Membrana batterica cellulare o citoplasmatica La membrana cellulare ha una struttura a mo­
saico fluido come quella degli eucarioti, tutta­
via è priva di steroli. Fanno eccezione i mico
­ plasmi, che incorporano gli steroli nella mem­
brana quando si sviluppano in terreni che li contengono. Le principali funzioni della mem­
brana sono: barriera semipermeabile, piatta­
forma di supporto per enzimi della catena re­
spiratoria e delle biosintesi di fosfolipidi di membrana, di polimeri della parete e del DNA.
Le membrane cellulari batteriche formano in­
troflessioni o mesosomi, di cui se ne distin­
guono due tipi: mesosomi settali, che interven­
gono nella formazione del setto durante la di­
visione cellulare; mesosomi laterali, che costi­
tuiscono una piattaforma sulla quale si asso­
ciano proteine cellulari, quali gli enzimi della catena respiratoria (svolgendo una funzione analoga a quella dei mitocondri).
Nei batteri fotosintetici, i pigmenti fotosintetici sono posti in lamelle, che sono formate dalle introflessioni della membrana cellulare e si trovano subito sotto di essa, talora avvolgendo­
si a formare particelle distinte, dette cromato
­ fori.
La membrana plasmatica di tutti i batteri con­
tiene proteine di trasporto che utilizzano il gradiente di ioni H+ o Na+ per trasportare una varietà di nutrienti nella cellula. Altre proteine di trasporto utilizzano l'energia liberata dall'i­
drolisi di adenosintrifosfato (ATP) per traspor­
tare zuccheri, aminoacidi, vitamine e piccoli peptidi. Le proteine di trasporto sono dette transporters o permeasi e sono responsabili della diffusione facilitata [tipo canale o tipo carrier (uniporto)], del trasporto attivo prima­
rio, del trasporto attivo secondario (tipo sim­
porto o antiporto) e del trasporto con fosforila­
zione del substrato (fosfotransferasi). Circa la metà delle proteine di trasporto dei batteri appartengono al sistema di trasporto attivo primario ABC (ATPase Binding Cassette) e al sistema di diffusione facilitata/trasporto attivo secondario MFS (major facilitator superfamily). Le permeasi batteriche sono generalmente inducibili, per cui la densità delle proteine di trasporto nella membrana è regolata dalla concentrazione del soluto nel mezzo e dalle necessità metaboliche della cellula.
Il trasporto dal citoplasma allo spazio extraci­
toplasmatico comprende due sistemi di efflus­
so noti, entrambi presenti nella membrana ci­
toplasmatica: sistema antiporto H+/farmaci e proteine della famiglia ABC.
Le ABC permeasi trasportano sia piccole mo­
lecole che macromolecole in risposta alla idro­
lisi di ATP. Questo sistema di trasporto è com­
posto da due proteine integrali di membrana con sei segmenti transmembranosi, due protei­
ne periferiche associate sul versante citopla­
smatico, che legano idrolizzano l'ATP, e un proteina o lipoproteina recettoriale periplasmi­
ca che lega il substrato. Le ABC permeasi più studiate comprendono il sistema di trasporto del maltosio di Escherichia coli e quello dell'i­
stidina di Salmonella typhimurium. Dal mo­
mento che i batteri gram­positivi sono privi della membrana esterna, il recettore, una volta secreto, si perderebbe nell'ambiente extracellu­
lare. Di conseguenza, questi recettori risultano legati alla superficie esterna della membrana citoplasmatica mediante ancore lipidiche. Poi­
ché di frequente i batteri vivono in mezzi dove la concentrazione di nutrienti è bassa, le pro­
teine ABC permettono alla cellula di concen­
trare i nutrienti nel citoplasma contro il gra­
diente di concentrazione.
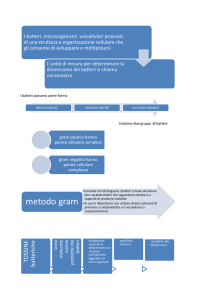
Parete cellulare La parete cellulare presenta una struttura note­
volmente diversa a seconda che si tratti di batte­
ri gram­positivi o gram­negativi, anche se il peptidoglicano costituisce la sostanza universal­
mente presente nella parete cellulare dei batteri. Nei batteri gram­negativi lo strato di peptidogli­
cano è piuttosto sottile, con uno spessore di cir­
ca 50­100 Å. La maggioranza dei batteri gram­
positivi ha invece una parete cellulare relativamente spessa (circa 200­800 Å), in cui al peptidoglicano sono covalentemente legati altri polimeri, quali acidi teicoici, polisaccaridi e peptidoglicolipidi. Esternamente al peptidoglicano i batteri gram­negativi hanno una membrana esterna di spessore di circa 75­100 Å.
mente più complessa, in quanto esternamente allo strato di peptidoglicano è presente la membrana esterna; le due strutture sono legate dalla lipoproteina.
