Corso di Chimica e Propedeutica Biochimica
Cinetica chimica ed enzimatica
2 H2 (g) + O2 (g) → 2 H2O (g)
1
Alcune immagini sono state prese e modificate da “Chimica” di Kotz, Treichel & Weaver, Edises 2007, III edizione
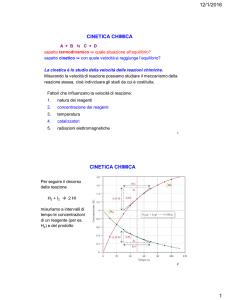
La cinetica chimica
A differenza della termodinamica che si occupa della stabilità
relativa tra reagenti e prodotti in una reazione chimica, la cinetica
chimica si occupa dello studio della velocità con cui avviene una
reazione chimica e della dipendenza di questa da vari fattori.
Oltre che della descrizione della dipendenza della velocità di
reazione da fattori sperimentali, la cinetica si occupa anche di
esaminare la sequenza dei processi chimici o fisici attraverso cui
ha luogo la conversione tra reagenti e prodotti. In questa maniera
lo studio della velocità di reazione mira ad ottenere informazioni
dettagliate sull’interazione tra le molecole nel corso della reazione.
2
La velocità di reazione
Il concetto di velocità si incontra in molti contesti anche non chimici
s
t
velocità
spazio percorso s
tempo impiegato t
La velocità di una reazione si definisce in modo analogo: essa corrisponde
alla variazione di concentrazione di una specie chimica nell’unità di tempo
velocità di reazione
variazione di concentrazione
intervallo di tempo
3
Si consideri la reazione di decomposizione del N2O5 che ha luogo
secondo la seguente reazione:
2 N2O5 (g) → 4 NO2 (g) + O2 (g)
• il progredire della reazione può essere seguito in diversi modi: attraverso
l’aumento della pressione parziale dell’O2 o di NO2 o la diminuzione di N2O5
• La quantità di O2 che si è formata è legata alla quantità di N2O5 inizialmente
presente: per ogni mole di O2 formatasi, 2 moli di N2O5 si sono decomposte
La velocità della reazione può essere espressa, per un qualunque intervallo di
tempo, come il rapporto tra la variazione di concentrazione di N2O5 e
l’intervallo di tempo trascorso
variazione di [N 2 O 5 ]
[N 2 O5 ]
velocità di reazione
intervallo di tempo
t
Il segno “” è dovuto al fatto che la [N2O5] diminuisce nel tempo.
4
Data la reazione:
k
A
B
la velocità di una reazione indica come cambia la concentrazione
dei reagenti o dei prodotti nel tempo.
velocità
media
[A ] [A]2 [A]1 [B]2 [B]1 [B]
v
t
t 2 t1
t 2 t1
t
[B]
[B]2
[B]
[B]1
t
t1
t2
t
5
[B]
È possibile definire la velocità istantanea come la derivata
rispetto al tempo della concentrazione di B (o A). Questa è la
tangente alla curva nel punto considerato.
A
[B] d[ B]
v lim
t 0 t
dt
k
B
[B]2
[B]
t
[B]1
unità di misura della velocità:
concentrazione / tempo
t1
t2
t
6
La velocità di reazione dipende da vari fattori:
•
•
•
•
•
la natura dei reagenti
la concentrazione dei reagenti
la temperatura di reazione
la presenza di catalizzatori
la superificie dell’interfaccia (se la reazione avviene tra
reagenti in due fasi diverse)
7
Dipendenza della velocità di reazione dalla concentrazione
Sperimentalmente si trova che la velocità di una reazione chimica dipende dalla
concentrazione dei reagenti. L’equazione che correla la velocità di una reazione
alle concentrazioni dei reagenti è nota come equazione cinetica o legge della
velocità ed ha la forma:
aA+ b B → c C + d D
m
v k [ A] [B]
n
cioè il prodotto delle concentrazioni dei reagenti elevate ad opportuni
coefficienti (i.e. m, n …, in genere diversi dai coefficienti stechiometrici)
moltiplicati per una costante (k). Questi coefficienti sono in genere (ma non
sempre) numeri interi e vanno determinati sperimentalmente.
