Introduzione al metabolismo
1
2
Funzioni del metabolismo
1- Ottenere energia chimica
2- Convertire le molecole
3-Polimerizzare i precursori monomerici
4- Sintetizzare e degradare le biomolecole
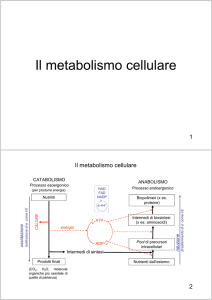
Il metabolismo è tradizionalmente
diviso in due parti:
1) Il catabolismo, o degradazione
2) L’anabolismo, o biosintesi
Catabolismo:
Sostanze nutrienti
CO2 + H2O + Energia utilizzabile
Anabolismo:
Energia utilizzabile + piccole molecole
Molecole complesse
In base all’agente ossidante utilizzato:
1. Aerobi obbligati
2. Anaerobi
3. Aerobi facoltativi
4. Anaerobi obbligati
6
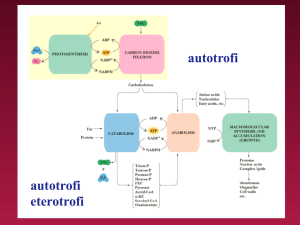
Strategie trofiche
In base alla fonte di energia libera:
1. Autotrofi (in grado di sintetizzare tutti i loro costituenti a partire
da molecole più semplici come H2O, CO2)
chemiolitotrofi ( attraverso l’ossidazione di composti inorganici
come NH3, H2S )
fotoautotrofi ( mediante fotosintesi )
2. Eterotrofi ( ottengono energia dall’ossidazione di composti organici
come carboidrati, lipidi e proteine )
7
Ruoli dell’ATP e del NADP+ nel metabolismo
(acetil CoA)
L’ATP e il NADPH sintetizzati mediante la degradazione
di metaboliti complessi sono la fonte di energia libera
per le reazioni di biosintesi e per altre reazioni.
SGUARDO GENERALE AL CATABOLISMO
Nella cellula le vie metaboliche hanno localizzazioni
specifiche.
Specializzazione dei tessuti:
ISOENZIMI
Catalizzano la stessa reazione
Sono codificati da geni diversi
Hanno proprietà cinetiche e regolatorie differenti
LDH tipo M e tipo H
COO-
NADH + H+
COO-
NAD+
HO
O
CH3
Acido piruvico
lattico deidrogenasi
H
CH3
Acido lattico
11
Energia, vita e
prima legge della termodinamica
lavoro meccanico, che muove una cellula, un organello
o un qualsiasi oggetto contro una forza gravitazionale
o frizionale.
lavoro di gradiente, che crea gradienti di
concentrazione e gradienti elettrici attraverso una
membrana.
lavoro di, sintesi che contempla la formazione e la
rottura di legami e la produzione di nuovi composti.
12
Prima legge della termodinamica
Durante una modificazione chimica o fisica,
l'energia non viene né creata né distrutta; la
quantità totale di energia resta costante e
può essere convertita in forme diverse.
13
Energia libera ed equilibrio
L'energia libera di un composto viene definita
come la variazione di energia che è associata alla
formazione di quel composto a partire da
elementi puri.
La variazione di energia libera (DG) di una
reazione chimica corrisponde all'energia libera
posseduta dai prodotti (G, prodotti) sottratta
l'energia libera dei reagenti (G, reagenti).
DG = G prodotti – G reagenti
14
Quando il sistema si modifica verso
uno stato che possiede meno energia
(G prodotti inferiore a G reagenti),
∆G ha valore negativo e la reazione è
detta esoergonica perché libera
energia.
Questa reazione procede
spontaneamente
Quando invece è necessario fornire energia
per fare avvenire una reazione che
altrimenti non avverrebbe,
∆G ha valore positivo e la reazione è detta
endoergonica.
Questa reazione non procede
spontaneamente.
16
I processi spontanei hanno ∆G negativo e
sono detti
esoergonici
I processi non spontanei hanno ∆G positivo
e sono detti
endoergonici
18
Energia libera ed equilibrio
All'energia libera (abbreviata con G, che sta per
energia libera di Gibbs) di qualsiasi elemento puro
nella sua forma più stabile alla temperatura di 298 K
(25°C) e alla pressione di 101,3 kPa (1 atm) viene
assegnato il valore O.
L'energia libera di un composto viene definita come la
variazione di energia che è associata alla formazione
di quel composto a partire da elementi puri.
