Parte II- Bioenergetica e metabolismo di carboidrati e lipidi
Risposte e breve discussione
1)
2)
3)
4)
5)
6)
7)
8)
9)
d. Il glucosio-6-fosfato è l'unico estere tra le sostanze elencate. L'energia libera di idrolisi
standard di questo composto è di circa 4 kcal/mole. Tutti gli altri composti hanno energie
libere di idrolisi superiori alle 7 kcal/mole.
d. Le ossidasi sono ossidoreduttasi che utilizzano ossigeno come accettore di elettroni
producendo acqua ossigenata. Le deidrogenasi, flaviniche e piridiniche, utilizzano i rispettivi
coenzimi FAD e NAD (NADP) come agenti ossidanti. Le perossidasi impiegano acqua
ossigenata come agente ossidante. Le decarbossilasi non catalizzano reazioni di
ossidoriduzione.
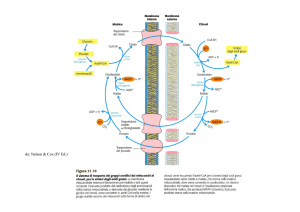
b. L'ipotesi chemioosmotica di Mitchell spiega l'accoppiamento tra la fosforilazione
ossidativa e il trasporto elettronico. Il trasferimento degli elettroni lungo la catena di
trasporto elettronico comporta un trasferimento di protoni dalla matrice mitocondriale allo
spazio intermembrana. I protoni vengono pompati a livello di tre siti della catena respiratoria
generando così un gradiente protonico. Quando i protoni tornano nella matrice attraversando
i canali protonici dell'ATPasi asimmetrica posta nella membrana mitocondriale interna, si
sintetizza ATP. Oggigiorno è comunemente accettato che l'ipotesi di Mitchell spieghi in
modo corretto il legame tra fosforilazione ossidativa e trasporto elettronico.
d. L'oligomicina inibisce l'ATPasi mitocondriale e quindi previene la fosforilazione
dell'ADP a formare ATP. Impedisce l'utilizzazione dell'energia derivata dal trasporto
elettronico per la sintesi di ATP. L'oligomicina non ha alcun effetto sull'accoppiamento ma
blocca la fosforilazione mitocondriale bloccando sia l'ossidazione sia la fosforilazione.
d. I sistemi navetta sono utilizzati per il trasferimento degli elettroni presenti sul NADH
prodotto nel citosol alla catena respiratoria mitocondriale. Si tratta di due sistemi, noti come
sistema del glicerofosfato e del malato-aspartato, che operano inizialmente una riduzione,
rispettivamente, del diossiacetone fosfato e dell'ossalacetato citosolico.
d. Il fruttosio è l'unico chetoso tra i monosaccaridi elencati. Il glucosio è l'aldoesoso più
importante da un punto di vista metabolico. Mannosio e galattosio sono epimeri del glucosio
rispettivamente in posizione 2 e 4. Il ribosio è un aldoso a cinque atomi di carbonio
particolarmente importante per la sua presenza nell'acido ribonucleico.
d. Il glicogeno e l'amido sono due polimeri costituiti esclusivamente da α-glucosio. In
entrambi questi polisaccaridi le singole unità monosaccaridiche sono unite tra loro con
legami 1,4- o 1,6-glicosidici. Il glicogeno, diffuso nel mondo animale, presenta un numero
di ramificazioni sensibilmente superiore rispetto all'amido. In nessuna delle due molecole
sono presenti catene polipeptidiche coniugate.
c. Il concetto di essenzialità si riferisce all'impossibilità di biosintetizzare una molecola in un
determinato organismo. In questi casi le molecole essenziali debbono essere introdotte con
l'alimentazione. Nel caso degli acidi grassi, l'uomo non è in grado di sintetizzare doppi
legami ad una distanza uguale o inferiore a sei atomi di carbonio dall'estremità metilica
terminale. L'acido linoleico, che presenta un doppio legame a distanza di sei atomi di
carbonio dall'estremità metilica (ω6), è pertanto essenziale per l'uomo. Gli acidi palmitoleico
e oleico sono rispettivamente ω7 e ω9 e quindi non sono essenziali. Gli acidi palmitico e
stearico non contengono doppi legami.
b. Le lecitine sono particolari tipi di fosfogliceridi costituiti da uno scheletro di glicerolo a
cui sono esterificate due molecole di acido grasso. La terza posizione alcolica del glicerolo è
esterificata da una molecola di acido fosforico che, a sua volta, è legata ad una molecola di
colina. Le molecole di zucchero, quali il ribosio, non sono presenti nelle lecitine. La
sfingosina è invece una molecola che costituisce lo scheletro di un'altra classe di fosfolipidi,
le sfingomieline.
