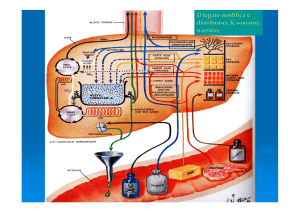
La gluconeogenesi
• La sintesi netta di glucosio, o la formazione di questo a
partire da una grande varietà di molecole che non sono
carboidrati è detta gluconeogenesi, una via metabolica che
utilizza come fonti di carbonio vari aminoacidi, lattato,
piruvato, propionato e glicerolo.
1
•La biosintesi del glucosio è una necessità assoluta
nei mammiferi, in quanto il cervello, il sistema
nervoso, la parte midollare del rene, i testicoli, gli
eritrociti e i tessuti embrionali utilizzano il glucosio
presente nel sangue come unica o principale
sostanza nutriente.
Il cervello umano consuma oltre 120 g di glucosio al
giorno
2
La gluconeogenesi permette il mantenimento
dei livelli di glicemia per molto tempo dopo la
3
completa utilizzazione di tutto il glucosio
La formazione di glucosio da precursori non
saccaridici è chiamata gluconeogenesi
(produzione di nuovo zucchero).
La gluconeogenesi è una via universale, identificata negli
animali, nelle piante, nei funghi e nei microrganismi. In tutti
i casi, le reazioni sono sempre le stesse.
Anche se le reazioni della gluconeogenesi sono le stesse in
tutti gli organismi, il contesto metabolico e la regolazione
della via differiscono da organismo a organismo e da tessuto
a tessuto.
4
I precursori del glucosio negli animali sono il lattato, il
piruvato, il glicerolo e alcuni amminoacidi (Fig. 20.1)
Negli animali superiori la gluconeogenesi avviene
prevalentemente nel fegato e in piccola parte nella corteccia
renale; il glucosio prodotto passa poi nel sangue per
rifornire gli altri tessuti.
5
6
Come la conversione glicolitica del glucosio in piruvato è
la via fondamentale del catabolismo dei carboidrati,
la conversione del piruvato in glucosio è una via
biosintetica essenziale.
Negli amimali, entrambe le vie avvengono
fondamentalmente nel citosol e necessitano di una
regolazione reciproca e coordinata. Le due vie non sono
identiche, anche se condividono diverse tappe (Fig. 20.2).
Sette delle reazioni enzimatiche della gluconeogenesi
sono l'inverso di reazioni della glicolisi
7
8
Tre tappe della glicolisi sono essenzialmente
irreversibili in vivo e non possono essere
utilizzate nella gluconeogenesi:
•
la conversione del glucosio in glucosio 6-fosfato
da parte dell'esochinasi,
•
la fosforilazione del fruttosio 6-fosfato a
fruttosio 1,6-bisfosfato da parte della
fosfofruttochinasi-1,
•
la conversione del fosfoenolpiruvato in piruvato
da parte della piruvato chinasi (Fig. 20.2).
Nelle cellule queste tre reazioni hanno una variazione di
energia libera, ∆G, fortemente negativa, mentre le
altre sette reazioni hanno un valore di ∆G vicino a 0
(Tabella 20.1).
9
10
•Queste tre tappe sono superate mediante un
gruppo diverso di enzimi, che catalizzano
reazioni diverse con, ovviamente, equilibri
diversi.
• Questi enzimi operano nella gluconeogenesi,
ma non nella glicolisi, e le loro reazioni sono
praticamente irreversibili nella direzione della
sintesi del glucosio.
•La glicolisi e la gluconeogenesi sono regolate
indipendentemente mediante controlli
esercitati su specifiche reazioni enzimatiche
che non sono comuni alle due vie.
11
Il fatto che la via che porta dal piruvato al PEP
attraversi anche i mitocondri non è certo casuale.
Il rapporto [NADH]/[NAD+) nel citosol è di 8 x 10-4,
circa 105 volte più basso di quello nei mitocondri.
