La cellula
Chimica e Macromolecole
Proteine
DNA replicazione
RNA trascrizione
Traduzione
Citoscheletro
Organelli Cellulari
Molti degli organelli
delimitati da
membrana
mantengono una
collocazione fissa
nella cellula
attaccandosi al
citoscheletro
(microtubuli) che fa
da binario per lo
spostamento degli
organelli e delle
vescicole oggetto di
scambio tra gli
organelli stessi
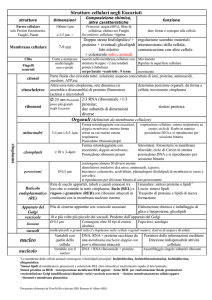
Funzione principale dei comparti delimitati da
membrana in una cellula eucariotica
Compartimento
Funzione primaria
Citosol
Contiene molte vie metaboliche; sintesi proteica
Nucleo
Contiene il genoma principale; sintesi di DNA e RNA
Reticolo endoplasmatico (RE)
Sintesi di quasi tutti i lipidi; sintesi di proteine da
distribuire a vari organelli e alla membrana plasmatica
Apparato di Golgi
Modificazione, smistamento e confezione di proteine e
lipidi o per la secrezione o per l’invio ad altri organelli
Lisosomi
Degradazione intracellulare
Endosomi
Smistamento di materiali assunti per endocitosi
Mitocondri
Sintesi di ATP per fosforilazione ossidativa
Cloroplasti
Sintesi di ATP e fissazione del carbonio per fotosintesi
Perossisomi
Ossidazione di molecole tossiche
Volumi relativi occupati dai principali organelli
delimitati da membrane (epatocita)
Compartimento intracellulare
Percentuale del volume
cellulare totale
Numero approssimativo
per cellula
Citosol
54
1
Mitocondri
22
1700
Reticolo endoplasmatico (RE)*
12
1
Nucleo
6
1
Apparato di Golgi
3
1
Perossisomi
1
400
Lisosomi
1
300
Endosomi
1
200
*20-30 volte l’area della membrana plasmatica
La compartimentalizzazione cellulare
• Le cellule svolgono migliaia di reazioni chimiche diverse
(spesso di segno opposto) in ogni istante quindi devono
tenerle separate tra di loro.
• La cellula si è evoluta sviluppando diverse strategie per
organizzare e compartimentare tali reazioni:
– Aggregare in un solo complesso proteico i vari enzimi per
cercare di catalizzare una certa sequenza di reazioni (v.d. DNA
e RNA).
– Confinare processi metabolici diversi in compartimenti diversi
(organelli delimitati da membrana).
La compartimentalizzazione cellulare
• E’ possibile per la presenza di organelli delimitati da membrana.
• E’ ottenuta mediante lo smistamento (sorting) delle proteine:
– Sistema utilizzato per mantenere la composizione proteica dei vari comparti (ogni comparto
possiede un corredo specifico di proteine arrivate dal citosol, sede della sintesi, per svolgere
la loro funzione)
– Questo processo di smistamento (sorting) si basa su appositi segnali inclusi nella sequenza
aminoacidica della proteina
• E’ ottenuta mediante il trasporto vescicolare
– Insieme dei contatti che alcuni dei comparti delimitati da membrana intrattengono tra di loro
tramite la formazione di sferule membranose che si staccano da un comparto attraversano il
citosol e vanno a fondersi con un altro comparto
• E’ ottenuta mediante l’utilizza delle vie secretorie
– Rilascio di proteine all’esterno della cellula (esocitosi)
– Assunzione di proteine dall’esterno della cellula (endocitosi)
La compartimentalizzazione cellulare
Sorting delle proteine
•
E’ possibile per la presenza di organelli delimitati da membrana.
