L’ORGANO DI RIPRODUZIONE DELLE ANGIOSPERME : IL FIORE
Una visione globale
La caratteristica peculiare delle Angiosperme, come abbiamo visto, è la
formazione del frutto nel quale tutto il corpo di un organo che abbiamo definito
ovario (in alcuni casi di tutto il fiore) si differenzia in modo tipico in ogni genere e
specie, protegge e promuove la dispersione del seme nell’ambiente. Dal quadro
evolutivo che la moderna sistematica oggi ci offre emerge che il frutto rappresenta
quindi una formidabile innovazione nell’ambito delle modalità di dispersione dei
semi e di conseguenza delle strategie adattative per l’affermazione della specie. Per
comprendere a pieno le ragioni evolutive che hanno portato alla formazione di tali
strutture occorre conoscere i processi attraverso i quali è possibile “manipolare” le
caratteristiche anatomiche, morfologiche e funzionali di un organismo.
La prima tappa per poter intraprendere questo percorso è quella di
individuare in un organismo la sede (l’organo o gli organi) nella quale questi
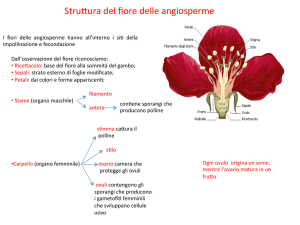
processi avvengono. Nelle Angiosperme questo organo è il fiore. Nel fiore si
distinguono soprattutto la parte riproduttiva femminile costituita dagli ovuli,
racchiusi in un involucro formato dai carpelli (ovario), e le sacche polliniche
formate dai tessuti dello stame che costituiscono la parte riproduttiva maschile;
inoltre c’è la presenza di altri elementi che ne costituiscono la parte sterile ed
assolvono ad altre funzioni quale quella protettiva e vessillare. Queste
caratteristiche peculiari del fiore sono direttamente in rapporto con le modalità con
cui avviene la riproduzione sessuata che ha come tappa iniziale il trasporto del
polline tramite diversi agenti (es. gli animali in tutte le Angiosperme primitive).
I granuli pollinici non vengono però a contatto diretto con gli ovuli, ma
sono appoggiati sullo stigma, una speciale struttura che si forma sui carpelli e che
svolge il ruolo attivo nel “riconoscimento” del polline: si tratta infatti meccanismi
di compatibilità istochimica che determinano l’accettazione o il rigetto del polline.
Solo un granulo pollinico che sarà risultato compatibile potrà produrre un budello
pollinico che dallo stigma trasporterà le cellule spermatiche all'ovulo ed avverrà la
fecondazione che consiste, nella sua fase culminante, nella fusione di una cellula
spermatica con la oosfera. Da questa fusione si origina una cellula che è detta
zigote dalla quale attraverso fenomeni di divisione e differenziazione cellulare si
originerà l'embrione.
Da questo processo emerge che la fecondazione nelle Angiosperme
è
completamente svincolata dalla presenza di acqua, cosa che invece risulta
indispensabile presso piante più primitive. Ciò ha garantito il successo evolutivo
delle Angiosperme accanto all'altro grado di differenziazione dei tessuti è e la
plasticita degli apparati vegetativi: infatti accanto alle forme biologiche legnose si
trovano forme erbacee piuriennali fino ad annuali.
Approfondimenti trasversali
Lo svincolamento delle piante dall’acqua. Modificazioni dei meccanismi della
riproduzione sessuata nel processo di colonizzazione dell’ambiente terrestre da
parte degli organismi vegetali.
IL FIORE DELLE ANGIOSPERME: UNA STRUTTURA RIPRODUTTIVA
INTEGRATA
ricettacolo
Dal punto di vista filogenetico il fiore della Angiosperme si è originato da un ramo
modificato e raccorciato le cui appendici fogliari si sono variamente trasformate
per assolvere i vari compiti inerenti la riproduzione
Nei fiori regolari ermafroditi (in cui sono presenti sia l’androceo che il gineceo)
che hanno più piani passanti per l'asse del fiore capaci di dividerlo in due metà
simmetriche, le varie parti del fiore sono disposte in cicli concentrici (verticilli) e
inseriti lungo un asse detto ricettacolo.