Poiché il NADH viene consumato dalla gluconeogenesi
(nella
conversione
dell'I,3-bisfosfoglicerato
a
gliceraldeide 3-fosfato; Fig. 20.2), la biosintesi del
glucosio non può procedere se non è continuamente
disponibile NADH.
12
La conversione del piruvato in fosfoenolpiruvato richiede due
reazioni esoergoniche
La prima «deviazione» nella gluconeogenesi è la
conversione del piruvato in fosfoenolpiruvato.
Questa reazione non può avvenire invertendo la
reazione catalizzata dalla piruvato chinasi della
glicolisi , che ha una variazione di energia libera
molto negativa ed è irreversibile nelle condizioni
intracellulari (Tabella 20.1).
La fosforilazione del piruvato avviene mediante una
sequenza di reazioni che nei mammiferi, e in qualche
altro organismo, richiede la partecipazione di enzimi
13
sia dei mitocondri sia del citosol.
Il piruvato viene prima trasportato dal citosol nei
mitocondri oppure viene prodotto sempre nei mitocondri
dall'alanina per transamminazione.
La piruvato carbossilasi, un enzima mitocondriale che
richiede biotina come cofattore, converte poi il piruvato in
ossalacetato (Fig. 20.3):
Piruvato + HCO3- + ATP ossalacetato + ADP +
Pi
La piruvato carbossilasi è il primo enzima regolatore della
via gluconeogenetica;
l'acetil-CoA è un modulatore positivo dell'enzima.
Questa è anche una reazione anaplerotica in quanto
rifornisce di intermedi il ciclo dell'acido citrico.
Il meccanismo della reazione coinvolge la biotina come
14
trasportatore di HCO3-;).
Piruvato carbossilasi:
Biotina come gruppo prostetico
PEPCK
15
Struttura a domini della piruvato carbossilasi.
Il dominio di legame per l’ATP attiva lo ione HCO3- e trasferisce CO2 al dominio
di legame per la biotina. Da qui CO2 viene trasferito al piruvato nel dominio
centrale.
La biotina è su un “guinzaglio”
flessibile che le permette di muoversi
tra il sito dell’ATP e del bicarbonato e il
sito del piruvato.
16
17
L'ossalacetato formato dal piruvato direttamente nei
mitocondri viene ridotto reversibilmente a malato dalla
malato deidrogenasi mitocondriale a spese del
NADH: Ossalacetato + NADH + H+ L-malato + NAD+
Questa reazione, a giudicare dalla sua ∆G'o, è
altamente esoergonica. Ma, in condizioni fisiologiche,
la reazione ha un ∆G=0 ed è quindi reversibile.
La malato deidrogenasi mitocondriale perciò
funziona sia nella gluconeogenesi sia nel ciclo
dell'acido citrico, anche se l'intero flusso di metaboliti,
nei due processi, ha direzione opposta.
18
Il malato esce dai mitocondri attraverso il
trasportatore malato-α-chetoglutarato presente
nella membrana mitocondriale interna.
Nel citosol, il malato viene riossidato a
ossalacetato, con la contemporanea produzione
di NADH citosolico:
Malato + NAD+
ossalacetato + NADH + H+
19
Trasporto dell’ossalacetato
dal mitocondrio al citosol
20
L’ossalacetato
viene
poi
convertito
in
fosfoenolpiruvato (PEP) dalla fosfoenolpiruvato
carbossichinasi in una reazione in cui gli ioni Mg2+
sono cofattori essenziali e il donatore del gruppo
fosforico è il GTP (Fig. 20.3):
Ossalacetato + GTP fosfoenolpiruvato + CO2 +
GDP
Nelle condizioni intracellulari questa reazione è
reversibile; la formazione di un composto fosforilato ad
alta energia (PEP) è bilanciata dall'idrolisi di un altro
21
composto ad alta energia (GTP).