10)
b. Una molecola di GTP si sintetizza a partire da GDP e fosfato inorganico nella reazione in
cui il succinil CoA è idrolizzato a succinato e CoA. Questa reazione rappresenta una
fosforilazione a livello del substrato e, in contrapposizione alla fosforilazione ossidativa, è
l'unica reazione del ciclo dell'acido citrico che fornisce direttamente una molecola fosforilata
ad alta energia libera di idrolisi. La sequenza di reazioni che parte dall'α-chetoglutarato per
arrivare al succinato è catalizzata dal complesso dell' α-chetoglutarato deidrogenasi e della
succinil CoA sintetasi.
α-chetoglutarato + NAD+ + CoA → succinil CoA + CO2 + NADH
succinil CoA + Pi + GDP → succinato + GTP + CoA
11)
12)
13)
14)
15)
16)
b. Equivalenti riducenti sono prodotti in quattro momenti del ciclo dell'acido citrico. Il
NADH è prodotto dalla conversione dell'isocitrato in α-chetoglutarato catalizzata
dall'isocitrato deidrogenasi, dalla conversione dell'α-chetoglutarato in succinil CoA
catalizzata dal complesso dell'α-chetoglutarato deidrogenasi e dalla conversione del malato
in ossalacetato catalizzata dalla malato deidrogenasi. Il FADH2 è prodotto dalla conversione
del succinato in fumarato catalizzata dalla succinato deidrogenasi. La succinil CoA sintetasi
catalizza la formazione di succinato a partire da succinil CoA, con la formazione
contemporanea di GTP a partire da GDP e fosfato inorganico.
b. La glicolisi aerobia può essere definita come la conversione ossidativa del glucosio a due
molecole di piruvato. In questo processo si generano due molecole di ATP e due molecole di
NADH. Poiché gli equivalenti riducenti citosolici debbono essere trasferiti nel mitocondrio
per la riossidazione, non è certo il numero di molecole di ATP prodotte. Assumendo che si
formino due molecole di ATP per ogni NADH citosolico trasferito nel mitocondrio con il
sistema navetta del glicerofosfato e qui riossidato, la resa energetica della glicolisi aerobia
può essere stimata in sei molecole di ATP per molecola di glucosio utilizzata.
e. Gruppi fosfato sono trasferiti su molecole di substrato in tre reazioni della glicolisi.
Tuttavia, solo il trasferimento di fosfato sull'aldeide 3-fosfoglicerica accoppiato alla sua
ossidazione e catalizzato dalla gliceraldeide 3-fosfato deidrogenasi genera un composto ad
alta energia libera di idrolisi, l'1,3-bifosfoglicerato. Le fosforilazioni del glucosio e del
fruttosio-6-fosfato, catalizzate rispettivamente da esochinasi e fosfofruttochinasi, portano
alla formazione di esteri fosforici il cui contenuto energetico non è considerato elevato. La
fosfoglicerato chinasi e la piruvato chinasi sono enzimi che catalizzano il trasferimento di
gruppi fosfato da composti ad alta energia libera di idrolisi all'ADP per formare ATP.
b. Il prodotto finale della glicolisi negli eritrociti, che non possiedono mitocondri, è il lattato.
La conversione del piruvato in lattato catalizzata dalla lattato deiderogenasi permette di
rigenerare il NAD+ dal NADH accumulato nella reazione di ossidazione della gliceraldeide3-fosfato. Ciò fa si che la glicolisi possa procedere. Il lattato si accumula anche nel muscolo
in attività intensa, quando l'ossigeno non è più sufficiente. Nei mammiferi, il lattato diffonde
in circolo ed è metabolizzato nel fegato. Il lattato si forma in modo simile nei microrganismi
quando questi crescono in condizioni anaerobie. In alternativa, i microrganismi (ma non gli
eritrociti) possono formare etanolo in condizioni di anaerobiosi. In tutti i casi il prodotto
finale favorisce la rigenerazione del NAD+ e il procedere della glicolisi.