19
La variazione di energia libera (∆
∆G) di una reazione chimica
corrisponde all'energia libera posseduta dai prodotti (G,
prodotti) sottratta dell'energia libera dei reagenti (G, reagenti):
∆G = G prodotti – G reagenti
Il ∆G di una reazione è una misura della quantità teorica
massima di lavoro che una reazione può generare a temperatura
e pressione costanti.
La maggior parte
dei lavori meccanici
dei lavori di gradiente
del lavoro di sintesi
che vengono prodotti dagli organismi viventi sono
generati dalle reazioni chimiche, in cui i prodotti
possiedono una quantità di energia libera
sostanzialmente più bassa di quella dei reagenti, e
quindi il valore di ∆G risulta significativamente
grande e negativo.
Considerazioni termodinamiche
Il ∆G di una reazione può essere calcolato
A+B C+D
∆G = ∆G° + RT In [C][D]
[A] [B]
∆G= variazione dell’energia libera
∆G°= variazione dell’energia libera standard
R = costante dei gas (8,315 J/mole.K)
T = temperatura assoluta (K)
22
Le misure termodinamiche sono basate sulla definizione di
uno stato standard in cui le concentrazioni dei reagenti e
dei prodotti sono fissate a 1 Molare .
Le energie libere standard ∆G° sono calcolate per reazioni
che avvengono a 25°C.
Per le reazioni che avvengono nei sistemi biologici il
cambiamento di energia libera ∆G°’ è misurato a pH 7.0
23
Termine costante
∆ G è correlata
che dipende soltanto dal tipo di reazione
che ha luogo.
Termine variabile
che dipende dalla concentrazione dei
reagenti e dei prodotti, dalla stechiometria
della reazione e dalla temperatura.
24
Il concetto di equilibrio
aA+bB cC+dD
La composizione di un sistema che sta reagendo
tende a cambiare continuamente fino a che non
viene raggiunto l’equilibrio.
Keq = [C]C [D]d
_______
[A]a [B]b
25
Quando i reagenti sono presenti a concentrazioni vicine
ai valori caratteristici della situazione all’equilibrio:
[C][D]/[A][B]= Keq
∆G=0
REAZIONI VICINE ALL’EQUILIBRIO
A+B C+D
Poiché ∆G è vicino a zero è possibile cambiare la direzione della
reazione modificando il rapporto dei prodotti rispetto ai reagenti.
26
REAZIONI VICINE ALL’EQUILIBRIO
A+B C+D
Gli enzimi che catalizzano queste reazioni tendono ad
agire velocemente per ripristinare le concentrazioni
vicine all’equilibrio.
La velocità della reazione è regolata dalle concentrazioni
dei reagenti e dei prodotti
27
REAZIONI IRREVERSIBILI
reazioni metaboliche che funzionano in
condizioni molto lontane dall’equilibrio.
L’enzima che catalizza queste reazioni ha
un’attività insufficiente per consentire il
raggiungimento dell’equilibrio.
28
REAZIONI IRREVERSIBILI
A+B
C+D
[C][D]/[A][B]=inferiore a zero
e DG negativo
29
In una via metabolica la maggior parte degli enzimi
operano in condizioni vicine all’equilibrio,
La velocità della reazione varia in funzione della
concentrazione dei substrati.
Alcuni enzimi che operano in condizioni lontane
dall’equilibrio sono localizzati, nelle vie metaboliche, in
posizione startegica.
30
Reazione irreversibile
La molecola del glucosio
induce un cambiamento
conformazionale
nell’esochinasi.
Esochinasi
II tappa di utilizzo dell’ATP (reazione irreversibile)
Fosfofruttochinasi
Svolge un ruolo centrale nel controllo della glicolisi
Controllo del flusso metabolico
Il flusso degli intermedi lungo una via metabolica in uno
stato stazionario è costante ed è determinato dalla
tappa che limita la velocità dell’intera via.
1. È la reazione più lenta
2. Il suo prodotto viene rimosso
3. Funziona in una condizione lontana dall’equilibrio
4. Possiede ∆G negativo
33
1. Le vie metaboliche sono irreversibili. Una reazione fortemente
esoergonica (una reazione con un ∆G < O) è irreversibile, cioè
raggiunge il completamento. Se una reazione di questo genere è
parte di una via metabolica costituita da più tappe, essa
conferisce una direzione precisa a tutta la via; rende cioè
irreversibile l'intera via metabolica.
2. Ogni via metabolica ha una prima tappa di comando. Anche se la
maggior parte delle reazioni di una via metabolica operano in
condizioni vicine all'equilibrio, di solito vi è una reazione
irreversibile (esoergonica) nella prima parte della via metabolica
che «comanda» al suo prodotto di continuare lungo la via, (così
come l'acqua che ha superato una diga non può tornare indietro).