Il calice, che costituisce il ciclo più esterno, è formato da elementi fogliacei,
verdi e a funzione protettiva detti sepali. Internamente al calice abbiamo uno o più
cicli costituenti la corolla composta da petali spesso vivacemente colorati. Calice
e corolla, che non sono direttamente coinvolti nella riproduzione, svolgono
funzioni accessorie e costituiscono il perianzio. Esso è direttamente collegato con
l'impollinazione zoofila: prima della apertura del fiore il perianzio protegge gli
organi riproduttivi non ancora maturi dagli animali visitatori; durante la fioritura
invece talune parti dell'involucro fiorale sono vistosamente colorate e attirano
l'attenzione degli impollinatori.
Internamente al perianzio troviamo gli stami, il cui insieme costituisce
l'androceo (casa del maschio) poi uno o più pistilli, il cui insieme costituisce il
gineceo (casa della femmina).Gli stami e i
pistilli costituiscono gli organi
riproduttivi veri e propri del fiore.
Qualche notizia in più
La disposizione a verticillo domina nella gran parte delle Angiosperme: essa
favorisce la netta separazione degli elementi fiorali nei verticilli adiacenti e ne fissa
il numero (per lo più 5 ma anche 4, 3 e 2 nelle Dicotiledoni, quasi sempre 3 nelle
Monocotiledoni). Il numero dei verticilli varia per ogni fiore. Particolarmente
frequenti sono i fiori pentaciclici con 5 verticilli: 2 cicli dei perianzio (calice e
corolla), 2 cicli di stami, 1 ciclo di carpelli
Dal punto di vista evolutivo la disposizione spiralata e con numerosi stami è la
più primitiva (poliandria primaria).
Nel corso dell’evoluzione della struttura
fiorale si assiste alla riduzione del numero di stami e alla disposizione su un unico
verticillo; nelle Angiosperme comunque si assiste anche ad un ritorno ad un
elevato numero di di stami (poliandria secondaria), indicando che esso non in tutti i
casi è sinonimo di primitività. Forse la proliferazione secondaria degli stami è in
diretto rapporto con il cambiamento funzionale che si ha in seguito al passaggio
dalla impollinazione anemofila (ad opera del vento) a quella entomofila (per mezzo
di insetti). Questo fenomeno è rilevabile particolarmente nei fiori che offrono ai
visitatori una grande quantità di polline
Anche nel gineceo delle Angiosperme la disposizione a spirale di numerosi
carpelli liberi è la più primitiva . La diminuzione conduce anche qui ad una
disposizione verticillata di carpelli liberi (spesso 5, 3 o 2) ed in seguito porla alla
riduzione fino ad un unico carpello. La moltiplicazione secondaria del numero di
carpelli è più rara (più spesso viene compensata dalla riduzione del numero di semi
per carpello, per es. nelle Ranunculaceae). Spesso si giunge, con il concrescimento
di carpelli liberi alla formazione di un gineceo concresciuto o cenocarpo.
Nei fiori delle Angiosperme si trovano tutte le possibili forme di distribuzione dei
sessi .Fu discusso a lungo su quale fosse la struttura più primitiva, ma oggi prevale
l'opinione che i fiori ermafroditi (quelli contenenti sia la parte femminile che
maschile) siano più primitivi di quelli unisessuati per i seguenti motivi: 1) nei fiori
di quasi tutti i gruppi unisessuati si trovano rudimenti rispettivamente di ovari e di
stami (per es. in Castanea o Urtica); 2) le Angiosperme più primitive hanno in
maggioranza fiori ermafroditi (per es. Magnolia ecc.); questi soltanto facilitano il
trasferimento e la ricezione dei polline per mezzo di animali; 3) il passaggio da
fiori ermafroditi a unisessuati è spesso la conseguenza della selezione avvenuta per
il cambio dall'impollinazine zoofila a quella anemofila.