Meccanismo di reazione della PEPCK
L’ossalacetato perde CO2 e l’ossigeno attacca il gruppo γ− fosforico del GTP
con la formazione di PEP e GDP.
22
La reazione complessiva di questa deviazione, è:
Piruvato + ATP + GTP + HCO3- fosfoenolpiruvato + ADP + GDP + Pi + CO2
∆G'o = 0,9 kJ/mole
23
La CO2 persa nella reazione della PEP carbossichinasi è la
stessa molecola che era stata aggiunta al piruvato nella
reazione della piruvato carbossilasi.
Questa sequenza di reazioni di carbossilazione e di
decarbossilazione rappresenta un sistema di «attivazione»
del piruvato, in quanto la decarbossilazione
dell'ossalacetato facilita la formazione del PEP.
Un sistema di attivazione simile viene usato per attivare
l'acetil-CoA nella biosintesi degli acidi grassi
24
Il trasporto del malato dai mitocondri al citosol e la
sua riconversione in ossalacetato ha come effetto
anche lo spostamento di equivalenti riducenti sotto
forma di NADH nel citosol, dove questo composto
tende a scarseggiare.
La via dal piruvato al PEP serve anche a bilanciare la
produzione e il consumo di NADH nel citosol durante la
gluconeogenesi.
25
Una seconda «reazione» piruvato PEP, più
breve,
diventa
predominante
quando
il
precursore della gluconeogenesi è il lattato (Fig.
20.4).
In questa via viene utilizzato il lattato prodotto
dalla glicolisi negli eritrociti o nel muscolo, in
particolare nei vertebrati di grandi dimensioni
dopo un esercizio fisico prolungato.
La conversione del lattato in piruvato nel
citosol degli epatociti genera NADH, e quindi
non è più necessaria l'esportazione di malato
dai mitocondri.
26
•Il piruvato prodotto nella reazione della lattato
deidrogenasi viene trasportato all'interno dei
mitocondri, dove viene trasformato in
ossalacetato dalla piruvato carbossilasi.
•L'ossalacetato viene convertito in PEP
direttamente nei mitocondri ad opera di una
forma mitocondriale di PEP carbossichinasi.
•Il prodotto di questa reazione esce dai
mitocondri per entrare nella via
gluconeogenetica.
27
Il promotore del gene per la PEPcarbossichinasi
Gli ormoni influenzano l’espressione genica variando la velocità di trascrizione.
IRE = elemento di risposta all’insulina
GRE = elemento di risposta ai glucocorticoidi
TRE = elemento di risposta all’ormone tiroideo
CRE I e CRE II = elementi di risposta all’cAMP
28
Le forme citosolica e mitocondriale di
PEP carbossichinasi sono codificate da
geni nucleari diversi.
Questo è un altro esempio di enzimi
distinti che, pur catalizzando la stessa
reazione, hanno localizzazioni cellulari e
funzioni metaboliche diverse
29
Vie alternative da piruvato a
fosfoenolpiruvato.
La via che prevale dipende dalla
natura chimica del precursore
della gluconeogenesi (lattato o
piruvato).
La via indicata sulla destra (che
è più breve di quella indicata a
sinistra) prevale quando il suo
precursore è il lattato, in quanto
il NADH citosolico viene prodotto
nella reazione catalizzata dalla
lattato deidrogenasi e non è
quindi necessario trasportarlo
fuori dal mitocondrio.
Fig. 20.4
L’importanza relativa delle due
vie dipende dalla disponibilità di
lattato e dalla richiesta di NADH
citosolico
necessario
per
la
30
gluconeogenesi.
La conversione del fruttosio 1,6-bisfosfato in
fruttosio 6-fosfato è la seconda deviazione.
La seconda reazione della sequenza catabolica
glicolitica che non partecipa al processo
anabolico della gluconeogenesi è la
fosforilazione del fruttosio 6-fosfato catalizzata
dalla fosfofruttochinasi-1 (Tabella 20.1).