La costante k è chiamata costante di velocità specifica o costante cinetica e
dipende solo dalla natura dei reagenti e dalla temperatura.
Ordine di reazione
Per una data reazione chimica di cui è nota l’equazione cinetica, si definisce
ordine di reazione rispetto ad un certo componente l’esponente della
concentrazione di quel componente nell’equazione cinetica
Si definisce invece ordine di reazione complessivo la somma degli esponenti di
tutti i reagenti presenti nell’equazione cinetica.
Ad esempio la generica reazione:
che ha equazione cinetica:
aA+ bB → c C+ d D
m
v k [ A ] [B]
n
è di ordine m rispetto al reagente A e di ordine n rispetto al reagente B e di
ordine m+n complessivo. Esempio, la reazione:
2 NO (g) + 2 H2 (g) → N2 (g) + 2 H2O (g)
con equazione cinetica v = k [NO]2[H2]
è del secondo ordine rispetto a NO, primo ordine rispetto a H2 e del terzo
ordine complessivo. Se un reagente non compare nell’equazione cinetica per
una data reazione questa è di ordine zero rispetto a tale reagente.
9
Determinazione sperimentale dell’ordine di reazione: metodo delle
velocità iniziali
La relazione tra velocità e concentrazione deve essere determinata
sperimentalmente. Un modo per affrontare questo problema è quello basato
sulla misura delle velocità iniziali (velocità misurata a t=0).
Misurare le velocità iniziale è conveniente perché le concentrazioni iniziali dei
reagenti è nota e in quanto evita possibili complicazioni derivanti da
interferenze da parte dei prodotti di reazione o dal verificarsi di altre reazioni.
CH3COOCH3 (aq) + OH (aq) → CH3COO (aq) + CH3OH (aq)
acetato di metile
ione ossidrile
acetato
metanolo
v [CH 3COOCH 3 ]m [OH ]n
La reazione è di ordine 1 rispetto all’acetato di metile e di ordine 1 rispetto
allo ione ossidrile.
10
CH3COOCH3 (aq) + OH (aq) → CH3COO (aq) + CH3OH (aq)
[acetato di metile]0 = 0.001 M, T = 25 °C
0.4 M
0.3 M
0.2 M
pendenza = n = 1
0.1 M
[OH]0
log
0.05 M
log
v 0 [CH 3COOCH 3 ]m0 [OH ]n0
log v 0 m log[CH 3COOCH 3 ]0 n log[OH ]0
11
La costante di velocità specifica k
La costante di velocità specifica k è la costante di proporzionalità che mette in
relazione velocità e concentrazione ad una data temperatura. È un parametro
importante perché, una volta noto, permette di calcolare la velocità di reazione
a qualunque valore di concentrazione.
La unità di misura della costante di velocità specifica deve essere consistente
con quelle degli altri termini contenuti nell’equazione cinetica.
k
A→
B
v = k∙[A] {k ≡ tempo1}
k
A+B→C+D
v = k∙[A]∙[B] {k ≡ concentrazione1∙tempo1}
12
Relazione tra concentrazione e tempo: leggi cinetiche integrate
Talvolta è utile e importante sapere per quanto tempo una reazione deve aver
luogo affinchè la concentrazione di un dato reagente raggiunga un dato valore
oppure prevedere il valore della concentrazione di un reagente o di un
prodotto dopo un certo tempo di reazione.
A tal scopo si deriva una equazione che mette in relazione le concentrazioni
ed il tempo: l’equazione cinetica integrata.