3. Le vie cataboliche e le vie anaboliche sono differenti. Se un
metabolita viene convertito in un altro metabolita mediante un
processo esoergonico, è necessario fornire energia libera per
convertire il secondo metabolita indietro nel primo. Questo
processo energeticamente «in salita» richiede una diversa via per
almeno alcune delle tappe di reazione.
L'esistenza di vie di interconversione indipendenti è
un'importante proprietà delle vie metaboliche
perché consente il controllo indipendente dei due
processi.
Se la cellula richiede
«spegnere» la via che
tempo «accendere» la
il metabolita 2, è necessario
porta da 2 a 1 e nello stesso
via da 1 a 2.
A
2
1
X
Y
Questo tipo di controllo indipendente non sarebbe
possibile se non fossero presenti vie metaboliche
diverse.
Controllo del flusso metabolico
Il flusso degli intermedi lungo una via metabolica in
uno stato stazionario è più o meno costante: la
velocità di sintesi e di demolizione di ogni intermedio
in ogni via metabolica ne mantiene costante la
concentrazione.
Il flusso attraverso una via che si trova nello stato
stazionario è costante e viene determinato dalla tappa
(o dalle tappe) che determina la velocità dell'intera
via.
La tappa
funziona
possiede
libera.
che limita la velocità di una via metabolica
in una condizione lontana dall'equilibrio e
una grande variazione negativa di energia
Alcuni meccanismi possono controllare il flusso
attraverso la reazione che limita la velocità di una via:
Controllo allosterico
Modificazione covalente (interconversione
enzimatica)
Cicli del substrato
Controllo genetico
37
Controllo allosterico.
A
B
C
P
Modificazione covalente o interconversione
enzimatica
39
Cicli del substrato
A
Due reazioni inverse
accoppiate:
la velocità può essere
aumentata
è più sensibile a effettori
allosterici
B
r
f
C
D
Controllo genetico
La concentrazione degli enzimi, e quindi l'attività
enzimatica, può essere modificata dalla sintesi
della proteina in risposta alle necessità metaboliche
Si tratta di un meccanismo di controllo a lungo termine.
Composti ad alta energia
glucosio
C6H12O6 + 6 O2 → CO2 + 6 H2O
produzione di una notevole quantità di energia
(∆G = - 2850 kj x mole-1).
palmitato
C16H3202 + 23 O2
16 CO2 + 16 H2O
(DG°’= - 9781 kJ. mole-1).
L’energia rilasciata può essere recuperata
ATP e trasferimento del gruppo fosforico
43
L’importanza biologica dell’ATP risiede nella grande
quantità di energia che si libera durante la rottura dei
suoi legami fosfoanidridici.
ATP + H2O
ADP + Pi
ATP + H2O
AMP + PPi
44
La conoscenza dell’energia libera di
idrolisi di alcuni composti fosforici ci fa
capire la tendenza che essi hanno a
trasferire i loro gruppi fosforici agli
altri accettori.
Tali valori negativi sono chiamati
potenziali di trasferimento del gruppo
fosforico
45
Possono trasferire
un gruppo P all’ADP
Potenziale di
trasferimento
intermedio
46
Reazioni accoppiate -
Le reazioni esoergoniche dei composti ad alta
energia possono essere accoppiate a processi endoergonici per portarli a
completamento
(a)
Semi-reazione endoergonica
Semi-reazione esoergonica
Reazione complessiva accoppiata
(b)
Semi-reazione esoergonica
Semi-reazione endoergonica
Reazione complessiva accoppiata
ROTTURA PIROFOSFORICA NELLA SINTESI DI UN
AMMINOACIL-t-RNA
L’idrolisi altamente esoergonica del pirofosfato (-33,5 kJ mole-1)
spinge la reazione totale verso il suo compimento.
Posizione dell’ATP rispetto a composti
fosforici ad “alta energia” e a “bassa
energia”
L’ATP è continuamente
idrolizzato e rigenerato.
Nella gerarchia termodinamica
l’ATP occupa una posizione
intermedia.
Fosforilazione a livello di
substrato: ADP + PEP
Fosforilazione ossidativa
ATP + glucosio (chinasi)
Composti ad alto potenziale di trasferimento
del gruppo fosforico
50
Fonti di ATP durante l’esercizio fisico
La fosfocreatina costituisce una riserva ad alta
energia per la formazione di ATP
ATP + creatina
Fosfocreatina + ADP
In condizioni di riposo
Intensa attività metabolica
Fosfocreatina = “tampone” dell’ATP in cellule che contengono
creatina chinasi
Il ciclo
ATP-ADP
I combustibili più importanti
I nucleosidi trifosfati si possono convertire
l’uno nell’altro
ATP + NDP
ADP + NTP
Nucleoside difosfato chinasi.