31
Nella cellula questa reazione è altamente
esoergonica
e
quindi
irreversibile,
la
formazione di fruttosio 6-fosfato da fruttosio
1,6-bisfosfato (Fig. 20.2) è catalizzata da un
altro enzima,
la fruttosio 1,6-bisfosfatasi Mg2+-dipendente,
che
produce
l'idrolisi
essenzialmente
irreversibile del gruppo fosforico sul C-1 (non
il trasferimento del gruppo fosforico all'ADP):
Fruttosio 1,6-bisfosfato + H2O fruttosio 6fosfato + Pi
∆G'o = - 16,3 kJ/mole
32
La conversione del glucosio 6-fosfato in glucosio
libero è la terza deviazione
La terza deviazione è la reazione finale della
gluconeogenesi, la defosforilazione del glucosio
6-fosfato a glucosio libero (Fig. 20.2).
Poiché la reazione dell'esochinasi nella glicolisi è
essenzialmente
irreversibile,
la
reazione
idrolitica è catalizzata da un altro enzima, la
glucosio 6-fosfatasi:
Glucosio 6-fosfato + H2O
glucosio + Pi
∆G'o=-13,8 kJ/mole
33
TERZA DEVIAZIONE
Glucosio 6-fosfato + H2O
glucosio + Pi
La glucosio 6-fosfatasi si trova soltanto nei tessuti che hanno il
compito metabolico di mantenere l’omeostasi del glucosio ematico:
fegato e in minor misura rene.
34
Questo enzima, la cui attività dipende dalla presenza
di ioni Mg2+, si trova nel reticolo endoplasmatico
degli epatociti e nelle cellule renali.
La glucosio 6-fosfatasi non è presente nel muscolo o
nel cervello, e quindi la gluconeogenesi non può
avvenire in questi tessuti.
Il glucosio prodotto dalla gluconeogenesi nel fegato o
nei reni o ingerito con la dieta viene trasportato al
muscolo e al cervello dal flusso sanguigno.
35
Sintesi di glucosio a partire dal glucosio 6-fosfato (reticolo endoplasmatico)
5 proteine per trasformare il glucosio 6-P citosolico in glucosio:
Glucosio 6-fosfatasi associata al RE
SP= proteina stabilizzante associata alla glucosio 6-fosfatasi
T1= trasporta glucosio 6-fosfato nel lume del RE
T2 e T3= trasportano a ritroso nel citosol rispettivamente Pi e glucosio
36
37
La gluconeogenesi è energeticam ente costosa
2 Piruvato + 4 ATP + 2 G TP + 2 N AD H + 4 H 2O
glucosio + 4 AD P + 2 GD P + 6 Pi + 2 N AD + + 2 H +
G lucosio + 2 AD P + 2 Pi + 2 NAD +
glicolisi
2 piruvato + 2 ATP + 2 N AD H + 2 H + + 2 H 2 O
38
La gluconeogenesi è energeticamente
costosa
Per ogni molecola di glucosio che si forma dal piruvato
vengono consumati sei legami fosforici ad alta energia,
quattro ricavati dall'ATP e due dal GTP.
Sono inoltre necessarie due molecole di NADH per la
riduzione di due molecole di 1,3-bisfosfoglicerato.
Questa reazione complessiva non è semplicemente
l'inverso della reazione della glicolisi, che converte
glucosio in piruvato producendo due molecole di ATP:
Glucosio + 2 ADP + 2 Pi + 2 NAD+ 2 piruvato + 2 ATP +
2 NADH + 2 H+ + 2 H2O
39
Molta dell'energia libera necessaria per rendere
possibile la gluconeogenesi rende la gluconeogenesi
stessa un processo irreversibile.
Nelle condizioni intracellulari, la variazione di energia
libera complessiva della glicolisi è di circa -63 kJ/mole.
Nelle stesse condizioni intracellulari, la variazione di
energia libera complessiva della gluconeogenesi è -16
kJ/mole.