c. La glicogeno sintetasi è un enzima che trasferisce unità glicosidiche dall'UDP-glucosio
alla molecola di glicogeno in crescita. Nelle piante è l'ADP-glucosio a fare la parte
dell'UDP-glucosio degli animali. L'enzima esiste in due forme: l'una attiva, defosforilata,
l'altra inattiva, fosforilata. L'inattivazione dipende perciò dalla fosforilazione di un residuo
specifico di serina. La depolimerizzazione del glicogeno, non la sintesi, è influenzata
positivamente dall'aumento dei livelli di ione calcio.
b. Le condizioni patologiche note come galattosemie possono derivare da un deficit di
galattochinasi, 4'-epimerasi o uridiltransferasi. Ognuno di questi deficit conduce ad un
alterato metabolismo del galattosio, che si forma per idrolisi del lattosio, lo zucchero del
17)
18)
19)
20)
21)
latte. L'accumulo prolungato di galattosio nel sangue, che viene in parte perduto con le
urine, porta alla cataratta. Il galattosio viene, infatti, ridotto a galattitolo e accumulato
nell'occhio. Tutti gli altri enzimi elencati nelle risposte partecipano al metabolismo del
glicogeno e il loro deficit porta all'accumulo di glicogeno.
d. Nel momento in cui una cellula facoltativa, che può metabolizzare glucosio sia in
condizioni aerobie sia anaerobie, passa dal metabolismo anaerobio a quello aerobio, riduce
nettamente il consumo di glucosio. In condizioni aerobie la cellula guadagna, in termini di
resa energetica del catabolismo del glucosio, fino a 18 volte rispetto al metabolismo
anaerobio. E' evidente che in tali condizioni la cellula richieda una minor quantità di
glucosio. L'aumento dei livelli di ATP citosolico e il rilascio del citrato dal mitocondrio che
avvengono in condizioni aerobie, inibiscono allostericamente la fosfofruttochinasi, enzima
chiave della via glicolitica.
d. Tutti gli enzimi elencati, ad eccezione della fosfoglicerato chinasi, sono specifici della
glicolisi o della gluconeogenesi. La fosfoglicerato chinasi è uno dei sette enzimi comuni alle
due vie metaboliche. Gli enzimi esochinasi, fosfofruttochinasi e piruvato chinasi catalizzano
irreversibilmente le rispettive reazioni della glicolisi. Nella gluconeogenesi tali passaggi
sono catalizzati da enzimi diversi. Il piruvato è trasformato in fosfoenolpiruvato attraverso
due passaggi. Nel primo si forma ossalacetato per carbossilazione catalizzata dalla piruvato
carbossilasi; nel secondo la fosfoenolpiruvato carbossichinasi decarbossila e fosforila
l'ossalacetato in presenza di GTP. L'altro passaggio irreversibile che deve essere saltato nella
gluconeogenesi prevede la formazione di fruttosio-6-fosfato da fruttosio-1,6-bifosfato per
azione della fruttosio-1,6-fosfatasi. Quando il glucosio-6-fosfato è finalmente prodotto nella
gluconeogenesi, questo è convertito in glucosio dalla glucosio-6-fosfatasi, un enzima
presente soltanto nel reticolo endoplasmatico degli epatociti. Il glucosio libero può così
diffondere nel sangue dal fegato. Tra tutti gli enzimi indicati nelle risposte, soltanto la
fosfoglicerato chinasi catalizza una reazione reversibile comune alla glicolisi e alla
gluconeogenesi.
d. L'adrenalina stimola l'adenilato ciclasi muscolare e epatica portando alla produzione di
AMP ciclico. Nel fegato, l'aumento dei livelli di cAMP comporta l'attivazione di una
fosfatasi che defosforila il fruttosio-2,6-bifosfato e, contemporaneamente, inattiva la chinasi
che lo produce. Il risultato di questo processo è che i livelli di fruttosio-2,6-bifosfato si
riducono e che l'attività della fosfofruttochinasi diminuisce. Nel muscolo scheletrico,
tuttavia, la chinasi responsabile della sintesi di fruttosio-2,6-bifosfato è attivata, piuttosto che
inibita, dal cAMP. Ne consegue che la stimolazione da adrenalina provoca nel muscolo un
aumento del flusso glicolitico, contrariamente a ciò che succede nel fegato. In entrambi i
tessuti la glicogeno fosforilasi è attivata e si ha intensa glicogenolisi. In queste condizioni, il
glucosio è impiegato in modo diverso nei due organi: nel muscolo serve per la produzione di
ATP utile per l'attività contrattile mentre nel fegato esso viene esportato nel sangue.