STAMI. Ciascuno stame è formato da un filamento e da una porzione terminale
rigonfia: l'antera. L'antera è costituita da due lobi, le sacche polliniche, ciascuna
formata da due cavità.. La sezione trasversale di una giovane antera per ogni sacca
pollinica mostra internamente una divisione in due cavità all’interno delle quali
verrà prodotto il polline. Queste cavità sono delimitate esternamente da una parete
costituita da almeno 4 strati. Questa parete, partendo dall'esterno è costituita da
un'epidermide (esotecio) , da uno strato fibroso (endotecio), da uno strato
intermedio e da un tessuto uni- o bistratificato detto tappeto. Quest'ultimo ha la
funzione di nutrire i granuli pollinici durante la loro maturazione all’interno delle
sacche polliniche.
Come avviene la produzione di polline
All’interno delle cavità delle sacche polliniche si forma un grande numero di
cellule madri del polline che sono diploidi (appendice 1a) da ciascuna delle quali
si formano attraverso il processo della meiosi (appendice 1b) 4 microspore aploidi
che restano attaccate costituendo una tetrade (appendice 1c). Ciascuna di queste
microspore è costituita quindi da un nucleo, dal citoplasma e da una parete
cellulare. Mentre sono ancora nelle sacche polliniche le microspore incominciano a
subire delle modificazioni: la microspora uninucleata si divide per mitosi
(appendice 2) in modo molto ineguale in una grande cellula vegetativa
uninucleata, che occupa quasi interamente il granulo e in una più piccola cellula
generativa, posta vicino alla parete cellulare. In una seconda divisione mitotica
che interessa solo la cellula generativa si ha la
formazione di due cellule
spermatiche. A questo stadio di maturazione (una cellula vegetativa uninuclata e
due cellule spermatiche uninucleate) la microspora diventa granulo di polline o
gametofito maschile (organismo da cui si origineranno i gameti)
Contemporaneamente avvengono delle modificazioni anche della parete del polline
che a questo stadio di maturazione produce una sostanza detta sporopollenina che
forma uno strato esterno alla parete denominato esina che può essere
pluristratificato e terminare internamente con uno strato cellulosico detto intina
anche esso pluristratificato. Sulla superficie dei granuli pollinici, si notano delle
aperture definite pori germinativi, che serviranno al momento della germinazione
del polline. Inoltre soprattutto nelle piante con impollinazione zoofila, si trova un
cemento pollinico costituito in parte da sostanze oleose che favorisce
l'aggregazione dei granuli pollinici in gruppi e la loro adesione all'anìmale
impollinatore; inoltre impedisce la prematura caduta del polline dall'antera aperta.
La ragione biologica per cui la pianta ha dotato il granulo pollinico di tegumenti
risiede nel fatto che esso rappresenta un organo di resistenza che una volta
abbandonata la pianta madre deve difendersi dal pericolo di disidratazione.
Qualche notizia in più
L’esina contribuisce a fornire una peculiare microstruttura dei granuli pollinici
che nelle Angiosperme raggiunge un più alto grado di differenziazione e riveste un
importante ruolo tassonomico. Infatti in considerazione dell'enorme varietà di tipi, i
granuli pollinici possono essere ordinati e classificati solo con l'aiuto di un sistema
artificiale (sistema N P C) che si basa sul numero (N), la posizione (P) ed il
carattere (C) dei pori germinativi. Inoltre vi sono molte differenze di importanza
tassonomica che riguardano la simmetria', la forma e le dimensioni dei granuli
pollinici e la microstruttura della loro esina.
Avvenuta la maturazione dei granuli pollinici le sacche polliniche si aprono per
mezzo di un meccanismo di coesione. Quando le cellule dei tessuti che
costituiscono l’antera perdono acqua si originano delle tensioni che provocano la
rottura della parete dell'antera, per lo più in senso longitudinale, in prossimità della
parete divisoria che separa le coppie di sacche polliniche, rilasciando il polline
all’esterno.
IL PISTILLO
Costituisce la parte femminile del fiore in quanto esso è
costituito da «foglie» del fiore più o meno trasformate avvolte e saldatesi se stesse
portanti gli ovuli prende nome di carpello. E’ formato da una porzione basale
rigonfia, l'ovario, sovrastata da una parte terminale di tipo peduncolare, lo stilo,
che rimane sterile; e nel suo interno passano e vengono nutriti i budelli pollinici.