In sostanza, sia la glicolisi sia la
gluconeogenesi sono processi essenzialmente
irreversibili nelle condizioni esistenti nella
cellula.
40
La gluconeogenesi è favorita
quando la cellula è ricca di
precursori biosintetici e di ATP.
41
42
Gli intermedi del ciclo dell'acido citrico e molti
amminoacidi sono glucogenici
La via biosintetica descritta in precedenza
consente una sintesi netta di glucosio non
solo dal piruvato, ma anche dagli intermedi
del ciclo dell'acido citrico:
1. citrato, isocitrato,
2. -α
α-chetoglutarato,
3. succinil-CoA,
4. succinato,
5. fumarato
6. malato
Tutti questi composti vengono ossidati nel
ciclo dell'acido citrico e sono trasformati in
ossalacetato.
43
Alcuni degli atomi di carbonio di molti
amminoacidi derivati dalle proteine sono
convertiti nelle cellule dei mammiferi sia in
piruvato sia in alcuni intermedi del ciclo
dell'acido citrico.
Dopo il distacco dei loro gruppi amminici nei
mitocondri epatici, lo scheletro carbonioso di
questi amminoacidi (rispettivamente i
chetoacidi piruvato ed α-chetoglutarato) è
incanalato nella gluconeogenesi.
44
Questi amminoacidi
possono essere
utilizzati per la
produzione di
glucosio e sono
chiamati glucogenici
(Tabella 20.3).
L'alanina e la
glutammina
sono particolarmente
importanti in quanto
sono le molecole che
trasportano i gruppi
amminici dai tessuti
extraepatici al fegato
** Questi amminoacidi sono anche chetogenici
45
La gluconeogenesi e la glicolisi sono
regolate in modo reciproco
Per assicurarsi che i cicli futili non avvengano nelle
condizioni normali, la gluconeogenesi e la glicolisi
devono essere regolate separatamente e con sistemi
integrati e complementari.
Il primo punto di controllo è rappresentato dalle
reazioni catalizzate dal complesso della piruvato
deidrogenasi e dalla piruvato carbossilasi della
gluconeogenesi (Fig. 20.6)
46
1. L'acetil-CoA
da una parte è il modulatore positivo
allosterico della piruvato carbossilasi
2. E’ il modulatore negativo della piruvato
deidrogenasi
3. Il
controllo si verifica mediante la stimolazione di
una proteina chinasi che inattiva la deidrogenasi.
47
48
•Quando le richieste energetiche della cellula
sono soddisfatte, la fosforilazione ossidativa
rallenta, il NADH non viene più consumato e il
ciclo dell'acido citrico è inibito, provocando
come conseguenza un accumulo di acetil-CoA.
• L'aumento della concentrazione di acetil-CoA
inibisce il complesso della piruvato
deidrogenasi, rallentando la sua formazione da
piruvato,
•contemporaneamente stimola la
gluconeogenesi attivando la piruvato
carbossilasi.
In questo modo l'eccesso di piruvato può essere
49
convertito in glucosio
Fig. 20.6
50
Il
secondo
punto
di
controllo
della
gluconeogenesi è a livello della reazione
catalizzata dalla fruttosio 1,6-bisfosfatasi, che è
inibita dall'AMP.
Il
corrispondente
enzima
glicolitico,
la
fosfofruttochinasi-1,
è
invece
stimolato
dall'AMP e dall'ADP, mentre è inibito dall'ATP e
dal citrato.
Le due reazioni delle due vie sono quindi
regolate in modo coordinato e complementare.
51
Quando nella cellula sono presenti
concentrazioni sufficienti di acetil-CoA
o del suo prodotto di condensazione con
l'ossalacetato (il citrato),
oppure quando i nucleotidi adenilici sono per
la maggior parte nella forma di ATP,
viene favorita la gluconeogenesi.
52
La
funzione
speciale
del
fegato
nel
mantenere costante il livello di glucosio nel
sangue richiede un ulteriore meccanismo di
regolazione che coordini la produzione al
consumo.