•
E’ ottenuta mediante lo smistamento (sorting) delle proteine:
– Sistema utilizzato per mantenere la composizione proteica dei vari
comparti (ogni comparto possiede un corredo specifico di proteine
arrivate dal citosol, sede della sintesi, per svolgere la loro funzione)
– Questo processo di smistamento (sorting) si basa su appositi segnali
inclusi nella sequenza aminoacidica della proteina
TUTTI QUESTI
PROCESSI
NECESSITANO DI
ENERGIA
NEI MECCANISMI 1 E 3
LA PROTEINA
MANTIENE LA SUA
CONFORMAZIONE
Macromolecole trasporto attivo
Piccole molecole per diffusione
Traslocatori proteici situati
nella membrana
NEL MECCANISMO 2 LA
PROTEINA DEVE
DISTENDERSI
Viaggiano a bordo di vescicole di
trasporto per andare dal RE
all’Apparato di Golgi, endosomi,
lisosomi e superficie cellulare
Sequenze segnale (15-60 aa)
In genere vengono poi rimosse una volta a destinazione
Sono necessarie e sufficienti a dirigere una proteina
in un determinato luogo
Funzione del segnale
Esempio di sequenza segnale
Importazione nel RE
+H3N-Met-Met-Ser-Phe-Val-Ser-Leu-Leu-Leu-Val-Gly-Iie-Leu-
Phe-Trp-Ala-Thr-Glu-Ala-Glu-Gln-Leu-Thr-Lys-Cys-Glu-ValPhe-Gln
Ritenzione nel lume del RE
-Lys-Asp-Glu-Leu-COO-
Importazione nei mitocondri
+H3N-Met-Leu-Ser-Leu-Arg-Gln-Ser-Ile-Arg-Phe-Phe-Lys-Pro-
Ala-Thr-Arg-Thr-Leu-Cys-Ser-Ser-Arg-Tyr-Leu-Leu
Importazione nel nucleo
-Pro-Pro-Lys-Lys-Lys-Arg-Lys-Val-
Importazione nei perossisomi
-Ser-Lys-Leu
Gli aa carichi positivamente sono scritti in rosso e quelli carichi negativamente in azzurro, in arancione un gruppo
cospicuo di aa idrofobici. Spesso conta di più le proprietà fisiche degli aa (idrofobicità, posizione degli aa) che la
sequenza in senso stretto per lo smistamento
La sequenza segnale è necessaria e sufficiente per inviare una proteina nel RE
Viene rimossa dalla proteina finita una volta che è stata smistata
TUTTI QUESTI
PROCESSI
NECESSITANO DI
ENERGIA
NEI MECCANISMI 1 E 3
LA PROTEINA
MANTIENE LA SUA
CONFORMAZIONE
Macromolecole trasporto attivo
Piccole molecole per diffusione
Traslocatori proteici situati
nella membrana
NEL MECCANISMO 2 LA
PROTEINA DEVE
DISTENDERSI
Viaggiano a bordo di vescicole di
trasporto per andare dal RE
all’Apparato di Golgi, endosomi,
lisosomi e superficie cellulare
Nucleo cellulare
Dimensione 5 micron. Doppia membrana a distanza di 20-40 nm
Pori nucleari di 100 nm, in cui membrana interna ed esterna si fondono
Membrana interna è costituita da lamina nucleare-intreccio di filamenti
intermedi
Contiene DNA complessato con proteine=nucleosomi…spiralizzato in
cromosomi
Sintesi di RNA da DNA: passa attraverso i pori nucleari
per essere tradotto in proteine
Figure 1-15 Essential Cell Biology (© Garland Science 2010)
La membrana nucleare
interna contiene proteine che
agiscono da legame per i
cromosomi e per la lamina
nucleare (rete di filamenti che
conferisce rigidità al nucleo)
La composizione
dell’involucro nucleare
esterno non differisce da
quella del RE di cui ne è la
continuazione
I pori nucleari (100 proteine)
sono i portali da cui entrano
ed escono dal nucleo tutte le
molecole, il traffico dai pori
circola in entrambi i sensi
Il complesso del poro
Circa 30 diverse proteine
Filamenti intermedi della lamina nucleare
• Sono costituiti da LAMINE
• Nucleo si disgrega nella divisione cellulare insieme alla lamina
anche che viene fosforilata, mentre la defosforilazione al termine
della divisione le fa riassociare assemblare.
• Fosfo-defosforilazione TESTE GLOBULARI dei filamenti
intermedi
• Mutazioni genetiche associate a mancanza di lamine
- Progeria invecchiamento precoce da danno al nucleo,
apoptosi generalizzata
- Neuropatie
- Cardiomiopatie
Sam Burns
• In rosso segnale di localizzazione nucleare
• Le fibrille guidano nel poro il complesso recettore verso il vettore
• Il trasporto è attivo ed utilizza energia liberata dall’idrolisi del GTP
• I pori nucleari trasportano le proteine già montata nella loro conformazione
nativa (in altri organelli le proteine devono distendersi per entrare)
TUTTI QUESTI
PROCESSI
NECESSITANO DI
ENERGIA
Macromolecole trasporto attivo
Piccole molecole per diffusione
NEI MECCANISMI 1 E 3
LA PROTEINA
MANTIENE LA SUA
CONFORMAZIONE
Traslocatori proteici situati
nella membrana
NEL MECCANISMO
2 LA PROTEINA
DEVE
DISTENDERSI
Viaggiano a bordo di vescicole di
trasporto per andare dal RE
all’Apparato di Golgi, endosomi,
lisosomi e superficie cellulare
Mitocondrio
• Il genoma mitocondriale contiene 17.000 coppie di basi e possiede 37
geni codificanti per due RNA ribosomiali (rRNA), 22 RNA di trasporto
(tRNA) e 13 proteine che fanno parte dei complessi enzimatici
deputati alla fosforilazione ossidativa.