CH3COOCH3 (aq) + OH (aq) → CH3COO (aq) + CH3OH (aq)
L’equazione cinetica integrata
descrive la tempo dipendenza di
formazione dei prodotti o di
scomparsa dei reagenti
13
Reazione del 1° ordine (monomolecolare)
d[A ]
v
k [A ]
dt
k
A
B
l’equazione cinetica
d[A]
k dt
[A]
d[A]
[A] k dt separazione delle variabili
ln[A] k t Q
l’equazione tempo integrata, dove Q = costante
La costante d’integrazione Q può essere calcolata imponendo
la condizione che per t = 0, [A] = [A]0 → Q = ln[A]0:
ln[A] k t ln[A]0 ln[A] ln[A]0 k t
[A]
ln
k t
[A]0
[A ] [ A ]0 e k t
14
Reazione del 1° ordine (monomolecolare)
[A] [A]0 e
k t
k
A
B
il decorso temporale è esponenziale
k = 6.93 s1 t½ = 0.1 s [A]0 = 1 mM
dopo
t½ [A]= 0.5 mM
2 t½ [A]= 0.25 mM
3 t½ [A]= 0.125 mM
il tempo di
dimezzamento t½
[A ]0
se [A]
2
t
1
2
ln 2
k
15
Radioisotopi in biologia e medicina
•
•
•
•
sono usati come traccianti o come fonti di energia distruttiva (radioterapia)
devono avere semivita di giorni o mesi
non devono accumularsi nell’organismo
si devono trasformare in isotopi stabili
radionuclide
emivita (t½)
tipo di
radiazione
3H
12 anni
14C
5715 anni
24Na
15 ore
32P
14 giorni
35S
87 giorni
40K
1.25∙109 anni
45Ca
163 giorni
125I
59 giorni
131I
8 giorni
16
Datazione al radiocarbonio
Il carbonio possiede tre isotopi: due stabili (12C e 13C) e uno
radioattivo (14C). Quest'ultimo si trasforma per decadimento
(elettroni) in azoto (14N), con un tempo di dimezzamento
medio (o emivita) di 5715 anni.
14
6
k
C
14
7
N
0
1
k
ln 2
t
1
2
0.693
1.2 10 4 anni 1
5715
La produzione di nuovo 14C avviene in natura negli strati alti
della troposfera e nella stratosfera, per la cattura di neutroni
termici da parte degli atomi di azoto. L'equilibrio dinamico che si
instaura tra produzione e decadimento radioattivo mantiene
quindi costante la concentrazione di 14C nell'atmosfera (dove è
presente principalmente legato all'ossigeno sotto forma di CO2).
17
Tutti gli organismi viventi che fanno parte del ciclo del C scambiano
continuamente C con l'atmosfera attraverso processi di respirazione (animali) o
fotosintesi (vegetali), oppure lo assimilano nutrendosi di altri esseri viventi o
sostanze organiche. Di conseguenza finché un organismo è vivo, il rapporto tra
la sua concentrazione di 14C e quella degli altri due isotopi si mantiene costante e
uguale a quella che si riscontra nell'atmosfera.
Dopo la morte, però, questi processi terminano e l'organismo non scambia più
C con l'esterno. Per effetto del decadimento, quindi, la concentrazione di 14C
diminuisce in modo regolare secondo la formula:
14
6
k
14
7
0
1
C
N
14
14
[ C] [ C]0 e
k t
k
ln 2
t
1
2
Quì, [14C]0 rappresenta la concentrazione di 14C al momento della morte
18
L’esame radiometrico del 14C sulla Sindone nel 1988 la datò fra
gli anni 1260 e 1390 (!?). Emivita del 14C = 5715 anni
La Sindone e [14C] = [14C]0 ekt
14
14
[ C] [ C]0 e
t MORTE
t
1
2
1
ln 2
t MORTE
t 2
[14 C]
ln 14
ln 2 [ C]0
19
Termodinamica e cinetica: le reazioni sono in genere reversibili
A+B
k1
k2
C
velocità della reazione diretta (vD)
L’equazione cinetica è: v k1 [A ] [B] k 2 [C]
velocità della reazione inversa (vI)
All’equilibrio vD = vI
e
K EQ
[C ]
k
1
[A] [B] k 2
Le reazioni dal punto di vista microscopico
Affinchè qualsiasi reazione chimica abbia luogo la teoria delle
collisioni stabilisce che devono essere soddisfatte 3 condizioni:
1. le molecole reagenti devono collidere fra loro
2. le molecole reagenti devono collidere con energia sufficiente
per rompere i legami coinvolti nella reazione
3. le molecole devono collidere secondo una orientazione che può
consentire il riarrangiamento degli atomi e la formazione dei
prodotti.
21
1. le molecole reagenti devono collidere fra loro
2. le molecole reagenti devono collidere con energia sufficiente
per rompere i legami coinvolti nella reazione
3. le molecole devono collidere secondo una orientazione che può
consentire il riarrangiamento degli atomi e la formazione dei
prodotti.