a. Le fonti metaboliche di NADPH per la sintesi degli acidi grassi e del colesterolo sono la
via del pentoso fosfato ed il malato che si genera durante il trasferimento di gruppi acetile
dal mitocondrio al citosol sottoforma di citrato. L'enzima citrato liasi spezza il citrato in
acetil CoA e ossalacetato. Quest'ultimo è ridotto a malato dal NADH. L'enzima malico
NADP-dipendente catalizza la decarbossilazione ossidativa del malato a piruvato e biossido
di carbonio. Quindi la diffusione del citrato in eccesso dal mitocondrio al citosol delle
cellule non fornisce soltanto acetil CoA per la sintesi di acidi grassi e colesterolo ma anche il
NADPH. Per ogni molecola di acetil CoA prodotta si forma una molecola di NADPH.
Tuttavia, la maggior parte del NADPH richiesto per la sintesi degli acid grassi è fornito dalla
via del pentoso fosfato. Per questo motivo nel tessuto adiposo questa via metabolica è
particolarmente attiva.
b. In condizioni normali, il galattosio è convertito rapidamente in glucosio-6-fosfato nel
fegato e negli eritrociti. La maggior parte degli altri organi non metabolizza il galattosio. Il
galattosio è inizialmente fosforilato dall'ATP per produrre galattosio-1-fosfato in presenza
22)
23)
24)
25)
26)
27)
dell'enzima galattochinasi. La galattosio-1-fosfato uridiltransferasi trasferisce quindi l'UDP
dall'UDP-glucosio al galattosio-1-fosfato per formare UDP-galattosio e glucosio-1-fosfato.
Grazie alla presenza di UDP-galattosio-4-epimerasi, l'UDP-galattosio è epimerizzato a UDPglucosio. Il glucosio-1-fosfato è, infine, isomerizzato a glucosio-6-fosfato dalla
fosfoglucomutasi. Il deficit genetico di galattosio-1-fosfato uridiltransferasi provoca la
galattosemia. In questa malattia autosomica recessiva, i bambini affetti hanno complicazioni
fino a che il latte, principale fonte di galattosio, non è eliminato dalla dieta. I principali
sintomi che permangono fino a quando non si elimina il galattosio dall'alimentazione sono
vomito, diarrea, epatomegalia, ittero e ritardo mentale. I problemi di sviluppo sono causati
dalla riduzione del galattosio a galattitolo (dulcitolo) in presenza dell'enzima aldoso
reduttasi. L'accumulo di galattosio-1-fosfato è inoltre responsabile dell'epatomegalia.
e. Il complesso dell'acido grasso sintetasi di mammifero è costituito da due subunità
identiche. Ognuna delle due subunità è un complesso multienzimatico costituito da sette
attività enzimatiche e dalla proteina trasportatrice di gruppi acile (ACP). Tutte le componenti
sono legate covalentemente tra loro e sono quindi comprese in una singola catena
polipeptidica che funziona soltanto in presenza di un'altra catena polipeptidica identica. Ogni
ciclo di sintesi vede il funzionamento della proteina trasportatrice di acili e di sei attività
enzimatiche: acetil transferasi, malonil transferasi, β-chetoacil sintasi, β-chetoacil reduttasi,
deidratasi ed enoilreduttasi. Quando la catena idrocarburica raggiunge la lunghezza finale, in
genere C16 , la tioesterasi idrolizza l'acido grasso staccandolo dal complesso della sintasi.
d. Nell'uomo il prodotto finale della sintesi degli acidi grassi che avviene nel citosol è l'acido
palmitico. La specificità del complesso multienzimatico dell'acido grasso sintetasi è tale che
non appena si raggiunge la lunghezza di 16 atomi di carbonio, una tioesterasi stacca l'acido
grasso dal complesso. L'allungamento e l'insaturazione del palmitato appena sintetizzato e
degli acidi grassi introdotti con la dieta avviene per azione di enzimi localizzati nelle
membrane del reticolo endoplasmatico liscio.
a. Durante ogni ciclo di β-ossidazione l'acil CoA saturo viene degradato attraverso un
accorciamento di due atomi di carbonio rilasciati in forma di acetil CoA. Complessivamente
avvengono quattro reazioni: l'ossidazione dell'acil CoA ad enoil CoA con produzione di
FADH2 ; l'idratazione, con consumo di H2 O, per formare L-idrossiacil CoA; l'ossidazione a
chetoacil CoA associata alla formazione di NADH + H+ ; infine, la rottura del chetoacil CoA
per introduzione di una nuova molecola di CoA per fornire acetil CoA e un acil CoA
accorciato di due atomi di carbonio.