• In ogni mitocondrio si trovano da due a dieci copie del genoma
circolare.
• Il resto delle proteine presenti nel mitocondrio deriva da geni nucleari
i cui prodotti vengono appositamente trasportati. Le proteine
destinate al mitocondrio generalmente vengono riconosciute grazie
ad una sequenza leader presente sulla loro parte N-terminale.
• Tale sequenza contiene da 20 a 90 amminoacidi, di cui nessuno
carico negativamente, con all'interno alcuni motivi ricorrenti, e sembra
che abbia un'elevata possibilità di dare origine ad una α-elica
anfipatica.
Figure 1-8b Essential Cell Biology (© Garland Science 2010)
Figure 1-18 Essential Cell Biology (© Garland Science 2010)
Per entrare nei mitocondri dal citosol le proteine perdono
la conformazione originale
Peptidasi e proteine secondatrici
scindono la sequenza segnale e
ridanno alla proteina la sua forma
TUTTI QUESTI
PROCESSI
NECESSITANO DI
ENERGIA
NEI MECCANISMI 1 E 3
LA PROTEINA
MANTIENE LA SUA
CONFORMAZIONE
Macromolecole trasporto attivo
Piccole molecole per diffusione
Traslocatori proteici situati
nella membrana
NEL MECCANISMO 2
LA PROTEINA DEVE
DISTENDERSI
Viaggiano a bordo di vescicole di
trasporto per andare dal RE
all’Apparato di Golgi, endosomi,
lisosomi e superficie cellulare
Reticolo endoplasmatico
•
•
•
•
•
•
Il Reticolo Endoplasmatico è il sistema di membrane più esteso delle cellule
eucariotiche (20-30 volte quello della membrana plasmatica)
Il RE è una struttura cellulare costituita da sacche e invaginazioni adese
senza soluzione di continuità alla membrana nucleare. Lo spazio interno
che si identifica tra le pieghe del reticolo è detto lume e presenta una serie
di enzimi che catalizzano diverse reazioni chimiche.
Il termine reticolo si riferisce alla sua forma articolata nello spazio, il termine
endoplasmatico sta a indicare che si trova all'interno (endo) del citoplasma.
Il RE presente all'interno delle fibre muscolari prende il nome di "Reticolo
sarcoplasmatico liscio" e "Reticolo sarcoplasmatico rugoso" a seconda della
propria composizione.
Vi è una distinzione tra reticolo endoplasmatico rugoso (RER) e reticolo
endoplasmatico liscio (REL).
Entrambi i reticoli hanno come compiti quelli di modificare molecole
precedentemente sintetizzate in altri distretti.
I ribosomi
Cosa sono
• I ribosomi (al singolare ribosoma o ribosomio) sono granuli scuri
presenti nel citosol, sulla membrana nucleare e sul reticolo
endoplasmatico rugoso (RER).
• Sono composti da RNA ribosomiale (r-RNA) e materiale proteico, e la
loro funzione è quella di sintetizzare, partendo da una catena di RNA
messaggero (m-RNA), le proteine.
• Furono messi in evidenza per la prima volta al microscopio elettronico
nel 1953.
I ribosomi
• Il ribosoma della cellula eucariote (fatta eccezione per quelli contenuti
nei mitocondri e nei cloroplasti), invece, è più grande ed ha una
massa molecolare di 4000 kDa, un diametro di 23 nm ed un
coefficiente di sedimentazione di 80 S.
– Anch'esso è composto da due unità (60 S e 40 S) che in
complessivo presentano più di 80 proteine.
I ribosomi
• I ribosomi liberi
– Sono così denominati i ribosomi che si trovano liberi nel citoplasma
cellulare o che fanno parte dei mitocondri o dei cloroplasti.