53
Quando il livello di glucosio nel sangue
diminuisce, l'ormone glucagone segnala al
fegato di produrre e rilasciare più glucosio.
Una delle fonti di questo glucosio è il
glicogeno conservato nel fegato
L'altra è la gluconeogenesi.
54
Il F2,6-bifosfato attiva
la fosfofruttochinasi I e
inibisce la fruttosio 1,6 bifosfatasi
Formazione e degradazione del fruttosio 2,6 bisfosfato
55
Struttura dei domini dell’enzima bifunzionale fosfofruttochinasi 2
PFK2 e FBPasi sono presenti in una singola catena polipeptidica
2
3
1
Sequenza amminoacidica dell’enzima
56
Fegato e cuore contengono differenti isoenzimi dell’enzima
bifunzionale PFK/FBPasi che danno differente risposta allo stesso
ormone (adrenalina).
adrenalina
glucagone
Cuore:
(attivazione glicolisi)
Fegato:
(inibizione della glicolisi)
57
Il glucagone e l’insulina hanno effetti a lungo termine sui livelli
degli enzimi epatici coinvolti nella glicolisi e nella gluconeogenesi.
1. Nel fegato un elevato rapporto ematico glucagone/insulina
aumenta l’attività degli enzimi glucogenetici mentre dimunuisce
quella degli enzimi glicolitici.
2. Un basso rapporto glucagone/insulina ha gli effetti opposti.
Nel primo caso il glucagone segnala l’induzione della sintesi di
quantità maggiori di:
PEP carbossichinasi
fruttosio-1,6-difosfato
glucosio-6-fosfatasi
varie transaminasi
58
Il legame del glucagone col propro recettore determina un
aumento dei livelli di cAMP
1. Attivazione della proteina kinasi A (PKA) che fosforila una proteina,
CREB (cAMP response element binding protein)
2. Nella sua forma fosforilata CREB, può legarsi all’elemento di
risposta al cAMP : CRE (cAMP response element)
3. CRE a sua volta è un elemento che agisce nella regione di regolazione
dei geni che rispondono al cAMP.
4. Tutto questo promuove la trascrizione dei geni che codificano enzimi
chiave della gluconeogenesi quali la PEP carbossichinasi.
5. Con un meccanismo simile che però causa repressione della
trascrizione genica il glucagone determina la diminuzione dei livelli
di glucochinasi,fosofrutto-1-chinasi e piruvato chinasi.
59
Il promotore del gene per la PEPcarbossichinasi
Gli ormoni influenzano l’espressione genica variando la velocità di trascrizione.
IRE = elemento di risposta all’insulina
GRE = elemento di risposta ai glucocorticoidi
TRE = elemento di risposta all’ormone tiroideo
CRE I e CRE II = elementi di risposta all’cAMP
60
L’Insulina si oppone all’azione del glucagone con un meccanismo
a cascata
1. Attivazione di una proteina legante l’elemento di risposta all’nsulina
IREB.
2. gluconeogenesi legandosi all’elemento di risposta all’insulina IRE nella
regione IREB inibisce la trascrizione dei geni che codificano gli enzimi
chiave della di regolazione di tali geni.
3. Quando la sintesi di glucosio non è necessaria, in conseguenza
dell’aumento del rapporto insulina/glucagone nel sangue viene interrotta la
biosintesi degli enzimi chiave della gluconeogenesi mentre viene attivata
quella degli enzimi chiave della glicolisi
61
62
ciclo di Cori e ciclo dell’alanina
Nell’ambito
della gluconeogenesi sono stati
descritti 2 importanti cicli cui partecipano differenti
tessuti.
Il ciclo di Cori e il ciclo dell’alanina prevedono
una prima fase di gluconeogenesi che si svolge nel
fegato, cui seguono la distribuzione e l’utilizzazione
del glucosio nei tessuti periferici.
