Premessa
Ho trovato del materiale utile a sostenere il progresso dell’ipotesi naturalistica di Freud e
della scuola a cui egli in sostanza ha sempre aderito (almeno da quanto si capisce dalla tua
tesi, io non sono un conoscitore di Freud). Di Crick (quello che scoprì la doppia elica con
Watson nel 1953, quando ancora qualcuno pensava che la trasmissione dei caratteri ereditari
fosse legata a cause trascendentali e religiose e che non si sarebbe mai potuta comprendere)
avevo anche il libro “The astonishing hypotesis”, cioè “l’ipotesi sorprendente” nel quale
Crick, nonostante l’età avanzata, illustra la sua missione, le sue idee, e le evidenze
sperimentali alla base dell’ipotesi che la coscienza visiva sia descrivibile come uno stato
della rete di neuroni di una rilevante parte del cervello, così come il modo in cui siamo fatti,
cresciamo e ci sviluppiamo con i nostri organi al loro posto, derivano da sequenze di
molecole ATGC nel nostro DNA. È evidente che la missione indicata da Crick va nella
direzione di spiegare nello stesso modo gli stati coscienti generali dell’uomo, ovvero, perché
abbiamo una consapevolezza del tutto che ci circonda e anche di noi stessi? La ricerca fa ben
sperare che la scienza impiegherà poco tempo per dare una risposta sperimentale a questo
interrogativo, al di là delle beghe e dei pregiudizi sul materialismo-antimaterialismo.
Eccoti dunque del materiale che sarai libera di organizzare.
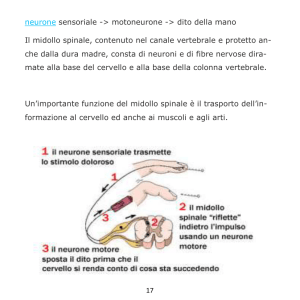
6. Integrazione cerebrale e coscienza.
Gli studi sperimentali sulla organizzazione del sistema nervoso centrale sono stati portati
avanti principalmente da Roger W. Sperry (1913). Subito dopo il dottorato in zoologia,
ottenuto con P.A. Weiss, Sperry si interessò agli effetti sulla coordinazione motoria di
alterazioni delle connessioni neuromuscolari e connessioni fra nervi ed organi terminali
provocate da interventi chirurgici. Si poteva osservare che attraverso una serie di sostituzioni
nelle funzioni svolte da alcuni nervi la coordinazione motoria rimaneva inalterata,
permettendo riadattamenti rieducativi. In studi di laboratorio Sperry cercò di individuare
quali meccanismi nervosi centrali permettevano questo riadattamento, ma non riuscì a
riprodurre il fenomeno perché i trapianti di muscoli e di nervi in ratti e scimmie provocavano
profondi e persistenti disturbi motori e sensoriali.
Nel 1945 Sperry svolse un’analisi approfondita di questi risultati che lo spinse a rifiutare
le teorie generalmente accettate sulla plasticità del sistema nervoso centrale. I risultati
sperimentali dimostravano infatti una stretta specificità funzionale dei circuiti fondamentali
del cervello dei vertebrati. Contrariamente alla ipotesi della totale plasticità, che rende il
sistema nervoso per definizione inaccessibile alla sperimentazione, l’ipotesi di una specificità
funzionale apriva un nuovo dominio sperimentale. È possibile infatti costruire circuiti basati
su questa specializzazione e controllarli mediante metodi neurofisiologici.
In alcuni invertebrati, come nelle rane e nei rospi, era possibile ottenere costantemente
delle reintegrazioni funzionali, nonostante drastiche modificazioni delle connessioni nervose.
Per risolvere questo problema Sperry iniziò una sperimentazione sistematica sulla crescita
selettiva delle connessioni nervose durante lo sviluppo e la rigenerazione. In questo ambito
dimostrò, nel 1941, l’esistenza di una stretta correlazione fra percezione visiva e disposizione
anatomica delle connessioni del nervo ottico. Una visione dal basso all’alto, una visione con
relazioni invertite fra davanti e dietro e varie combinazioni fra questi due tipi potevano essere
direttamente collegate all’orientamento dato chirurgicamente all’occhio e alla distribuzione
delle fibre nervose che collegano l’occhio al cervello.