CO (g) + NO2 (g) → CO2 (g) + NO (g)
reagenti
avvicinamento dei reagenti
complesso
collisionale
riconoscimento
prodotti
separazione dei prodotti
22
1. le molecole reagenti devono collidere fra loro
2. le molecole reagenti devono collidere con energia sufficiente
per rompere i legami coinvolti nella reazione
3. le molecole devono collidere secondo una orientazione che può
consentire il riarrangiamento degli atomi e la formazione dei
prodotti.
distribuzione di Maxwell-Boltzmann
23
L’energia cinetica minina richiesta per la reazione è detta energia di
attivazione Ea (l’energia dello stato di transizione o complesso
attivato)
A+ B → C+ D
reazione esotermica
reazione endotermica
complesso attivato
Energia →
energia →
complesso attivato
avanzamento della reazione→
avanzamento della reazione→
avanzamento della reazione = coordianta di reazione
24
L'equazione di Arrhenius
L'equazione di Arrhenius mette in
relazione la temperatura con l'energia di
attivazione Ea (cioè la minima energia che
le molecole debbono possedere perché la
reazione proceda).
k A e
Ea
RT
Ea 1
ln k ln A
R T
dove:
k rappresenta la costante di velocità specifica
A il fattore di frequenza, che è correlato con la frequenza degli urti e
con la probabilità che essi si verifichino con orientamento favorevole
Ea l'energia di attivazione
R la costante universale dei gas
T la temperatura assoluta
25
1. le molecole reagenti devono collidere fra loro
2. le molecole reagenti devono collidere con energia sufficiente
per rompere i legami coinvolti nella reazione
3. le molecole devono collidere secondo una orientazione che può
consentire il riarrangiamento degli atomi e la formazione dei
prodotti.
urto produttivo
prima della collisione
collisione
urto non produttivo
dopo la collisione
26
Gli Enzimi
tripsina
sito attivo
27
Proprietà generali degli enzimi (e differenze rispetto ai catalizzatori
non biologici)
Velocità di reazioni più elevate: le velocità delle reazioni catalizzate dagli enzimi
sono normalmente da 106 a 1014 superiori rispetto a quelle delle corrispondenti
trasformazioni chimiche non catalizzate e almeno di diversi ordini di grandezza
superiori a quelle delle corrispondenti reazioni catalizzate per via chimica.
Condizioni di reazione più blande: le reazioni catalizzate dagli enzimi avvengono
in condizioni relativamente più moderate, a temperature inferiori a 100 °C e
pressione atmosferica e pH prossimo alla neutralità. Al contrario, la catalisi
chimica richiede sovente temperature e pressioni elevate e valori di pH estremi.
Maggiore specificità di reazione: gli enzimi esibiscono un grado specificità per
substrati e prodotti straordinariamente superiore rispetto ai catalizzatori chimici.
Possibilità di regolazione: le attività catalitiche di numerosi enzimi variano in
risposta alle concentrazioni di sostanze diverse dai loro substrati. I meccanismi
inerenti a tali processi regolatori comprendono il controllo allosterico, la
modificazione covalente degli enzimi, nonché la variazione della concentrazione
degli enzimi.
Velocità di reazioni più elevate: le velocità delle reazioni catalizzate dagli enzimi sono
normalmente da 106 a 1014 superiori rispetto a quelle delle corrispondenti trasformazioni
chimiche non catalizzate e almeno di diversi ordini di grandezza superiori a quelle delle
corrispondenti reazioni catalizzate per via chimica.
Tempo di dimezzamento:
t
1
t
2
1
2
ln 2
5.3 s
1
1.3 10
t
1
2
ln 2
k
reazione non enzimatica
ln 2
6 7 10 7 s 0.7 s reazione enzimatica
10
Condizioni di reazione più blande: le reazioni catalizzate dagli enzimi avvengono
in condizioni relativamente più moderate, a temperature inferiori a 100 °C e
pressione atmosferica e pH prossimo alla neutralità. Al contrario, la catalisi
chimica richiede sovente temperature e pressioni elevate e valori di pH estremi.