Lo
scopo di entrambi i cicli è quello di rifornire
continuamente di glucosio i tessuti che dipendono da
esso come fonte energetica primaria.
63
I cicli sono attivi soltanto tra il fegato e i tessuti
che non ossidano completamente il glucosio a CO2 e
H2O.
Per partecipare a questi processi, il tessuto
periferico deve liberare alanina o acido lattico come
prodotto finale del metabolismo del glucosio.
64
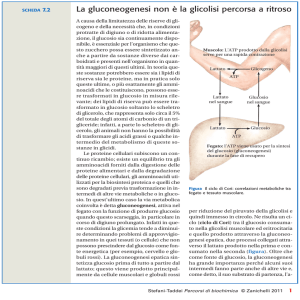
Ciclo di Cori
65
(a) Ciclo di Cori
(a)
Correlazione fra
gluconeogenesi
epatica e glicolisi
nel resto
dell’organismo.
(b)
b) Ciclo dell’alanina
66
La differenza sostanziale tra:
il ciclo di Cori
e il ciclo dell’alanina
• consiste nel tipo di intermedio a tre atomi di
carbonio che viene riciclato; infatti nel ciclo di Cori il
carbonio torna al fegato come lattato e nel ciclo
dell’alanina come alanina.
• un’altra differenza è che nel ciclo dell’alanina il
NADH ottenuto dalla glicolisi non può essere utilizzato
per produrre piruvato dal lattato
67
Nei tessuti forniti di mitocondri, gli elettroni del NADH
vengono trasferiti in questi organelli attraverso il sistema
navetta malato-aspartato o quello del glicerofosfato per
la sintesi di ATP attraverso la fosforilazione ossidativa
NADH + H+ + 1/2O2 + 3ADP + 3Pi
NAD+ + 3 ATP + H2O
oppure
FADH2 + 1/2O2 + 2ADP + 2Pi
FAD + 2ATP + H2O
Di conseguenza per ogni molecola di glucosio che partecipa
nel ciclo dell’alanina si possono produrre più molecole di ATP,
mentre nel ciclo di Cori vengono prodotti soltanto 2 ATP per
ogni molecola di glucosio.
68
Nel fegato le molecole di ATP
necessarie per la gluconeogenesi.
sono
Il ciclo dell’alanina trasferisce anche energia
nei tessuti periferici; poiché produce da 6-8
molecole di ATP per molecola di glucosio, il
ciclo dell’alanina è più efficiente dal punto di
vista energetico.
69
La partecipazione dell’alanina al ciclo porta al fegato
azoto amminico, che deve essere eliminato come
urea.
La sintesi dell’urea è energeticamente dispendiosa,
richiedendo il consumo di 4 molecole di ATP per ogni
molecola di urea prodotta.
Le necessità energetiche per la sintesi dell’urea
determinano un aumento della quantità di ATP
necessario per sintetizzare una singola molecola di
glucosio nel fegato.
70
Cooperazione
metabolica
tra
muscolo scheletrico e fegato.
Durante un'attività muscolare molto
intensa,
il
muscolo
scheletrico
utilizza glicogeno come fonte di
energia,
generando
lattato
attraverso la glicolisi.
Durante il periodo di recupero, una
parte del lattato formato nel muscolo
viene trasportato al fegato e usato
per produrre glucosio mediante la
gluconeogenesi.
Il glucosio prodotto viene rilasciato
nel sangue e ritorna al muscolo per
ripristinare le riserve di glicogeno.
Questa via (glucosio lattato 71
glucosio) costituisce il ciclo di Cori.
1
L'uso dell'alanina come trasportatore di
ammoniaca dal muscolo scheletrico che
sta lavorando al fegato è un altro esempio
dell'economia esistente negli organismi
viventi.
Il muscolo che si contrae violentemente
opera
in
condizioni
anaerobiche,
producendo non solo ammoniaca dalla
degradazione delle proteine, ma anche
una grande quantità di piruvato e lattato
dalla glicolisi.