Generalmente sono deputati alla sintesi di proteine che verranno
rilasciate ed utilizzate nel citoplasma o nella parte interna della
membrana cellulare negli organelli precedentemente citati ed anche nei
perossisomi. Più ribosomi sono in grado di unirsi tra loro, utilizzando
come legame una stessa molecola di RNA messaggero attraverso la
quale si muovono dal codone iniziale fino a quello finale, in direzione 3',
a formare un poliribosoma (o polisoma).
• I ribosomi legati alle membrana
– Si trovano legati alle membrane costituenti il nucleo cellulare od il
reticolo endoplasmatico rugoso. Si occupano di sintetizzare e rilasciare
proteine all'interno delle membrane di queste strutture, dove poi
saranno condotte alla loro destinazione finale che può essere sia
intra- che extracellulare (nel qual caso vengono spesso espulse per
esocitosi). È da segnalare che i ribosomi attaccati alle membrane sono
una sezione dell'insieme dei ribosomi liberi, la quale viene poi reclutata
non appena iniziano a sintetizzare proteine specifiche.
Reticolo endoplasmatico
•
I ribosomi citosolici vengono indirizzati al RE se la proteina che stanno
sintetizzando presenta una sequenza segnale per il RE, riconosciuta nel
citosol da una particella di riconoscimento del segnale (SRP): il legame tra il
complesso ribosoma-SRP e un recettore posto sulla membrana del RE
innesca il processo che guida il polipeptide in allungamento oltre la
membrana del RE attraverso un apposito canale di traslocazione.
•
Le proteine solubili destinate alla secrezione o al volume interno di un
organello passano completamente nel lume del RE, mentre le proteine
transmembrana destinate alla membrana del RE o ad altre membrane
cellulari restano ancorate al doppio strato lipidico dalle eliche α che lo
attraversano uno o più volte.
Un unico parco di ribosomi serve a sintetizzare le proteine destinate a
rimanere nel citosol e quelle da trasportare agi organelli delimitati da
membrane (RE compreso
L’SRP (signal recognition particle) è presente nel citosol e si lega alla
sequenza segnale per il RE, un recettore per SRP immerso nella membrana
del RE riconosce la SRP e rallenta la sintesi proteica, il canale di
traslocazione risucchia la catene polipeptidica nella membrana e la trasloca
dall’altro lato, la sequenza segnale viene poi tagliata da una peptidasi
Le sequenze di trasferimento e di arresto determinano la formazione di
una proteina transmembrana
Le sequenze di trasferimento e di arresto determinano la formazione di
una proteina transmembrana
• La maggior parte delle proteine che entrano nel RE va incontro a
modificazioni chimiche:
• Ponti disolfuro stabilazzano la struttura (Cys-Cys)
• Glicoproteine (glicosilazione) la catena laterale oligosaccardiica si attacca
come blocco unico di 14 zuccheri ad una Asp poi vengono modificate per
dare forma finale sia nel RE che nel Golgi
Le proteine secondatrici (chaperon) trattengono nel lume del RE proteine mal
conformate o assemblate in modo incompleto
Se le proteine mal conformate non riescono ad assumere la struttura
tridimensionale giusta vengono trasportate nel citosol e demolite
Unfolded protein response
Il trasporto vescicolare
Ogni vescicola di trasporto deve portare con se
solo le proteine necessarie al destinatario e deve
fondersi soltanto con la membrana bersaglio
appropriata
Le vescicole mostrano un rivestimento sulla
faccia rivolta verso il citosol (vescicole rivestite)
che conferisce la conformazione a gemma e
contribuisce ad inglobare le molecole da
trasportare
Tipo di vescicola
rivestita
Proteina di
rivestimento
Origine
Destinazione
Clatrina
Clatrina + adaptina 1
Apparato di Golgi
Lisosomi (via
endosomi)
Clatrina
Clatrina + adaptina 2
Membrana plasmatica
endosomi
COP (coat protein)
Proteine COP
RE
cisterna di Golgi
apparato di Golgi
Apparato di Golgi
cisterna di Golgi
RE
Le proteine v-SNARE (vesicle) e t-SNARE (target) svolgono un ruolo fondamentale
nel guidare le vescicole di trasporto alle membrane bersaglio
(ogni organello ed ogni tipo di trasporto ha la usa SNARE)
Soluble NSF (N-ethylmaleimide Sensitive Fusion proteins) Attachment REceptor
Il trasporto vescicolare-esocitosi
• Una via secretoria maestra (rosso) diretta verso l’esterno arriva alla superficie
cellulare partendo dalla regione della biosintesi e della traslocazione proteica
nella membrana del RE attraverso l’apparato di Golgi
• Una via endocitica maestra (verde) diretta verso l’interno preposta l’ingestione e
degradazione di molecole extracellulari porta materiale dalla membrana
plasmatica ai lisosomi passando per gli endosomi
Entrata (verso il citosol)
Uscita (verso la membrana cellulare)
Sacche appiattite delimitate da membrane dette cisterne (o dittiosomi).