Dopo il 1945 Sperry si dedicò allo studio dell’integrazione nervosa a livello della
corteccia cerebrale nei mammiferi, con l’applicazione delle stesse tecniche microchirurgiche
utilizzate sino ad allora sugli anfibi, mediante sezionamenti a diversi piani della corteccia e
l’inserimento di elementi dielettrici o conduttori nelle varie parti della corteccia in modo da
escludere selettivamente campi elettrici e collegamenti. All’inizio degli anni Cinquanta
Sperry si occupò particolarmente del corpo calloso, per il quale precedenti risultati
sperimentali sembravano dimostrare che l’interruzione di questo grande fascio di fibre
nervose nel cervello non provocava alcuna disfunzione evidente. Questa era naturalmente una
evidenza contraria alle teorie sulla specificità e Sperry, insieme a R. Meyers, utilizzando le
tecniche microchirurgiche e l’attento studio dei comportamenti, riuscì a dimostrare nei gatti
una chiara dissociazione delle funzioni visive in seguito alla interruzione dei collegamenti fra
i due emisferi cerebrali (split-brain). Se si controlla l’afflusso di informazione nel cervello
separatamente per i due emisferi si trova che «ciascuno di essi ha in tali condizioni una sua
propria sfera mentale o sistema cognitivo, cioè processi percettivi, di apprendimento, di
memoria e altri processi mentali propri e indipendenti ... è come se gli animali avessero due
cervelli separati» (1968).
In genere uno dei due emisferi è fortemente dominante e svolge le attività di livello
superiore, mentre l’altro emisfero contribuisce ad una funzione generalizzata di base. La vita
vegetativa richiede una unità dei centri inferiori, mentre i centri superiori possono agire anche
in aperto contrasto oppure più spesso produrre una integrazione fra due diverse strutture, che
mantengono la specificità delle relative competenze e della «storia» di ognuna. Nel cervello
non esiste quindi solo una divisione del lavoro, che è molto flessibile, dato che tutte le
funzioni sono generalizzate o generalizzabii, ma una complessa organizzazione gerarchica,
all’interno di ogni emisfero e tra i due emisferi.
Negli anni successivi Sperry ha sviluppato ricerche su soggetti umani che
erano stati sottoposti alla separazione chirurgica dei due emisferi per ragioni terapeutiche. I
risultati sperimentali mostravano che solo l’emisfero dominante, che nel 98% dei casi è il
sinistro, contenente i centri del linguaggio, mostra una attività autocosciente, mentre
l’emisfero minore è paragonabile al cervello di un animale superiore.
Dopo il recupero parziale e mediato dell’idea della localizzazione dei processi cerebrali, si
è diffusa l’idea di una corrispondenza logica fra forma percepita e una forma che si instaura
nel cervello. Il modello che riflette la realtà non è solo uno schema interpretativo, ma sembra
corrispondere ad urla effettiva organizzazione all’interno del cervello, corrispondenza non
elemento per elemento, ma topologica, qualitativa. Nel cervello è quindi possibile individuare
aree differenziali, che non sono tuttavia completamente separate, ma dispongono di circuiti di
integrazione a diversi livelli.
Anche qui, per tornare al piano del dibattito teorico (come nel caso della morfogenesi) la
dicotomia fra specificità e plasticità poté venire superata, considerando i diversi livelli a cui
si svolgono i fenomeni: da una parte la indeterminazione dei processi microscopici, dall’altra
la presenza di un campo, che limita tale indeterminazione subordinandoia ad un «piano», ad
un programma.
Questi metodi sono stati applicati ai due gruppi in cui è possibile schematicamente
suddividere il complesso delle attività nervose e che vengono chiamati da John C. Eccles
«outer sense» (senso esterno) e «inner sense » (senso interno) (1974). Il primo comprende i
fenomeni percettivi del mondo esterno (luce, colore, suono ecc.) e i dati sullo stato
fisiologico dell’organismo; il secondo è costituito dalle attività psichiche, non direttamente
collegate ai dati percettivi, ma provenienti dall’attività autonoma del cervello: il pensiero, la
memoria, i sogni, l’immaginazione, ecc.
7. La visione e la memoria.
Una particolare attenzione è stata dedicata a partire dagli anni ‘70 allo studio del processo
percettivo legato alla visione, anche perché più facilmente soggetta ad una analisi
sperimentale. In tale processo, meglio che nelle altre attività sensorie, era possibile
individuare il ruolo di modelli nel cervello e delle attività di selezione attiva degli stimoli
luminosi. In questo ambito si sono anche sviluppati programmi integrati fra psicologia,
neuroscienze e cibernetica ed informatica per cercare di simulare il funzionamento del
cervello biologico. I sistemi visivi, come era risultato in linea generale dalle indagini
neurofisiologiche già a partire dalla fine degli anni Venti, usano una gerarchia di
rappresentazioni che sviluppano l’informazione visiva in molti stadi intermedi in modo da
riempire la distanza esistente fra i segnali in ingresso ed i simboli cognitivi. L’esistenza di
diversi sistemi di elaborazione visiva ha fatto avanzare l’idea che i segnali vengano trattati in
modo parallelo in certi casi e sequenziale in altri, a seconda della natura e dell’importanza
delle cose da discriminare visivamente. La visione deve infatti analizzare dati visivi variabili
nel tempo in tutta la loro complessità ed in tempo reale. Inoltre i sistemi biologici usano
elementi di processo neurale che sono di sei ordini di grandezza più lenti dei componenti al
silicio, utilizzati nei calcolatori elettronici. Nonostante ciò il cervello è in grado di rispondere
agli stimoli visuali in poche centinaia di millisecondi.