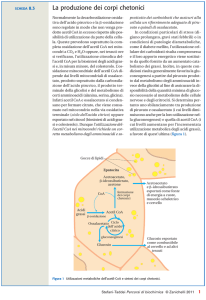
e. Il precursore metabolico dell'acetacetil CoA prodotto dalla condensazione di acetil CoA
nel fegato è il 3-idrossi-3-metilglutaril CoA (HMG CoA). In condizioni normali, la
produzione di HMG CoA avviene nel citosol degli epatociti, nel quadro della biosintesi del
colesterolo. A digiuno, tuttavia, ma anche in pazienti diabetici, la produzione di HMG CoA
avviene nei mitocondri epatici durante la biosintesi dei corpi chetonici. In questo processo,
l'HMG CoA viene spezzato dalla HMG CoA liasi per dare acetacetato e acetil CoA.
L'enzima β-idrossibutirrato deidrogenasi, NADH-dipendente, converte la maggior parte
dell'acetacetato in β-idrossibutirrato che diffonde nel sangue ed è trasportato ai tessuti
periferici.
c. Nei mammiferi l'acido arachidonico (5,8,11,14-eicosatetraenoico) può essere sintetizzato
esclusivamente a partire da acidi grassi essenziali introdotti con la dieta. Dall'acido linoleico
(9,12-ottadecadienoico) si può ottenere acido arachidonico mediante due insaturazioni e
allungamento della catena. Nonostante l'acido linolenico (9,12,15-ottadecatrienoico) sia
anch'esso un acido grasso essenziale, la sua insaturazione e l'allungamento portano alla
formazione di acido 8,11,14,17-eicosatetraenoico, una molecola diversa dall'acido
arachidonico. Gli acidi palmitico, oleico e stearico non sono acidi grassi essenziali nei
mammiferi e non possono portare alla formazione di acido arachidonico.
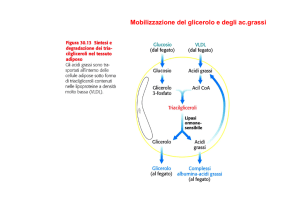
b. La lipolisi negli adipociti è regolata direttamente dagli ormoni. L'adrenalina stimola
l'adenilato ciclasi a produrre AMP ciclico che, a sua volta, attiva una proteina chinasi. La
28)
29)
30)
chinasi attiva la trigliceride lipasi attraverso una reazione di fosforilazione. La lipolisi quindi
prosegue e porta al rilascio di acidi grassi e glicerolo. L'inutile riesterificazione dell'acido
grasso è prevenuta dal fatto che gli adipociti possiedono una bassissima attività
glicerochinasica per fosforilare il glicerolo libero, che deve quindi essere metabolizzato nel
fegato. L'inibizione della lipolisi avviene in presenza di insulina, che determina un
abbassamento dei livelli di AMP ciclico. La lipoproteina lipasi non è un enzima presente
negli adipociti.
d. la regolazione del metabolismo del colesterolo è per definizione esercitata su di una tappa
obbligata. Questa è la reazione catalizzata dalla 3-idrossi-3-metilglutaril CoA reduttasi.
L'attività reduttasica diminuisce in modo sensibile quando la dieta è ricca di colesterolo e ciò
rappresenta un efficace sistema di controllo retrogrado del metabolismo del colesterolo.
b. I chilomicroni sono lipoproteine ricche in trigliceridi di origine alimentare. Le
lipoproteine a densità molto bassa (VLDL) contengono trigliceridi e colesterolo di origine
epatica. La progressiva riduzione della quota di trigliceridi contenuta nelle VLDL porta alla
formazione delle lipoproteine a densità intermedia (IDL) e, infine, di quelle a densità bassa
(LDL). Da qui l'ordine chilomicroni → VLDL → IDL → LDL
e. Gli acidi biliari sono spesso coniugati con la glicina per formare l'acido glicocolico o con
la taurina per formare l'acido taurocolico. Nella bile umana l'acido glicocolico è senz'altro il
più abbondante. La presenza del gruppo carbossilato della glicina o del gruppo solfato della
taurina aumenta l'idrofilicità degli acidi biliari e comporta, di conseguenza, una maggior
efficacia da parte di questi nell'emulsionamento dei lipidi durante la digestione.