Lo stilo porta uno s t i g m a, la zona recettrice del polline in quanto coperto per lo
più da papille o da muciliagine vischiosa capace di trattenere i granuli pollinici. Il
pistillo può essere formato da un solo carpello (pistillo semplice), oppure come
spesso avviene, essere formato da più carpelli saldati assieme ( pistillo composto o
cenocarpico). Il loculo è la cavità interna (o le cavità interne) dell'ovario dove sono
sistemati gli ovuli. Spesso nei pistilli composti il numero dei loculi corrisponde al
numero dei carpelli componenti (come abbiamo avuto modo di vedere anche nei
frutti). A volte le porzioni interne dei carpelli mancano più o meno completamente
e il pistillo composto da una singola cavità (monoloculare). Vi possono essere altri
elementi della morfologia esterna del pistillo che rivelano il numero dei carpelli
costituenti: l'ovario può, ad es., presentare un numero corrispondente di costole (es:
ovario di Iris, ovario delle leguminose)
In genene l'ovario contiene uno o più ovuli. Abbiamo già visto come l'ovulo sia
attaccato alla parete interna dell'ovario mediante il sospensore. La zona di attacco
viene detta placenta e la disposizione degli ovuli all'interno dell'ovario
placentazione. La placentazione è una caratteristica tassonomica importante.
Quando in un ovario composto gli ovuli sono inseriti in file longitudinali sulla
parete interna, abbiamo una placentazione parietale. Quando gli ovuli di un
ovario composto pluriloculare sono inseriti su di una colonna in posizione assile,
abbiamo la placentazione assile. Le progressive modificazioni della placentazione
dovrebbero essere dirette al miglioramento della nutrizione degli ovuli e dei semi.
La struttura del pistillo quindi consente in tal modo agli ovuli ancora delicati di
essere protetti dagli insetti e dal disseccamento, mentre è controllato l'accesso
diretto del polline mediante lo stilo e lo stimma.
Ovuli.
Gli ovuli vengono prodotti dalla placenta e che li rifornisce di sostanze
nutritive attraverso fasci conduttori. L’ovulo è costituito da tegumenti (in genere
due) e da un tessuto interno che è detto nucella. I tegumenti si interrompono in un
punto che è detto micropilo da dove il tubetto pollinico penetrerà nell’ovulo.
Analogamente a quanto è avvenuto nella linea maschile anche nell’ovulo devono
avvenire dei processi alla fine dei quali si deve avere la produzione del gamete
femminile. Entriamo nei particolari; una sola cellula della nucella si divide per
meiosi e forma 4 macrospore aploidi disposte in tetrade lineare. Tre di queste
degenerano e una sola di queste va incontro a quattro divisioni mitotiche formando
così 8 cellule. Queste cellule si orientano in modo peculiare all’interno dell’ovulo
in modo che il gamete femminile (oosfera) e due cellule compagne (sinergidi)
siano poste dal lato del micropilo, tre cellule (antipodi) siano poste sul lato opposto
al micropilo e le restanti due siano poste al centro dell’ovulo dove si fondono
formando una cellula diploide. L’ovulo al termine di questo stadio di sviluppo si è
trasformato in gametofito femminile.
Tavola 2 Dissezione del fiore nei suoi componenti
Tavola 3. Qualche esempio di struttura fiorale
Impollinazione. Nelle Angiosperme il polline è catturato dallo stigma adesivo e
papilloso; ciò comporta un'accentuazione dell'indipendenza dall'umidità ambientale
della riproduzione sessuale.
La formazione degli stigmi è conseguente alla
presenza degli ovuli nei carpelli ed è un dispositivo di protezione associato con la
impollinazione zoofila e anche con il corrispondente ermafroditismo del fiore.
Tutto ciò indica che le Angiosperme primitive erano ermafrodite e zoofile e che i
vantaggi derivantì da questa loro condizione rispetto alle loro progenitrici
unisessuali e anemofile, ne favorirono la diffusione.