Il principio fondamentale di costruzione dell’informazione visiva è dunque quello di
rappresentazione gerarchica dei processi percettivi e cognitivi. Questa gerarchia può essere
ritrovata a livello morfologico in aree cerebrali distinte dedicate alla attività visuale, ognuna
delle quali può essere considerata una « mappa » grossolana di una parte del mondo visuale.
Ai livelli superiori della gerarchia queste mappe divengono molto più diffuse e i neuroni
rispondono a caratteri più complessi e diverse aree corticali sono in grado di rispondere a
caratteri diversi, come il colore e il movimento. Nonostante questa grande frammentazione
dei modi di risposta, una immagine è un tutto unitario; devono quindi esistere meccanismi di
integrazione.
Nella seconda metà degli anni Settanta A. Treismann e i suoi collaboratori hanno
realizzato una serie di esperimenti nei quali si presentavano a dei soggetti forme colorate da
riconoscere con tempi molto rapidi ed in presenza di vari « distrattori ». Se si chiede di
riconoscere una 5 in mezzo a un insieme casuale di lettere X verdi o I marroni, la risposta è
molto rapida. Tuttavia se si chiede di individuare una T verde, il che richiede di riconoscere
al tempo stesso una data forma ed un dato colore, il processo richiede molto più tempo. Per
spiegare questo fenomeno è stata introdotta l’ipotesi che il cervello analizzi gli oggetti in
serie, spostandosi su un oggetto dopo l’altro, con una velòcità calcolata in 70 millisecondi per
ogni passo. Il processo èstato equiparato per analogia a quello della illuminazione con un
riflettore in un ambiente poco illuminato di un oggetto alla volta.
Treisman ha proposto che questo meccanismo funzioni attraverso la formazione di «
giunzioni » temporanee. di neuroni. Nel 1981 von der Malsburg ha proposto che questo
avvenga per mezzo del rafforzamento di sinapsi esistenti, in modo da poter creare in tempi
brevissimi una nuova linea nervosa. In questo modo si introduce una distinzione fra
alterazioni di lungo termine dovuto all’apprendimento e alterazioni transienti che
avverrebbero durante l’atto della percezione.
Nel 1981 F.H.C. Crick, con il consueto ed innovativo rigore teorico, ha sviluppato queste
idee, affermando che l’ipotesi di von der Malsburg rappresenta una rottura con le concezioni
tradizionali sulle reti di neuroni1, basate sul fatto che le sinapsi hanno caratteri costanti
Le reti neurali sono dei programmi per l’autoapprendimento dei computer. In tali programmi esiste una serie di
“automi”, o “cellule” virtuali, capaci di ricevere inviare stimoli (1 o 0) a secondo del loro particolare stato. Le
cellule sono divise in diversi strati, di cui il più esterno è quello che riceve gli stimoli provenienti dall’ambiente
(come quando scriviamo una domanda alla tastiera, o l’occhio della telecamera); gli strati interni elaborano in
maniera apparentemente casuale tali stimoli, propagandoli orizzontalmente e verticalmente, fino ad arrivare allo
strato più interno che produce dei segnali di risposta (es. una risposta a una domanda, o una traduzione di una
frase, o un nome viene assegnato all’immagine di input) da inviare all’esterno. Le prime “risposte” fornite dalla
rete neurale sono completamente assurde. L’insegnante (può essere un’altra macchina che ha in memoria le
risposte esatte, per questioni di rapidità) fornisce delle valutazioni alle risposte date dalla rete neurale. Queste
1
mentre sono in funzione, e possono variare solo durante un processo di apprendimento,
mentre le sinapsi ipotizzate da von der Malsburg si devono modificare.
Analizzando la struttura del talamo e del complesso reticolare che vi è collegato e le
differenti azioni inibitorie od eccitatorie, nel 1984 F.H.C. Crick ha introdotto alcune ipotesi
per spiegare questo fenomeno: 1) il «proiettore »è controllato dai complesso reticolare del
talamo; 2) l’espressione del proiettore è la produzione di scariche rapide in un sottoinsieme
dei neuroni talamici attivi; 3) le congiunzioni prodotte dal proiettore sono mediate da
«sinapsi di von der Malsburg».
