettiva del punto X nell’inset. L’eliche blue sono mostrate in
Fig.3a, eliche magenta in Fig.3b ed eliche gialle in Fig.3c.
Fig.2 Sezioni orizzontali attraverso i dimeri di NhaA. I monomeni sono mostrate in giallo e blue. A. 8 A sopra il
centro della membrana; b. al centro della membrana; c; sotto il centro della membrana. In ciascuna sezione, si
vedono 12 picchi per ciascun monomeno di NhaA.
Fig.3 Sezioni verticali attraverso la mappa 3D di NhaA. La molecola è ruotata di 90° per rivelare le eliche
perpendicolari alla membrana dalla prospettiva di X di Fig.1 (inset). a. Le sei eliche che mediano i contatti intra ed
inter dimero (blue nell’inset di Fig.1); b. le tre lunghe eliche inclinate (magenta nell’inset di Fig.1) che si rivelano
quando il primo strato delle 6 eliche viene rimossi; c. il gruppo di tre eliche non completamente risolto (gialle
nell’inset di fig.1) che si vedono quando lo strato delle tre eliche in b è rimosso.
Fig.4 Interazione tra dimeri adiacenti di NhaA. La mappa è contornat in magenta. La frecce bianca mostra i contatti
interdimerici tra eliche di dimeri adiacenti.
36
Nelle cellule eucariote un gran numero di proteine sono legate alla membrana tramite lipidi legati covalentemente.
Si dividono in tre classi:
Le proteine della superficie cellulare appartenenti alla prima classe come thy-1, fosfatasi alcalina, trealasi, sono
ancorate alla superficie esoplasmatica tramite complesso fosfolipidico glicosilato che contiene Nacetilglucosammina ed inositolo. l trattamento con fosfolipasi C, enzima che rompe il legame fosfatoglicerolo, stacca le proteine dalla superficie cellulare. A questa schiera appartengono diverse proteine
coinvolte nei meccanismi di adesione cellulare ed in particolare la caderina, localizzata tra le altre sulla
membrana cellulare dei miocardiociti dove oltre ad avere un ruolo nel riconoscimento cellulare importante per
la crescita del tessuto potrebbe secondo alcune ipotesi funzionare come spugna del calcio, favorendo così i
processi di contrazione miocardica.; Una seconda classe di proteine è ancorata alla faccia citoplasmatica della
membrana tramite l’acido miristico, acido grasso saturo a 14 atomi di carbonio (legame ammidico al residuo
di glicina presente sull’azoto terminale di queste proteine). Fa parte di questo gruppo la v-src, mutante della csrc; entrambe sono tirosina protein-chinasi (aggiunge gruppi Pi a tirosine di alcune proteine). V-src è una
proteina trasformante in quanto determina la trasformazione o cancerogenesi di cellule in coltura, fosforilando
altre proteine, solo se è legata alla membrana plasmatica. Infine un terzo gruppo di proteine a cui appartiene la
P21 ras, GTPasi monomerica sono ancorate alla faccia citoplasmatica della membrana tramite residuo
farnesilico legato con legame tioestere ad un residuo di cisteina localizzato vicino all’estremità
37
Come i lipidi di membrana le proteine non passano attraverso il doppio strato ma possono invece ruotare intorno ad
un asse perpendicolare al piano del doppio strato (diffusione rotazionale); molte proteine sono capaci di muoversi
lateralmente nella membrana (diffusione laterale). La prova diretta di ciò si ebbe nel 1970 tramite un esperimento
in cui le cellule di topo vennero fuse artificialmente con cellule umane per produrre cellule ibride (eterocarionti).
Due anticorpi diversi marcati con rodamina o fluresceina, distinguibili al microscopio a fluorescenza furono usati
per distinguere proteine umane da quelle di topo. Sebbene inizialmente le proteine umane e quelle di topo fossero
confinate nelle rispettive metà dell’eterocarionte appena formato, le due serie di proteine diffondevano e si
mescolavano nel giro di mezz’ora.
I valori dei coefficienti di diffusione per le proteine di membrana sono molto variabili, ma variano in genere da 1
decimo ad un 100 dei valori corrispondenti per le molecole fosfolipidiche di una stessa membrana. Questi dati
confermano ancora una volta la validità del modello a mosaico fluido proposto da Nicolson nel 1972.