A seconda degli agenti esterni che trasportano il polline possiamo distinguere le
Spermatofite in anemofile, zoofile e idrofile:
L'impollinazione per mezzo di animali (zoofilia = zoogamia) presuppone che gli
animali impollinatori visitino i fiori regolarmente e si trattengano in essi per un
tempo sufficientemente lungo, affinchè i fiori possano sostenere la sollecitazione
meccanica provocata dai pronubi, che il polline e lo stigma vengano toccati
regolarmente e il polline aderisca sufficientemente agli impollinatori e giunga con
sufficiente sicurezza sullo stigma di altri fiori. 1 fiori che devono essere visitati
dagli animali impollinatori dispongono quindi di esche (polline, nettare ecc.), di
stimoli (colore, profumo ecc.) e di pollini adesivi
Nel
corso
dell'evoluzione
delle
Angiosperme
si
verificò
un'importante
differenziazione dei meccanismi di richiamo oltre che alle modificazioni strutturali
dei fiere e vi fu un aumento progressivo dei gruppi di animali coinvolti nel
processo dell'impollinazione, soprattutto insetti e diversi gruppi di uccelli. Da
casuali visite a fiori da parte di animali diversi si svilupparono gradualmente
rapporti di reciproco vantaggio tra fiori particolarmente specializzati «in senso
zoofilo» e animali «antofili». Il perfezionamento dei meccanismi di attrazione di
particolari impollinatori rende possibile alla pianta una impollinazione più sicura
con una quantità minore di polline e con una migliore produzione di semi. Infatti
mentre il rapporto granuli pollinici-ovuli nelle piante anemofile è nell'ordine di
grandezza di 106: 1, nelle piante entomofile specializzate (come per es. le
Orchidee) può essere solo ca.
1:1
La concorrenza tra animali impollinatori
specializzati e generici «antofili» andò progressivamente riducendosi e
l'impollinazione diretta, svolta dai primi sulle «loro» piante da cui traggono
nutrimento, torna a loro vantaggio. L'evoluzione filogenetica delle Angiosperme
zoofile e dei gruppi di animali impollinatori ad esse adattati è da intendersi come
una «co-evoluzione» reciprocamente determinata. L'adattamento reciproco dei
partner è a volte tanto avanzato che ciascuno di essi non potrebbe vivere senza
l'altro.
Qualche notizia in più
Il mezzo originario di attrazione dei fior delle Angiosperme fu senza dubbio il
nutrimento, cioè l'abbondanza di polline che contiene proteine, grassi, carboidrati e
vitamine. Tali fiori da polline, accessibili anche agli insetti primitivi con apparato
masticatore si trovano nelle Magnoliidae (per es. Anemone, Papaver, Victoria),
nelle Rosidae (per es. Rosa) e nelle Dilleniidae primitive (per es. Paeonia). Lo
stadio successivo, ma ancora primitivo, fu caratterizzato dall'offerta agli
impollinatori di tessuti nutritivi specialmente di succhi zuccherini (nettare). Ciò
rende possibile un risparmio sulla produzione già elevata di polline. La stragrande
maggioranza delle Angiosperme zoofile attuali ha fiori nettariferi. La disponibilità
di nettare ha indotto un miglioramento degli apparati succiatori degli impollinatori.
I nettarii sono generalmente prodotti dal «disco» dell'asse fiorale, da stami
trasformati, ma anche da determinate zone di carpelli, di petali e di sepali. In
condizioni primitive il nettare era libero sui fiori, quindi accessibile anche agli
impollinatori con apparato succiatore corto, come per es. sui carpelli di Magnolia o
sul ricettacolo di molte Rosaceae; successivamente venne raccolto in profondità e
accumulato in speciali contenitori come per es. negli speroni cavi di Viola, o nei
lunghi tubi dove è accessibile solo per certi animali, per es. i Lepidottori, con
proboscide lunga.
Alcune Angiosperme (per es. Calceolaria) offrono come nutrimento agli
impollinatori specializzati un olio (lipidi). In alcuni casi rari, da parte dei fiori può
anche essere sfruttato l'istinto di procreazione di particolari animali.