Dato che un neurone si può scaricare per effetto di un numero molto elevato di possibili
combinazioni degli stimoli differenti ai quali è sottoposto e quindi la sua risposta è aspecifica
e non può essere direttamente legata ad un dato stimolo, era una idea tradizionale della
neurofisiologia che questa ambiguità venisse rimossa prendendo in considerazione gruppi di
cellule. L’alternativa che ogni neurone abbia la sua specificità sarebbe infatti molto meno «
economica». Questo insieme di cellule erano state definite ma bisognerebbe concordare tutto
con «insieme » « reti associative » e molto lavoro sperimentale è stato fatto su di esse. Queste
reti sono costituite da neuroni dello stesso tipo e vengono perciò definite, da Crick,
aggregazioni orizzontali. L’idea di von der Malsburg permette tuttavia un altro tipo di
aggregazione, verticale, tra diversi livelli di integrazione dei neuroni, associazioni
temporanee che permettono di collegare fra di loro i diversi aspetti di un oggetto. Così, un
neurone di livello superiore capace di indicare l’idea generale di « viso » potrebbe associarsi
con neuroni di livello inferiore, che indicano parti del viso od alcuni caratteri particolari. Di
qui una quarta ipotesi introdotta da Crick e cioè che « le congiunzioni sono espresse da
aggregazioni cellulari, aggregazioni specialmente tra cellule di differenti regioni corticali».
Esiste quindi una interrelazione fra la precisione di molte connessioni nervose e
l’esistenza di reti associative, la cui importanza relativa cambia a seconda de] grado di
organizzazione dei sistemi viventi. Nel caso di Caenorhabditis elegans, studiato da Brenner, i
279 neuroni di questo organismo elementare sono collegati sempre esattamente nella stessa
maniera. Negli organismi superiori queste connessioni sono molto meno precise, ma non
casuali e la forza di queste associazioni dipende dall’«esperienza». Per assicurare la
percezione il cervello utilizza due strategie: mappature successive e multiple basate su
connessioni precise e l’esistenza di reti associative all’interno di piccole regioni in cui tutto è
collegato con tutto. All’inizio del trattamento dei segnali, uno acustico ed uno visivo ad
esempio, questi non sono collegati fra loro. Ma presto nel passaggio da una mappa a quella di
livello superiore si crea una integrazione sempre maggiore. Il cervello quindi non è una
immensa rete, ma un insieme di reti locali, alcune in parallelo, altre arrangiate serialmente.
Questo problema esemplifica in modo significativo le linee di ricerca che si sono
sviluppate a partire dagli anni ‘70. In esse si è stabilita una stretta interazione fra ricerche di
tipo molto diverso: l’indagine anatomico-morfologica, quella molecolare, quella psicologica.
Questa convergenza multidisciplinare indica anche la centralità che lo studio del sistema
nervoso e dei cervello ha acquisito negli ultimi decenni. In un articolo del 1979 Crick ha
paragonato lo stato problematico della neurobio]ogia alla fine degli anni ‘70 a quello della
risposte, positive o negative, innestano una retroazione che stimola il cambiamento delle “regole” con cui le
singole cellule rispondono agli stimoli in funzione dello stato in cui si trovano (ma non modificano le
connessioni di ogni cellula con le cellule vicine come avviene nel cervello). Questi cambiamenti sono
completamente casuali, ma interessano maggiormente quelle cellule che hanno portato a generare errori nella
risposta finale e stabilizzano quelle che hanno generato risposte corrette. Dopo molti tentativi ed errori il sistema
riesce a incrementare i punteggi medi che riceve dall’insegnante, vale a dire inizia a dare risposte sempre più
corrette, ovvero apprende. Una volta finita la parte dell’addestramento, ottimizzate e fissate tutte le regole di
comunicazione tra le cellule, il sistema è pronto per rispondere a domande (chiaramente sullo stesso settore in
cui è stato “addestrato”) anche senza voti, premi o punizioni. Ciò corrisponde più o meno quanto fa il traduttore
di Google.
genetica e della biochimica alla fine degli anni ‘40, prima della rivoluzione molecolare: «ciò
che chiaramente manca è una ampia struttura di idee con la quale integrare i differenti modi
di affrontare il problema». Ciò che manca, allora, è una « teoria generale del cervello) »,
basata sulle analisi di tipo anatomico-morfologico, psicologico, biochimico e biofisico e sulla
teoria matematica della comunicazione. La speranza è quella di trovare per la neurobiologia
ciò che la doppia elica era stata per la genetica, un meccanismo semplice alla base della
complessa macchina « barocca » del sistema nervoso.
Fonte: da La Neurobiologia, di Bernardino Fantini, in Storia della Scienza, a cura di Paolo
Rossi, vol. 8, gruppo editoriale l’Espresso, 2006.
La nota sulle reti neurali, le evidenziazioni e la premessa sono mie (a.t.)