72
2
Questi prodotti devono essere trasportati
al fegato: l'ammoniaca per essere
convertita in urea ed essere escreta, il
piruvato e il lattato per essere riconvertiti
in glucosio e ritornare al muscolo.
Il ciclo del glucosio-alanina, insieme con il
ciclo di Cori realizza questa operazione. Il
dispendio energetico della gluconeogenesi
è quindi a carico del fegato e tutto l'ATP
presente nel muscolo può essere utilizzato
per la contrazione muscolare.
73
L’aspartato fuoriesce dal mitocondrio e partecipa
alciclo dell’urea reagendo con la citrullina. Il
carbonio dell’aspartato abbandona il ciclo dell’urea
sotto forma di fumarato, successivamente
trasformato in malato ad opra della fumarasi
citosolica.
Sia questo malato che quello mitocondriale sono
trasformati in glucosio per azione degli enzimi
citosolici della gluconeogenesi.
74
I triacilgliceroli sono formati da tre gruppi O-acilici
combinati con una molecola di glicerolo.
L’idrolisi di un triacilglicerolo fornisce tre acidi
grassi e glicerolo.
Il glicerolo è un eccellente substrato per la
gluconeogenesi.
La fosforilazione del glicerolo ad opera della
glicerolo –chinasi produce glicerolo-3-fosfato che
può essere trasformato in diidrossiacetonfosfato,
un intermedio della gluconeogenesi per mezzo della
glicerolofosfato deidrogenasi.
75
76
Ossidazione degli acidi grassi a catena dispari:
Il propionato
si forma propionil CoA che
viene convertito in
succinil CoA
Il succinil CoA non viene direttamente consumato
dal ciclo dell’acido citrico ma viene prima
convertito in piruvato e poi in acetil CoA
77
Il propionil CoA che proviene dall’ossidazione degli cidi grassi a numero DISPARI di atomi di
carbonio nonché da alcuni amminoacidi, viene convertito a succinil CoA un intermedio del ciclo di
Krebs
La metil malonil CoA mutasi utilizza
un gruppo prostetico 5’ deossiadenosilcobalammina,
un derivato della vitamina B12
78
L’assunzione di etanolo inibisce la gluconeogenesi.
O
CH2-CH2-OH + NAD+
etanolo
CH3-CH + NADH + H+
acetaldeide
Piruvato + N ADH + H+
Lattato + NAD+
Oppure
Ossalacetato + NADH+H+
malato+ NAD+
L’etanolo viena ossidato principalmente nel fegato con produzione di una grande quantità di equivalenti
riducenti che devono essere trasportati nel mitocondrio attraverso il sistema navetta malato-aspartato.
L’eccesso di NADH nel citosol crea dei problemi per la gluconeogenesi epatica poiché spinge
l’equlibrio delle reazioni catalizzate dalla lattato deidrogenasi e della malato deidrogenasi
rispettivamente nella direzione del lattato e del malato.
In condizioni in cui tali reazioni vengono forzate a decorrere nella direzione in cui sono scritte si ha
inbizione della sintesi di glucosio in quanto risultano limitanti le quantità di piruvato e ossalacetato
79
disponibili per le reazioni catalizzate, rispettivamente dalla piruvato carbossilasi e dalla PEPcarbossichinasi.
Ipoglicemia e intossicazione da alcol
Acidosi lattica
Ipoglicemia nei neonati prematuri:
I bambini sono più sensibili degli adulti all’ipoglicemia perché il
rapporto in peso fra cervello e corpo è maggiore nei bambini e il
loro cervello impiega quantità di glucosio sproporzionatamente
maggiori rispetto al resto del corpo.
1) Limitata capacità chetogenetica
2) Scarsa quantità di PEPCK: ridotta capacità di sintetizzare
glucosio da lattato e alanina.
3) Ridotti depositi di glicogeno epatico
Neonati più dipendenti dalla gluconeogenesi di quelli normali
80
FINE
81