La tignola Tegiticula yuccasella impollina i fiori della agavacea Yucca e deposita
poi le sue uova nell'ovario; le larve si nutrono con una parte dei semi che si stanno
sviluppando. I fiori dell'orchidea mediterranea simulano nella forma, pelurie e
profumo delle femmine di determinate api o vespe, ed inducono i maschi di queste
specie a tentativi di accoppiamento con effetti positivi per l'impollinazione.
Quest'ultimo esempio appartiene alla categoria dei fiori che con un «inganno»
ottengono la visita di animali senza dare niente in cambio; tra questi sono anche da
ricordare i fiori-trappola
I mezzi di richiamo dei fiori delle Angiosperme sono principalmente ottici e
chimici e non di rado operano contemporaneamente, ma il loro effetto può essere
diverso da vicino o da lontano.
La comprensione esatta dell'efficacia ottica e chimica dei fiori presuppone una
sicura conoscenza della fisiologia sensoriale degli animali impollinatori, che finora
si ha solo per alcuni come l'ape, i bombi e alcuni colibrí.
Per quanto è stato accertato sul senso dei colore di questi animali sappiamo che
l'ape ed i bombi, per es. non percepiscono il rosso puro, mentre percepiscono
l'ultravioletto da 400 a 310 nm invisibile all'uomo, e che tra gli altri colori fiorali
percepiscono solo una banda di giallo da 650-520 nm, una banda di blu-violetto da
490-400 nin, ed il bianco che viene percepito come verde azzurro. Invece la
percezione ottica degli uccelli è molto simile a quella dell'uomo ed essi sono
particolarmente sensibili al rosso.
L'attrazione chimica dei fiori è fondata soprattutto sulla produzione di profumi
emanati dal polline, dai petali o anche da altri organi fiorali. Alcuni di questi
profumi possono essere sgradevoli per l'uomo come nel caso particolare delle
Araceae che emanano odore di cadavere e di escrementi e che vengono impollinati
da insetti necrofagi e coprofagi. Le emanazioni irregolari dei profumi determinano
un avvicinamento degli animali impollinatori irregolarmente e con minore
sicurezza di quanto avviene nel caso dello stimolo ottico cui corrisponde una
risposta diretta. Per le api e i bombi l'odore è importante su brevi distanze. Molti
fiori posseggono un «odore guida» analogamente al «colore guida», talvolta
ambedue localizzati nella stessa parte dei fiore (per es. nella corolla accessoria di
Narcissus).
Lo sviluppo di questi meccanismi di stimolo è parallelo al continuo miglioramento
degli organi sensori degli impollinatori. Le ripetute visite coronate da successo a
determinati fiori provocano nelle api e in altri imenotteri una «affinità» che le
induce temporaneamente ad un'attività di raccolta più intensa e legata a quei dati
fiori. Ciò dipende sia dall'efficacia stimolante degli odori specifici dei petali, e
della quantità di nettare e di polline raccolti, sia dalla «memoria» altamente
sviluppata di questi animali e dalla loro capacità di comunicare con un proprio
«linguaggio».
Gli adattamenti meccanici dei fiori rendono possibile l'impollinazine solo ad
animali forniti di una particolare struttura morfologica, essendo questi guidati per
determinate vie che garantiscono un adeguato contatto con il polline e lo stigma.
Può essere imposta anche una certa durata del soggiorno, per assicurare
l'impollinazione oppure il trasporto dei polline può essere garantito da determinati
meccanismi a leva, di adesione, di agganciamento e di proiezione.