Per la sintesi delle proteine, sintesi degli acidi nucleici
AMP + ATP
2 ADP
Cambiamenti conformazionali dell’adenilato
chinasi in conseguenza del legame con il
substrato
Struttura
chimica
dell’acetil CoA
L’acetil CoA è un composto ad alta energia.
L’idrolisi del suo legame tioestere è -31,5 kJ
mole -1
(reazione leggermente più esoergonica
dell’idrolisii dell’ATP )
La formazione di un legame tioestere in un
intermedio metabolico conserva una parte di
energia
Reazioni di ossidoriduzione
1. Quando avviene l'ossidazione a CO2 dei carburanti
metabolici, gli elettroni vengono trasferiti su
trasportatori molecolari che, negli organismi aerobici, li
trasferiscono all'ossigeno molecolare.
2. Il processo di trasporto degli elettroni
formazione di un gradiente di concentrazione
transmembrana che promuove la sintesi
(fosforilazione ossidativa).
causa la
protonica
di ATP
3. Le reazioni di ossido-riduzione (chiamate anche reazioni
redox) forniscono agli esseri viventi la maggior parte
della loro energia libera che viene conservata durante le
reazioni d’ossidazione dei substrati.
NAD+ e FAD
Riduzione reversibile
in modo da poter accettare gli elettroni,
passarli su altri trasportatori di elettroni e
quindi essere rigenerati per partecipare ad
altri cicli di ossidazione e di riduzione.
Riduzione del NAD+ a NADH
Riduzione del
FAD
a FADH2
Vitamina B2:
La vitamina B2, è un complesso di varie vitamine:
Riboflavina
Folato
Nicotinato
Pantotenato
Riboflavina: è un componente dei gruppi prostetici
flavina mononucleotide (FMN) e flavina adenina
dinucleotide (FAD).
O
N
CH3
N
CH3
N
O
N
C H2
H
OH
H
OH
H
OH
C H2O H
Ossidazioni e riduzioni
Ossidazione = perdita di elettroni
Riduzione = acquisto di elettroni.
I due processi sono sempre accoppiati
Donatore di elettroni e- + accettore di elettroni
Accettore di elettroni + e- donatore di elettroni
A forma ossidata + B forma ridotta A forma ridotta + B
forma ossidata
Durante le reazioni redox, gli elettroni trasferiti possono
passare direttamente dai donatori agli accettori:
Fe2+ + Cu2+ Fe3+ + Cu+
In alternativa, gli elettroni possono essere trasferiti da un
donatore a un accettore mediante un trasportatore di
elettroni:
X +Y XH2 + Y H2
Un atomo di idrogeno neutro è in grado di
trasferire un singolo elettrone:
H e- + H+
Lo ione negativo idruro (H-) è invece in grado di
trasferire due elettroni:
H- 2e- + H+
L’ossidazione biologica completa di sostanze organiche
nutrienti crea un flusso di elettroni che procede verso
l’ossigeno mediante una serie di trasportatori di
elettroni intermedi che si comportano come un ponte a
cestelli:
reazione redox esoergonica: gli elettroni passano
spontaneamente da un trasportatore a bassa affinità per
gli elettroni ad un altro con affinità maggiore e così via. La
reazione complessiva è:
sostanza organica + O2 CO2 + H2O + energia
Potenziali di riduzione
donatore(1) e- + accettore(1)
e- + accettore (2) donatore (2)
Reazione complessiva = donatore (1) + accettore (2)
accettore (1) + donatore (2)
Il potenziale di riduzione standard (E0) è una
misura quantitativa dell'affinità di un accettore
per gli elettroni in una semi-reazione.
Per convenzione internazionale, alla semi-reazione
H+ + e- 1/2H2
viene assegnato un potenziale di riduzione
standard pari a zero, quando la concentrazione di
ioni H+ è 1 M e H2 è presente alla pressione
parziale di 101,3 kPa.
Questa
semi-reazione
viene
usata
come
riferimento per la determinazione dei potenziali
delle altre semi-reazioni.
Il più forte agente
ossidante cioè ha
elevata affinità per
gli elettroni
Le reazioni di trasferimento degli elettroni
hanno una grande importanza biologica.
Nella catena di trasporto degli elettroni, gli
elettroni vengono trasferiti dal NADH lungo
una serie di accettori di elettroni con
potenziali di riduzione crescenti fino all’O2.
Questo porta alla sintesi di ATP