N2 + 3 H 2
2 NH3
In laboratorio: pressioni e temperature altissime
Rhizobium trifolii: condizioni ambientali
N2 + 8 H+ + 8 e + 16 ATP
2 NH3 + H2 + 16 ADP + 16 Pi
La nitrogenasi da
Klebsiella pneumoniae
Rhizobium trifolii
Maggiore specificità di reazione: gli enzimi esibiscono un grado specificità per
substrati e prodotti straordinariamente superiore rispetto ai catalizzatori chimici.
Un sito di legame per un substrato è costituito da una
fessura posta sulla superficie dell’enzima, la cui forma
è complementare al substrato (complementarietà
geometrica). I residui amminoacidici presenti nel sito
attivo sono disposti in modo da attrarre il substrato
mediante interazioni di van der Waals, elettrostatiche,
idrofobiche e legami idrogeno (complementarietà
elettronica).
La complementarietà
substrato/enzimi è il
fondamento del modello
chiave-serratura di Fischer
Emil Fischer
premio Nobel 1902
Sebbene i siti attivi della maggior parte degli enzimi siano in larga misura preformati,
essi vanno incontro ad un certo grado di modificazione conformazionale indotto dal
binding del substrato (adattamento indotto, induced fit)
La reazione esochinasica
NH2
ossidrile in 6
N
N
2+
Mg
OH
O
O
+
OH
HO
OH
O
-
-
O
P
O
-
O
P
O
O
-
P
O
N
N
O
ATP
O
O
OH
a-D-glucosio
OH
OH
esochinasi
NH2
O
O
P
O
O
O
OH
HO
OH
OH
a-D-glucosio-6-fosfato
N
N
-
2+
Mg
-
O
+
O
-
P
O
-
O
O
P
-
N
N
O
O
ADP
O
OH
OH
L’adattamento indotto rende conto anche della regiospecificità della reazione
esochinasica
ossidrile in 6 del
glucosio
Stereospecificità degli enzimi
Gli enzimi sono altamente specifici sia nel legame di substrati chirali, sia nel
catalizzare le loro reazioni.
Esempi:
• la tripsina idrolizza proteine con L-amminoacidi ma non con D-amminoacidi
• gli enzimi coinvolti nel metabolismo del glucosio sono specifici solo per il Dglucosio.
La cinetica enzimatica
Iniziò nel 1902 quando Adrian J. Brown studiò la reazione di idrolisi del saccarosio da parte
di un enzima di lievito -fruttofuranosidasi (invertasi) purificato:
OH
OH
O H
H
OH
H
OH
H
H
HO
OH
O
H
+
H
H2O
OH
O
OH
HO
HO
H
HO
H
H
OH
H
O
H
HO
H
H
OH
O H
HO
H
OH
OH
OH
a-D-glucosio
saccarosio
b-D-fruttosio
Egli scoprì che quando la [saccarosio] >> [enzima], nell’equazione cinetica la
velocità di reazione era di ordine zero rispetto alla [saccarosio].
v k [saccarosio]0
Reazioni di ordine zero (i.e. indipendenti dalla concentrazione del reagente) sono
dette anche cinetiche a saturazione. Brown propose che la reazione complessiva fosse
composta da 2 reazioni elementari, in cui il substrato prima si lega all’enzima ed in
seguito è decomposto nei prodotti, rigenerando l’enzima libero.
k1
E+S
k2
ES
k-1
E+P
37
Le due reazioni più comunemente trovate negli enzimi.
nella reazione
v
k
A
d[A ] d[B]
k [A]
dt
dt
nella reazione
A+B
B
l’equazione cinetica è:
ed è del primo ordine rispetto ad A. Le
dimensioni di k sono tempo1 (es. s1)
k
C
l’equazione cinetica è:
d[A ]
d[B] d[C]
v
k [A ] [B]
dt
dt
dt
il binding
ed è del primo ordine rispetto ad A e B, complessivamente del
secondo ordine. Le dimensioni di k sono M1∙tempo1 (es. M1s1)
38
L’equazione di Michaelis-Menten (Henri)
Leonor Michaelis
(1875-1940)
Maud Menten
(1879-1960)
Essa descrive la velocità di una reazione enzimatica in funzione
della concentrazione del substrato.
k1
E+S
k2
ES
k1
E+P
VMAX [S]
v0
K M [S]
39
k1
E+S
kCAT
ES
E+P
k1
Nel meccanismo E ed ES rappresentano l’enzima libero ed il complesso
enzima-substrato.