La visione della membrana come un mare lipidico in cui tutte le proteine galleggiano liberamente è una
semplificazione eccessiva. Molte cellule sono infatti in grado di confinare le proteine di membrana entro domini
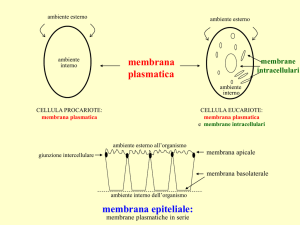
specifici del doppio strato continuo di lipidi. Nelle cellule epiteliali intestinali e renali alcuni enzimi o sistemi di
trasporto sono localizzati sulla porzione apicale altri su quella basolaterale della membrana plasmatica. Questa
separazione avviene solo tra lipidi del monostrato esterno tramite l’esistenza di giunzioni intercellulari le tight
junctions. La membrana plasmatica degli spermatozoi consiste in almento tre domini distinti, al di fuori dei quali
lipidi e proteine appartenenti ad un domino, non possono diffondere.
Le cellule hanno modi ancora più drastici per impedire la diffusione laterale delle proteine. I grossi aggregati come
la molecola di batteriorodopsina diffonde molto lentamente. Un modo ancora più comune per evitare la diffusione è
quello di agganciare i complessi proteici al citoscheletro interno
38
E’ questo il caso di alcune proteine di membrana dei globuli rossi. L’ Eritrocita ha una forma di disco
biconcavo di 7 micrometri di diametro. E’ nella sua forma matura, privo di nucleo. Il citoscheletro
sostiene la membrana ed è unita ad essa in molti punti. Due principali proteine integrali sono presenti:
glicoforina e proteine della banda 3 o scambio anionico. La banda 3 è un dimero costituito da 2 catene di
929 aacidi ognuna; il segmento C-terminale di ogni catena è inserito nella membrana plasmatica che
attraversa molte volte formando 14 eliche trans-membrana. Il segmento N-terminale si ripiega in
filamenti idrosolubili che sporgono nel citosol ed ancorano il citoscheletro alla membrana. I maggiori
costituenti del citoscheletro sono le spettrine alfa e beta (bande 1 e 2), i polipeptidi che hanno peso
molecolare più alto. Si combinano a formare tetrameri (alfa-beta)2 ; le terminazioni libere dei tetrameri
sono tenute insieme da corte catene di actina (banda 5); il filamento di actina è costituito da 13
monomeri di actina ed un filamento di tropomiosina. La proteina della banda 4.1 e l’adducina, altre due
proteine del citoscheletro, favoriscon interazione actina – spettrina. Una singola fibra di actina si lega a
più molecole di spettrina, costituendo un reticolo. L’ancoraggio alla membrana eritrocitaria avviene
tramite l’anchirina che ha due domini: uno si lega strettamente e specificamente ad un sito beta della
spettrina vicino al centro del tetramero, l’altro ad un dominio citosolico della banda 3. La proteina della
banda 4.1 si lega al domino carbossi-terminale della glicoforina e anche della banda 3. Anche queste
interazioni possono contribuire all’unione dello scheletro della spettrina alla membrana. Patologie
ereditarie come la sferocitosi e l’ellittocitosi sono determinate da mutazioni geniche. Il difetto si verifica a
causa di peptidi anomali di spettrina che si legano male a proteina 4.1 ed anchirina. In altri casi
l’anchirina è difettosa o assente. Tali eritrociti difettosi sono più rapidamente eliminati dalla milza e di
conseguenza in questi pazienti si possono verificare anemie.
39
E’ ormai accertato che nelle membrane esistono delle zone specializzate, dei microdomini, chiamati raft,
in cui predominano alcuni lipidi. Queste zone sono caratterizzate dal non essere solubilizzate da
detergenti non-ionici, tanto è vero che hanno acronimi diversi come DRMs (detergent-resistant
membranes) and Digs (detergent-insoluble glycolipid-enriched membrane domains. In alcune zone della
membrana, esistono delle regioni le caveole, che sono zone morfologicamente distinguibili come
invaginazioni delle membrane plasmatiche, caratterizzate dalla presenza di proteine denominate
caveoline. Una delle ragioni per le quali queste regioni della membrana sono attualmente oggetto di
studio è che queste possano essere coinvolte nei meccanismi di trasduzione del segnale.