Ogni pila (insieme di cisterne a forma di sacca) contiene da 3 a 20 cisterne.
Le pile vanno da 1 a qualche centinaio, dipende dal tipo di cellule.
Lato di ingresso nella pila (lato cis adiacente al RE), lato di uscita dalla pila
(lato trans rivolto verso la membrana plasmatica.
Esocitosi
•
•
•
•
•
•
L’apparato di Golgi riceve proteine di nuova fabbricazione provenienti dall’RE, ne modifica
la parte oligosaccaridica, le smista e le spedisce dal reticolo di Golgi alla membrana
plasmatica, ai lisosomi o alle vescicole secretorie.
In tutte le cellule eucariotiche dal reticolo di Golgi gemmano continuamente vescicole di
trasporto, che vanno a fondersi con la membrana plasmatica in un processo denominato
esocitosi costitutiva.
In tal modo pervengono alla superficie cellulare i rifornimenti di lipidi e proteine di
membrana e si riversano anche molecole fuori dalla cellula: in questo ultimo caso si parla
di secrezione.
Le cellule specializzate nella secrezione hanno anche un percorso di esocitosi regolata,
nel quale le molecole, immagazzinate in apposito vescicole secretorie, si liberano fuori
dalla cellula per esocitosi quando la cellula riceve il segnale di secernere.
Le cellule assumono liquidi, molecole e talvolta anche particelle per endocitosi, nel corso
della quale zone di membrana plasmatica si invaginano e si distaccano formando vescicole
endocitiche.
Gran parte del materiale internalizzato per endocitosi passa negli endosomi e quindi nei
lilsosomi, che lo digeriscono con i loro enzimi idrolitici; invece i componenti della membrana
vescicolare vengono in gran parte riciclati in vescicole di trasporto che tornano alla
membrana plasmatica.
Esocitosi costitutiva ed esocitosi regolata (ormoni, muco , enzimi digestivi)
Esocitosi regolata
(aumenti ematici di glucosio inducono la secrezione dell’ormone insulina)
Nelle vescicole secretorie regolate le proteine si aggregano e quindi possono trovarsi in
grandi quantità cosa che non avviene nelle vescicole costitutive
Endocitosi
• Endocitosi: assunzione di liquidi e molecole dall’esterno
• Due tipi di endocitosi:
- Pinocitosi (parti di mambrana plasmatica
e liquidi <150nm)
- Fagocitosi (particelle di grandi
dimensioni >250 nm-microorganismi, detriti, a
carico di cellule specializzate)
• Cellule intestinali (grosse particelle di cibo)
• Cellule fagocitiche del sangue: macrofagi e neutrofili.
I batteri espongono dei recettori riconosciuti da anticorpi.
I batteri ricoperti da anticorpo vengono fagocitati da
macrofagi
Le vie endocitiche e le vescicole endocitiche
Due macrofagi: a destra un macrofago sta fagocitando due eritrociti
Endocitosi mediata da recettori (LDL-colesterolo, Vit. b12, Ferro, HIV)
LDL= low density lipoprotein complessate a colesterolo, idrofobico nel sangue
• Gli endosomi smistano le
molecole internalizzate per
endocitosi (pinocitosi)
• Il comparto endosomico funge
da centrale di smistamento
della via endocitica diretta
verso l’interno (Golgi
smistamento via secretoria,
endosoma smistamento via
endocitica)
• Esistono due gruppi di
endosomi:
• Endosomi precoci subito sotto
la membrana plasmatica
• Endosomi tardivi subito sotto il
nucleo
• Esitono tre vie di destino finale
riciclaggio, degradazione,
transcitosi (1/2/3)
• Il lisosoma contiene enzimi idrolitici ed una pompa per H+
• Degradazione proteine, oligosaccaridi e lipidi
• Proteine di trasmembrana affacciate nel lumen dell’organello
associate a carboidrati complessi per proteggersi dalla denaturazione
• Enzimi lisosomali: complessati in RE e Golgi con mannoso-6-fosfato che con
recettore vengono racchiusi in vescicole e destinati al lisosoma
Per arrivare ai lisosomi i materiali destinati alla degradazione fanno percorsi diversi
Fagocitosi
Pinocitosi
Autofagia (degradazione di parti consumate della cellula)