Il peptidoglicano è un polimero composto da: una catena principale, identica in tutte le specie batteriche, formata da subunità disaccaridiche di N­acetilglucosamina e da acido N­acetilmu
­ ramico, unite da legami Beta, 1­4; catene latera­
li di un identico tetrapeptide, legato all'acido N­
acetilmuramico; di solito, una serie di ponti peptidici trasversali, che uniscono i tetrapeptidi di polimeri adiacenti. I tetrapeptidi dei polimeri adiacenti possono essere legati, invece che da ponti peptidici, da legami diretti tra la D­alanina di un tetrapeptide e la L­lisina o l'acido diami­
nopimelico del tetrapeptide adiacente. Le catene tetrapeptidiche laterali e i ponti trasversali va­
riano a seconda della specie batterica.
saccaride composto da una parte centrale e da una serie terminale di unità ripetute. Il lipide A è formato da una catena di disaccaridi della glu­
cosamina, uniti da ponti di pirofosfato, a cui sono legati numerosi acidi grassi a catena lunga, fra cui l'acido beta­idrossimiristico (C14), sem­
pre presente è caratteristico di questo lipide.
La componente proteica della lipoproteina è unita con legame peptidico ai residui di DAPA (acido diaminopimelico) delle catene laterali te­
trapeptidiche del peptoglicano, mentre la com­
Il peptidoglicano, detto anche mucopeptide o ponente lipidica è fissata con legame covalente mureina, è composto da un peptide complesso alla membrana esterna, del cui foglietto interno formato da un polimero di aminoglucidi e pepti
­ è una componente importante.
di. Nei batteri gram­positivi, è disposto in mol­ La membrana esterna ha la struttura tipica delle teplici strati, tanto da rappresentare dal 50% al membrane biologiche. Gran parte del foglietto 90% del materiale della parete cellulare, mentre fosfolipidico esterno è composto da molecole di nei gram­negativi vi sono uno o al massimo due lipopolisaccaride (LPS), o endotossina dei bat­
strati di peptidoglicano, che costituiscono il teri gram­negativi, formato da un lipide com­
5%­20% della parete.
plesso, chiamato lipide A, a cui è unito un poli
­ La parte centrale del polisaccaride è costante in tutte le specie batteriche gram­negative, mentre le unità ripetute sono specie­specifiche e sono costituite di solito da trisaccaridi lineari oppure da tetrasaccaridi o pentasaccaridi ramificati. Il polisaccaride costituisce l'antigene O di superfi­
cie e la specificità antigenica è dovuta alle unità ripetute terminali. La tossicità del LPS è invece Il peptidoglicano dei batteri gram­positivi è le­ dovuta al lipide A.
gato a molecole accessorie, come acidi teicoici, Fra le principali proteine della membrana ester­
acidi teucuronici, polifosfati o carboidrati. La na, le più abbondanti sono le porine. Le porine maggior parte dei batteri gram­positivi contiene sono proteine transmembranose, organizzate in considerevoli quantità di acidi teicoici, fino al triplette, ciascuna subunità è formata da 16 do­
50% del peso umido della parete. Si tratta di mini in conformazione beta a disposizione an­
polimeri idrosolubili, formati da ribitolo o gli
­ tiparallela che danno origine ad una struttura ci­
cerolo, uniti da legami fosfodiesterici. Il ribitolo lindrica cava. Il canale consente la diffusione di e il glicerolo possono legare residui glucidici, molecole idrofiliche di p.m. < 600­700 Da (fo­
come glucosio, galattosio o N­acetilglucosami­ sfati, disaccaridi,ecc.), mentre le molecole idro­
na, e di solito D­alanina, in genere legata in po­ fobiche (compresi alcuni antibiotici beta­latta­
sizione 2 o 3 del glicerolo oppure 3 o 4 del ribi­ mici, come ampicillina e cefalosporine) posso­
tolo. Gli acidi teicoici rappresentano i principali no attraversare la componente lipidica della antigeni di superficie dei batteri gram­positivi membrana esterna.
che li contengono.
Altre proteine della membrana esterna permet­
La parete dei batteri gram­negativi è notevol­ tono la diffusione facilitata di numerose sostan­
ze, quali maltosio, vitamina B12, nucleosidi e Tra la membrane interna e quella esterna è com­
complessi ferro­carboniosi, mentre non sembra preso lo spazio periplasmico, parzialmente oc­
siano presenti sistemi di trasporto attivo.
cupato dal peptoglicano con la sua porosità. In Oltre alle proteine di trasporto,sono presenti re­ questo spazio sono presenti le proteine peripla­
cettori per la coniugazione batterica, per i fagi e smiche: binding­proteins, che specificamente le colicine (il recettore per il fago T6 e la colici
­ legano zuccheri, aminoacidi e ioni, coinvolte na k è anche implicato nel trasporto dei nucleo
­ nell'attività recettoriale e di trasporto; enzimi, come le betalattamasi, codificate dai plasmidi.
sidi).
Rispondi sul quaderno
1) Cos'è un antigene?
2) Cos'è un recettore?
3) Cos'è un sistema di diffusione facilitato?


![5\) batteri [modalità compatibilità]](http://s1.studylibit.com/store/data/000928299_1-28a224c15810efb654d8d9e4b207aa31-300x300.png)