LIPIDI
Riserve energetiche (trigliceridi), sono molecole più ridotte rispetto agli
zuccheri e dalla loro ossidazione viene liberata una quantità di energia
maggiore.
Costituzione delle membrane biologiche (glicerofosfolipidi, sfingolipidi)
Isolamento e rivestimento termico, protezione delle superfici (cere)
Funzioni altamente specializzate, di tipo ormonale, riconoscimento
cellulare.
Sono molecole insolubili in soluzione acquosa.
Possono essere totalmente idrofobici oppure anfipatici.
Costituenti basilari dei lipidi sono gli ACIDI GRASSI
Acidi grassi: Composti da una coda idrocarburica ± lunga e una funzione acida
(COOH in posizione 1)
I più comuni possiedono
da 12 a 20 atomi di carboni
Possono essere saturi,
Monoinsaturi o poliinsaturi
(il primo doppio legame è
solitamente in posizione 9-10,
i successivi in posizione 12-13,
e 15-16, non sono coniugati).
In genere il doppio legame è CIS
e viene indicato nella
nomenclatura con il simbolo Δn
n = posizione del doppio legame
Ac. ottadecanoico
Ac.
Ac. Stearico
cis-Δ9-ottadecanoico
Ac. Oleico
Ac. Linoleico
Ac. linolenico
Ac. cis-Δ9,12-ottadecanoico
Ac. cis-Δ9,12,15-ottadecanoico
LIPIDI DERIVANTI DAL GLICEROLO
TRIGLICERIRIDI
(o TRIACILGLICEROLI)
GLICEROFOSFOLIPIDI
LIPIDI DERIVANTI DALLA SFINGOSINA
H
H
SFINGOMIELINE
GLICOLIPIDI
LIPIDI CON NUCLEO STEROIDEO (STEROLI)
COELSTEROLO
AC. BILIARI
ORMONI STEROIDEI
TRIGLICERIDI
Lipidi di riserva energetica. Dalla loro
degradazione e ossidazione si ricava energia.
sono lipidi NEUTRI, totalmente APOLARI
Sono immagazzinati in forma anidra nelle cellule
adipose sotto forma di vacuoli limitati da
membrane, da cui sono mobilizzati per produrre
energia in condizioni di digiuno (es. durante il
sonno).
Glicerolo + acidi grassi
(legati attraverso legame ESTERE)
glicerolo
Esistono Triacilgliceroli
Diacilgliceroli
Monoacilgliceroli,
1-stearil,2-linoleil,3-palmitoil glicerolo
GLICEROFOSFOLIPIDI (costituenti delle membrane biologiche)
Struttura base dei glicerofosfolipidi: acido fosfatidico o fosfatide
Composto da: Glicerolo 3-fosfato esterificato con 2 acidi grassi (C-1 e C-2)
Legami estere
H
Legame estere
glicerolo 3-fosfato
Ac. fosfatidico
I fosfatidi si differenziano per la natura delle loro 2 code idrocarburiche (per il n° di
atomi di Carbonio e le insaturazioni).
Il gruppo fosfato solitamente forma un secondo legame estere con un’altra molecola
che porta una funzione alcolica e con cui costituisce così la “testa polare” del
glicerofosfolipide
Testa polare
idrofilica:
fosfato + colina
I GLICEROFOSFOLIPIDI
SONO MOLECOLE
ANFIPATICHE
Coda idrofobica:
glicerolo + catene
idrocarburiche (stearato
in C1 e oleato in C2)
- in C2 solitamente una
catena acilica insatura -
Per studiare la struttura dei glicerofosfolipidi e identificare i loro componenti, essi
devono essere idrolizzati con enzimi specifici: le lipasi.
Fosfolipasi A2 (idrolizza
il legame estere in
posizione 2)
Fosfolipasi A1 (idrolizza
il legame estere in
posizione 1)
H
GLICEROLO H
H
IDROLISI COMPLETA
H
2 AC. GRASSI
HO-X
Fosfolipasi C (idrolizza il
legame estere fra il fosfato
e il glicerolo)
Fosfolipasi D (idrolizza
il legame estere fra il
fosfato e la testa polare)
H
TESTA POLARE
FOSFATO
Le fosfolipasi sono presenti nel succo pancreatico e servono a digerire i fosfolipidi
assunti con la dieta.
Il veleno di serpenti, api e vespe contiene la fosfolipasi A2; concentrazioni molto
elevate di quest’enzima nel sangue può causare la lisi delle membrane plasmatiche,
perché va a idrolizzare i glicerofosfolipidi che la costituiscono.
Glicerofosfolipidi in cui in posizione 1 la coda
idrocarburica è legata attraverso un legame etere,
mentre in C-2 il gruppo acilico è legato mediante
legame estere
PLASMALOGENI
Abbondanti nelle membrane dei neuroni (cervello e nervi periferici),
nel tessuto muscolare e cardiaco
Particolarmente resistenti
alle fosfolipasi
Legame etere con una catena idrocarburica alchilica
Agisce nel processo di
coagulazione del
sangue e riparazione
delle ferite.