Queste regioni sono ricche di glicosfingolipidi, colesterolo e di proteine che non hanno una forma
particolare. La loro dimensione è compresa tra 70 e 300 nm. Oltre a queste molecole nel monostrato
esterno sono presenti proteine ancorate al fosfatidil-inositolo. Nel monostrato interno sono invece
localizzate proteine tipo Ras, coinvolte nella trasduzione del segnale. Ciò rafforza il concetto che i domini
RAFT agiscano come piattaforma per concentrare molecole segnale in una regione particolare della
membrana e anche per raggruppare specifiche proteine ancorate al fosfatidil-inositolo. Questi raft non
rimangono stazionari nella membrana, ma si muovono lateralmente, e la loro mobilità è fortemente
infuenzata dal contenuto di colesterolo della membrana. In particolare la riduzione del colesterolo riduce
la mobilità dei raft.
Le caveole sono caratterizzate dalla loro associazione con le caveoline. Queste proteine interagiscono
con parecchie molecole segnale nella membrana e costituiscono l’impalcatura per organizzare i
preassemblati complessi della segnalazione. Differenti molecole segnalanti sono associate a Digs e
caveole. Digs si associano a Ras e molecole correlate a Ras. Domini ricchi di caveoline contengono src,
proteine G, protein chinasi C e RhoA.
I lipidi di membrana possono assumere anche delle configurazioni non-lamellari. Questo determina una
regione micellare nel piano trasversale della membrana che aumenta lo spessore locale e la curvatura
della membrana, in relazione alla morfologia piatta e planare del resto della membrana. L’organizzazione
biochimico e fisica dei non-bilayer lipid è differente dagli altri lipidi, in particolare per la grandezza della
loro testa polare e delle loro catena acilica. Generalmente, in regioni non a doppio strato, molecole
lipidiche con una testa polare più piccola e catene alifatiche più lunghe (fosfatidil-etanolammina) sono
40
impaccate densamente sul piano laterale, determinando un incremento della curvatura su uno o su
entrambi gli strati. Questa alterazione nel doppio strato lipidico esercita una pressione laterale o stress di
curvatura, sulle proteine presenti in quella zona, che è stato dimostrato essere essenziale per il
funzionamento della batteriorodopsina e della protein chinasi C. Infatti fosfatidiletanolammnina è stata
rinvenuta in strutture cristalline di diverse proteine di membrana.
40
E’ ormai accertato che nelle membrane esistono delle zone specializzate, dei microdomini, chiamati raft, in cui
predominano alcuni lipidi. Queste zone sono caratterizzate dal non essere solubilizzate da detergenti non-ionici,
tanto è vero che hanno acronimi diversi come DRMs (detergent-resistant membranes) and Digs (detergentinsoluble glycolipid-enriched membrane domains. In alcune zone della membrana, esistono delle regioni le
caveole, che sono zone morfologicamente distinguibili come invaginazioni delle membrane plasmatiche,
caratterizzate dalla presenza di proteine denominate caveoline. Una delle ragioni per le quali queste regioni della
membrana sono attualmente oggetto di studio è che queste possano essere coinvolte nei meccanismi di trasduzione
del segnale.
Queste regioni sono ricche di glicosfingolipidi, colesterolo e di proteine che non hanno una forma particolare. La
loro dimensione è compresa tra 70 e 300 nm. Oltre a queste molecole nel monostrato esterno sono presenti proteine
ancorate al fosfatidil-inositolo. Nel monostrato interno sono invece localizzate proteine tipo Ras, coinvolte nella
trasduzione del segnale. Ciò rafforza il concetto che i domini RAFT (ZATTERA) agiscano come piattaforma per
concentrare molecole segnale in una regione particolare della membrana e anche per raggruppare specifiche
proteine ancorate al fosfatidil-inositolo. Queste zattere non rimangono stazionarie nella membrana, ma si muovono
lateralmente, e la loro mobilità è fortemente infuenzata dal contenuto di colesterolo della membrana. In particolare
la riduzione del colesterolo riduce la mobilità dei raft.
Le caveole sono caratterizzate dalla loro associazione con le caveoline. Queste proteine interagiscono con
parecchie molecole segnale nella membrana e costituiscono l’impalcatura per organizzare i preassemblati
complessi della segnalazione. Differenti molecole segnalanti sono associate a Digs e caveole. Digs si associano a
Ras e molecole correlate a Ras. Domini ricchi di caveoline contengono src, proteine G, protein chinasi C e RhoA.