Particolarmente vari e numerosi sono i f i o r i v isitati da api (melitofíli). Il loro
stile è caratterizzato da fiori con parti adatte per l'appoggio degli insetti, con colori
gialli, violetti e blu, e con profumi leggeri, con macchie guida per il nettare che è
riparato a media profondità (per es. Salvia)
I fiori impollinati da uccelli (ornitofili) si distinguono da quelli entomofili per il
loro stile molto particolare. Mancano zone di appoggio in quanto gli uccelli,
essendo troppo pesanti, devono svolgere le loro visite rimanendo sospesi in aria
(colibrì) o posandosi in un punto più stabile dei fiore. I fiori ornitofili sono grandi
e generalmente di tipo tubuliformi o a spazzola; i colori, vivaci e contrastanti, sono
soprattutto il rosso, ma anche il blu, il giallo e perfino il verde «<colori dei
pappagalli»); non hanno profumi poichè non potrebbero essere percepiti
dall'odorato poco sviluppato degli uccelli, il nettare invece è presente in buona
quantità e in forma piuttosto liquida nelle zone più profonde dove può essere
raggiunto solo da lingue allungate e a forma di pennello. Alla maggiore richiesta
di sostanze liquide da parte degli uccelli nelle regioni più calde, e alla presenza di
piante che fioriscono per gran parte dell'anno è da collegare la prevalenza della
ornitofílía in quelle zone. Il polline viene trasportato con il becco o tramite altre
parti del capo dove rimane attaccato.1 fiori ornitofili si trovano in quasi tutte le
famiglie zoofile dei Tropici
1 fiori impollinati dai pipistrelli (chiropterofili) sono limitati alla zona tropicale e
vengono ricercati da pteropi e vampiri.
Il loro stile è caratterizzato da una
posizione ben esposta, per lo più caliciformi, con fauce ampia, a forma di spazzola,
con antesi notturna, spesso con colori tetri, odore pungente di frutti o di
fermentazione e molto nettare e polline. Come esempi si possono citare Carnegiea
(Cactaceae), Adansonia (Bombacaceae).
Cobaea (Polemoniaceae, alcune
Bignoniaceae e specie dei generi Musa e Agave.
L'anemofilia (= anemogamia) comporta una produzione adeguata e una buona
distribuzione dei granuli pollinici e una loro rapida e uniforme dispersione. Inoltre
i granuli pollinici devono essere tali da mantenersi sospesi nell'aria per un periodo
abbastanza lungo e gli stigmi devono essere grossi e in buona esposizione perchè
intercettino il polline con frequenze elevate. Nel corso dell'evoluzione delle
Angiosperme si sono sviluppate stirpi con anemofilia secondaria in tempi e in
gruppi diversi proprio in luoghi bene esposti al vento, poveri in animali e con
aggruppamenti vegetali tendenzialmente puri, condizioni favorevoli a questo tipo
di impollinazione. In quasi tutti i gruppi delle Angiosperme anemofile si trovano
vestigia di un primitivo errnafroditisrno e di una precedente zoofilia.
I granuli pollinici sono privi di sostanze appiccicose, quindi secchi e lisci, si
separano con facilità e grazie alla loro forma minuscola rimangono a lungo in
sospensione nell'aria. La loro elevata quantità è realizzata da un forte aumento dei
fiori (circa 2 milioni e mezzo di granuli pollinici contro un ovulo). La caduta del
polline viene favorita dalla mobilità dei filamenti (per es. nelle Graminaceae). dei
peduncoli fiorali (per es. in Cannabis) oppure dell'intero asse dell'infiorescenza
(per es. negli amenti di Quercus); il polline viene in un primo tempo deposto e poi
trasportato dal vento.
L'impollinazione per mezzo dell'acqua, (idrofilia = idrogamia), è riscontrabile in
poche Angiosperme. Anche tra le piante acquatiche l'idrofilia non è molto diffusa e
spesso i loro fiori emergenti dalla superficie dell'acqua sono impollinati dal vento .
In alcune specie i fiori maschili distaccati, raggiungono gli stigmi tenuti fuori
dall'acqua per breve tempo.
Fecondazione. Gli eventi notevoli del ciclo biologico di una angiosperma
avvengono tutti nel fiore.
Il fiore è una struttura nella quale alloggia
permanentemente il gametofito femminile, ove inizia e termina l'esistenza il
gametofito maschile, nel fiore si realizza la fecondazione e si forma l’embrione.