Secreto dai leucociti
nel sangue, stimola le
piastrine ad aggregarsi
e a secernere
serotonina
(vasocostrittore).
Ac. Acetico esterificato in C-2
La testa polare contiene colina.
FATTORE ATTIVANTE LE PIASTRINE
SFINGOLIPIDI (costituenti delle membrane biologiche)
La molecola base è la sfingosina (amminoalcol) con una lunga catena idrocarburica
monoinsatura che parte dal C-3, gruppi OH legati in C-3 e C-1 e un gruppo amminico
in C-2.
CERAMMIDE: Struttura base degli sfingolipidi = sfingosina + 1 ac. grasso (OH in C-1 libero)
H
H
Sfingomieline
Cerebrosidi
Globosidi
Gangliosidi
La sfingosina lega 1 solo acido grasso attraverso
un legame CARBOAMMIDICO
Catena acilica
dell’ac. grasso
il cerammide legherà la testa
polare sul carbonio 1
testa polare
fosfocolina o fosfoetanolammina tramite legame estere
monosaccaride (Glucosio o galattosio tramite legame glicosidico)
Di-, Tri-, Tetra-saccaride (legame glicosidico)
oligosaccaride complesso (legame glicosidico)
Sfingomieline:
Sono ANFIPATICHE
Sono Fosfolipidi
Coda idrofobica:
catena idrocarburica
della sfingosina e
catena acilica dell’ac.
grasso
Cerebrosidi, globosidi e gangliosidi sono GLICOSFINGOLIPIDI o GLICOLIPIDI
Non sono fosforilati
Cerebrosidi e globosidi sono glicolipidi neutri
Gangliosidi: possono avere uno o più residui di zuccheri acidi, hanno una testa polare
carica negativamente a pH 7. La testa polare è molto voluminosa
GLUCOSILCERAMMIDE
LATTOSILCERAMMIDE
Glucosio legato con
legame β glicosidico
GANGLIOSIDE
STEROLI
Lipidi strutturali delle membrane plasmatiche
Nucleo steroideo, rigido e planare
Anello tetraciclico condensato: 3 cicloesani e 1 ciclopentano
porzione idrofobica
Sterolo più abbondante nelle
membrane delle cellule animali
Anfipatico
Testa polare.
Il colesterolo libero è
esterificato in questa
posizione con un ac. grasso
Nelle piante = stigmasterolo
Nei funghi = ergosterolo
Gi steroli sono precursori di composti
steroidei con attività specifiche
ACIDI BILIARI = liberati
nell’intestino durante la
digestione agiscono come
detergenti dei grassi favorendo
a loro degradazione
ORMONI STEROIDEI = controllano
la funzionalitò di diversi organi e
regolano l’espressione genica e il
metabolismo, la riproduzione
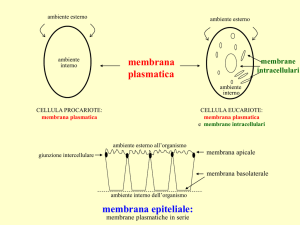
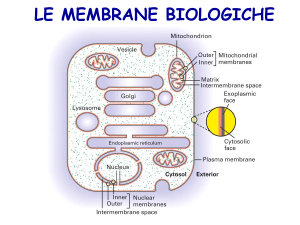
LE MEMBRANE CELLULARI
FUNZIONI:
- Protezione
- Sostegno
- Scambio di molecole e segnali
- Siti di reazioni metaboliche
Formate dall’aggregazione di lipidi strutturali, proteine e glucidi.
La loro composizione varia nei diversi tipi cellulari, nelle diverse specie e nei diversi
organuli delle cellule eucariotiche.
La loro composizione riflette la loro specializzazione funzionale.
Sono impermeabili a soluti polari o carichi e sono permeabili a
soluti non polari, il passaggio di molecole polari e ioni attraverso
la membrana è regolato da specifici sistemi di trasporto
La struttura delle membrane biologiche rispetta il
MODELLO A MOSAICO FLUIDO
La membrana biologica è costituita da un
DOPPIO STRATO LIPIDICO che racchiude un
compartimento acquoso interno e lo separa da
quello esterno.
I lipidi di membrana sono tutti lipidi anfipatici :
Fosfolipidi (glicerofosfolipidi e sfingomieline)
Glicolipidi
Colesterolo
Superficie
idrofilica
interno
Superficie
idrofilica
L’effetto idrofobico è la forza che traina i
lipidi anfipatici ad aggregarsi in un doppio
foglietto:
- l’acqua viene totalmente esclusa
dall’interno del doppio foglietto, le code
idrocarburiche dei lipidi costituiscono un
ambiente interno anidro.
- Le code idrocarburiche dei lipidi si
impacchettano strettamente e
stabiliscono interazioni idrofobiche e di
van der Waals che stabilizzano il doppio
foglietto.
- Le teste polari interagiscono tra loro e
con le molecole d’H2O.
La distribuzione dei lipidi è
ASIMMETRICA nella membrana
cellulare: i due foglietti non hanno la
stessa composizione e questa può
variare in funzione dei ruoli biologici
che la membrana assume.