Ognuna delle reazioni elementari (qui 2) è descritta da una costante di
velocità specifica:
k1: costante di velocità per la formazione di ES (è una costante del secondo
ordine con dimensioni M-1s-1)
k-1: costante di velocità per la dissociazione di S da ES (è una costante del
primo ordine con dimensioni s-1)
k2: costante di velocità per la formazione di P da ES (è una costante del
primo ordine con dimensioni s-1)
Nota: per semplicità si assume che la reazione di formazione del prodotto P
sia irreversibile. Ciò deve essere giustificato da esperimento ed in genere gli
enzimi catalizzano reazioni reversibili.
VMAX [S] k CAT E 0 [S]
v
K M [S]
K M [S]
v
VMAX
[S]
KM
v VMAX
[S] = KM
41
VMAX [S] k CAT E 0 [S]
v
K M [S]
K M [S]
Valori di KM, kCAT e kCAT/KM per
alcuni enzimi e substrati
HO
CH3
O
O
O
NH
O
H3C
O
CH3
O
H3C
HN
CH3
CH3
O
NH
CH3
O
O
CH3
N-acetilglicina etilestere N-acetilvalina etilestere
N-acetiltirosina etilestere
42
Determinazione grafica dei parametri stazionari kCAT e KM
grafico dei doppi reciproci o di LineweaverBurk
• grafico di Eadie-Hofstee
43
Grafico di Lineweaver-Burk
44
Concetto importante: gli enzimi abbassano l’energia di
attivazione della reazione
reazione non
catalizzata
reazione
catalizzata
45
L’inibizione enzimatica
• proteasi a serina
• acetilcolinesterasi
diisopropilfluorofosfato
• La maggior parte dei farmaci sono inibitori enzimatici
• Lo studio della cinetica enzimatica è utile per la progettazione di nuovi farmaci
• Gli studi cinetici permottono di stabilire l’efficacia di un farmaco in vitro
• Le tossine naturali e molti agenti chimici bellici sono inibitori degli enzimi
46
L’inibizione enzimatica
Gli inibitori sono sostanze che legandosi all’enzima ne riducono l’attività,
influenzandone il legame con il substrato (KM) o il suo numero di turnover (kCAT).
Possono essere suddivisi in:
• inibitori irreversibili (o inattivatori): questi si legano così saldamente
all’enzima da bloccarne permanentemente l’attività
• inibitori reversibili: riducono l’attività legandosi reversibilmente
all’enzima. Sono suddivisi in:
1.
inibitori competitivi
2.
inibitori incompetitivi
3.
inibitori non competitivi (inibizione mista)
47
Inibizione competitiva
48
Gli inibitori competitivi
Si manifesta quando una sostanza strutturalmente simile al substrato si lega al sito
attivo dell’enzima. La sostanza compete con il substrato per occupare il sito attivo
La triosofosfato isomerasi catalizza l’isomerizzazione fra D-gliceraldeide-3-fosfato e
diidrossiaceton fosfato. Il glicerolo-3-fosfato è un inibitore competitivo
H
C
O
substrato
HO
O
O
-
O
-
P
O
H
H
C
HO
OH
O
O
-
inibitore
O
-
P
O
pdb: 1m7p
Inibizione incompetitiva
Per esempio il farmaco anti-HIV nevirapina
può legarsi a ed inibire la DNA polimerasi
virale soltanto dopo che questa si è legata al
proprio substrato . Quando la polimerasi si
lega al DNA cambia conformazione
generando una tasca idrofobica
(precedentemente non disponibile) a cui
l'inibitore può legarsi
50
Inibizione mista (non competitiva)
Alcuni inibitori si legano
covalentemente a residui
amminoacidici presenti
sull'enzima. Il binding del Pb2+ a
residui di cisteina
nell'enzima ferrochelatasi
inibisce l'attività di inserzione del
Fe2+ nella protoporfirina IX.
51