Passiamo ai particolari. Il granulo pollinico deriva dalla microspora e contiene a
maturità tre cellule: una cellula somatica molto attiva, la cellula del tubetto
pollinico e due cellule spermatiche che sono i gameti maschili. Quando il granulo
arriva sullo stigma la cellula del tubetto si accresce, esce da uno dei pori del
granulo, produce un lungo filamento che, digerendo i tessuti dello stilo, entra nelle
cavità dell'ovario, penetra nell'ovulo attraverso il micropilo e stabilisce il contatto
col gametofito femminile. Una volta stabilitosi il contatto tra tubetto e ovulo, le
due cellule spermatiche dispongono di una galleria protoplasmatica di cui si
servono per entrare nell’ovulo ed effettuare la doppia fecondazione tipica delle
angiosperme. Ossia un nucleo spermatico si unisce con l’oosfera operando così la
fecondazione con la formazione di una cellula diploide che è detta zigote. Il
secondo nucleo spermatico si unisce alla cellula centrale presente nell’ovulo
formando una cellula triploide. Questo processo è detto doppia fecondazione
perché in seguito a ciò dall'oosfera si forma l'embrione e dalla cellula del centrale
l'endosperma. Contemporaneamente l'ovulo diventa seme e l'ovario frutto. Le
rimanenti parti fiorali, terminata la loro funzione, generalmente avvizziscono e
cadono (sfioritura).
Formazione dell'embrione, dell'endosperma e del seme. Dopo la fecondazione,
dallo zigote si forma un embrione; dal nucleo dell'endosperma e dal plasma residuo
dei sacco embrionale invece si forma un tessuto nutritivo, denominato endosperma
secondario. Lo zigote si divide per mezzo di uno o spesso più setti trasversali in
una corta fila di cellule denominata proembrione. Soltanto la prima cellula o cellule
di questa fila, rivolte verso il sacco embrionale, formano più tardi l'embrione, la cui
formazione inizia con la comparsa di pareti longitudinali. Le cellule rimanenti
diventano il sospensore. Esse spingono l'embrione entro il tessuto nutritivo che si
sta sviluppando e probabilmente conducono sostanze nutritive all'embrione.
Spesso esse confinano con l'embrione tramite una cellula chiamata «ipofisi», che
dopo ulteriori suddivisioni può partecipare alla formazione della caliptra e
dell'apice radicale. L'embrione all'inizio consiste in una struttura pluricelluiare,
globosa, divisa in quadranti e poi in ottanti, situata sulla parte terminale del
sospensore. Più tardi si differenzia in un primordio della radice con la cuffia
radicale (caliptra) rivolto verso il micropilo mentre dalla altra parte si formano i
cotiledoni e il meristema apicale del germoglio (plumula). Nelle Dicotiledoni si
formano 2 cotiledoni laterali, tra i quali sorge il meristema apicale. Nelle
Monocotiledoni si forma un solo cotiledone terminale, mentre il meristema apicale
è situato lateralmente
Nel seme è anche presente l’endosperma. Generalmente ancora prima della
divisione dello zigote, il nucleo endospermico si suddivide, iniziando la
formazione
dell'endosperma
secondario.
Questo
nutre
l'embrione,
ma
successivamente viene trasformato in un tessuto di riserva del seme, che
l’embrione utilizza prima e durante la germinazione.
La formazione dell'embrione e dell'endosperma richiede un abbondante
rifornimento di sostanze nutritive. L'endosperma secondario (di solito triploide)
delle Angiosperme si forma solo dopo la (doppia!) fecondazione. Ciò comporta 2
vantaggi: 1) lo sviluppo dei gametofito viene ulteriormente abbreviato e accelerato
e 2) il tessuto nutritivo si costituisce dopo l'avvenuta formazione dello zigote, cioè
quando il nutrimento dell'embrione è veramente necessario (economia! ).3)
essendo triploide il tessuto ha la possibilità di produrre un maggior quantitativo di
sostanze di riserva.
Approfondimento trasversale: modulo di Didattica della Chimica
Il seme e le sostanze di riserva: le principali molecole organiche che ritroviamo
come tipiche sostanze di riserva
Tavola 4. Alcuni tipologie di foglie. Identificane le peculiarità