Il doppio foglietto lipidico delle membrane biologiche è una struttura fluida e dinamica: i
lipidi possono diffondere lungo lo stesso foglietto da un punto ad un altro (diffusione
laterale).
rapida
È molto lenta invece la diffusione trasversale, dal foglietto interno a quello esterno e
viceversa (movimento a flip-flop) e quando avviene è guidato da enzimi chiamati Flippasi e
Floppasi, e avviene per ragioni funzionali precise.
Per es.: il passaggio della fosfatidilserina dal
foglietto interno a quello esterno della membrana
plasmatica è uno dei segnali che indirizzano la
cellula verso la morte programmata (apoptosi)
La FLUIDITA’ del doppio strato lipidico dipende da:
a) temperatura
b) tipo di lipidi (natura delle code idrocarburiche)
Sopra la Temperatura di transizione
Temperatura
di
transizione
Sotto la Temperatura di transizione
A temperature superiori
alla temperatura di
transizione si trova in
uno stato liquido
disordinato (rotazioni
continue intorno ai
legami C-C)
A temperature intermedie: STATO LIQUIDO
ORDINATO (il movimento delle catene è
minore, avviene uno spostamento laterale dei
lipidi). È lo stato in cui si trovano le membrane
biologiche a temperature corporee
A temperature relativamente basse il doppio strato
lipidico si trova in uno stato di gel semisolido o
paracristallino (movimenti limitati)
Se consideriamo una temperatura compresa fra i 20 e i 40 °C la fluidità del doppio
strato lipidico aumenta con l’aumentare di catene idrocarburiche insature e corte.
Catene più corte
Maggior grado di INSATURAZIONE
Catene più lunghe
Maggior grado di SATURAZIONE
MINORI CONTATTI DI VAN DER WAALS
MAGGIORI CONTATTI DI VAN DER WAALS
Impaccamento
più rilassato
Impaccamento
più stretto
Le curvature
imposte dai doppi
legami cis rigidi
allontanano le code
idrocarburiche.
Infatti, la temperatura di fusione di un acido grasso dipende dalla lunghezza della catena
idrocarburica e dalla presenza di insaturazioni.
T. Fusione stearato (saturo, 18 C) = 70°C
T. Fusione oleato (1 insaturazione, 18 C) = 13°C
T. Fusione linolenico (3 insaturazioni, 18 C)= -17°C
Anche gli STEROLI (colesterolo) quando presenti in una membrana
cellulare hanno lo stesso effetto delle insaturazioni: gli steroli hanno un
corpo idrofobico non lineare che crea delle interruzioni nel doppio
foglietto diminuendo le interazioni da impaccamento e rendendo la
membrana più fluida.
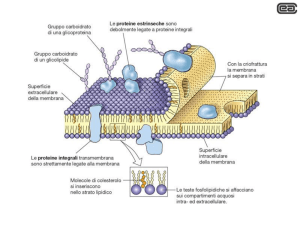
PROTEINE DI MEMBRANA
PROTEINE PERIFERICHE associate alla superficie della membrana tramite
interazioni elettrostatiche e legami idrogeno con le teste polari dei lipidi, o con le
porzioni esterne di proteine integrali.
PROTEINE INTEGRALI immerse nel doppio strato lipidico, a cui sono stabilmente
associate tramite interazioni idrofobiche con le code dei lipidi. Alcune proteine
attraversano completamente la membrana con un dominio transmembrana e
presentano dei domini che sporgono o nello spazio extracellulare o nel citosol o in
entrambi
α-eliche idrofobiche
β-foglietti idrofobici in un motivo a barile
Residui apolari
Residui
polari
Canale transmembrana per
l’attraversamento di soluti polari
PROTEINE TRANSMEMBRANA
Dominio transmembrana: αelica idrofobica
Dominio N-terminale
glicosilato esterno
Spazio
extracellulare
citosol
Dominio C-terminale
idrofilico citosolico
Le proteine di membrana sono disposte in modo asimmetrico
Proteine e lipidi di membrana si organizzano come tessere di un mosaico che sono tenute
insieme da interazioni stabili che nel contempo assicurano la fluidità; infatti i componenti
della membrana sono liberi di spostarsi lateralmente mantenendo intatta la membrana
stessa
glicolipidi
glicoproteine
fosfolipidi
Proteina periferica
Proteina integrale
transmembrana
colesterolo
α-elica
idrofobica
PROTEINE legate covalentemente alle ANCORE LIPIDICHE
Le ancore lipidiche sono Acidi grassi a catena lunga,
Isoprenoidi, e Derivati Glicosilati del
FosfatidilInositolo (GPI)
Sono inserite nel doppio strato lipidico
Ac. Palmitico legato al
residuo di Cys/Ser
della proteina
Ac. miristico (14 C) legato alla
Gly N-terminale della proteina
-
Ancora= Fosfatidilinositolo +
pentasaccaride + foetanolammina.
La proteina è legata col suo
C-terminale
Gruppo farnesilico (15 C) legato al
residuo di Cys C-terminale della
proteina