I lipidi di membrana possono assumere anche delle configurazioni non-lamellari. Questo determina una regione
micellare nel piano trasversale della membrana che aumenta lo spessore locale e la curvatura della membrana, in
relazione alla morfologia piatta e planare del resto della membrana. L’organizzazione biochimico e fisica dei nonbilayer lipid è differente dagli altri lipidi, in particolare per la grandezza della loro testa polare e delle loro catena
acilica. Generalmente, in regioni non a doppio strato, molecole lipidiche con una testa polare più piccola e catene
alifatiche più lunghe (fosfatidil-etanolammina) sono impaccate densamente sul piano laterale, determinando un
incremento della curvatura su uno o su entrambi gli strati. Questa alterazione nel doppio strato lipidico esercita una
pressione laterale o stress di curvatura, sulle proteine presenti in quella zona, che è stato dimostrato essere
41
essenziale per il funzionamento della batteriorodopsina e della protein chinasi C. Infatti fosfatidiletanolamina è
stata rinvenuta in strutture cristalline di diverse proteine di membrana.
41
L’internalizzazione dei ligandi inizia quando per diffusione nella membrana il complesso recettore-ligando si
localizza sopra le fossette rivestite; queste poi si separano dalla membrana sotto forma di vescicole rivestite. Nella
stessa fossetta rivestita o nella vescicola si possono rintracciare due tipi di ligandi legati a recettori di superficie, la
transferrina e le LDL. Il tessuto nervoso ne è particolarmente ricco. Esse possono essere coinvolte nei processi di
trasporto di proteine dall’apparato di golgi alla superficie cellulare. Il 2% della superficie cellulare di epatociti e
fibroblasti è occupata da fossette rivestite. Dimostrazione al microscopio elettronico delle fasi iniziali
dell'endocitosi mediata da recettore di particelle LDL in fibroblasti in coltura. (a) Una fossetta rivestita; i piccoli
punti visibili sopra la stessa rappresentano particelle LDL marcate con ferritina. (b) Una fossetta contenente LDL
apparentemente chiusa su se stessa a formare una vescicola rivestita. {c) Una vescicola rivestita contenente
particelle LDL. (d) Particelle marcate con ferritina in un endosoma a superficie liscia, 6
minuti dopo l'esposizione delle cellule al composto marcato.
42
Biosintesi della fosfatidiletanolammina nelle cellule animali. I precursori sono un acido grasso-acetil CoA (una
molecolaanfipatica immersa nella membrana del RE), il glicerolo 3-fosfato e la citidina difosfo-etanolammina
(CDP-etanolammina) (due molecole solubili in acqua presenti nel citosol). Nella membrana del RE vengono
assemblati altri lipidi tramite analoghe vie metaboliche che partono da acido grasso-acetil CoA e piccole molecole
solubili. Per la sintesi della fosfatidiletanolammina, le cellule batteriche utilizzano una diversa via metabolica che
parte anch' essa da un acido grasso-acetil CoA e ha luogo nella membrana plasmatica batterica.
Figura 17.4 Test per la rivelazione di una flippasi di fosfolipidi. Il fosfolipide dibutirrilfosfatidilcolina è
idrosolubile perche contiene corte catene di acidi grassi a quattro atomi di carbonio. Se addizionato a una
sospensione di vescicole composte da fosfolipidi puri (liposomi), esso si inserisce spontaneamente nel foglietto
esterno ma, in assenza di un enzima di natura proteica chiamato flippasi, non può spostarsi nel
foglietto interno. la proteina flippasi, come si trova nelle membrane del RE, catalizza i movimenti di questo piccolo
fosfolipide verso il foglietto interno (esoplasmatico); da lì, esso può spostarsi spontaneamente nel
lume della vescicola dove trova un ambiente acquoso. Quindi, si può saggiare la presenza di una flippasi
verificando se vi è una proteina che permette alla dibutirrilfosfatidilcolina, aggiunta alI' esterno di una vescicola, di
accumularsi nel lume della stessa. (Da W.R. Bishop e R.M. BelI, 19B5, CelI, 42, pp. 50-60; Y. Kawashimae R.M.
BelI, 1987, I. Bio/. Chem., 262. 00. 16495-16502.
43