Gluconeogenesi e sintesi dei
carboidrati
La gluconeogenesi è il processo che produce glucosio da
molecole più semplici. Nelle piante è una delle principali
vie che porta alla possibile sintesi di carboidrati più
complessi
Gli animali possono essere invece divisi in due gruppi a
seconda dell’importanza della gluconeogenesi:
1) I non ruminanti che prendono il glucosio presente nel
cibo attraverso la rottura dei carboidrati a livello del
carbonio e in cui, a parte un modesto apporto di glicogeno
sintetizzato, il metabolismo del glucosio si riduce
essenzialmente all’ossidazione di tale composto
2) I ruminanti che ricevono un limitato apporto di
glucosio dall’intestino contando maggiormente
sulla sintesi e utilizzo di acidi grassi,
principalmente propionato.
Il percorso è veramente attivo perché i ruminanti
hanno una grossa richiesta per il glucosio come
fonte energetica nei tessuti e, nel caso di animali in
lattazione, per produrre lattosio
I materiali di partenza
Molti metaboliti possono essere usati come materiali di
partenza per la gluconeogenesi
Composti a tre atomi di carbonio come il PIRUVATO, il
FOSFOENOLPIRUVATO o i TRIOSO FOSFATI possono
essere tutti trasformati in glucosio e sono detti materiali
gluconeogenetici
La reazione dove l’acetil-CoA è prodotto dal piruvato
(piruvato deidrogenasi) è irreversibile in organismi dove
manca il ciclo del gliossilato
Quindi l’acetil-CoA e i composti collegati (gli acidi grassi,
corpi chetonici, gli amminoacidi chetogeni) non possono
essere usati per la sintesi del glucosio
Il ciclo gluconeogenetico
In realtà il percorso metabolico è molto simile a una
glicolisi inversa. Esistono tuttavia differenze tali che
permettono alla via di procedere verso la sintesi di
glucosio all’interno della cellula
Come noto la glicolisi presenta delle reazioni irreversibili
che debbono venire in qualche modo “superate” se
vogliamo procedere in senso inverso. Ciò viene effettuato
attraverso l’utilizzazione di enzimi diversi che catalizzano
reazioni che altrimenti sarebbero irreversibili
Differenze tra gluconeogenesi e
glicolisi
Ci sono tre reazioni nella glicolisi che sono
essenzialmente irreversibili perché sono accompagnate
da un cambiamento di energia libera estremamente
negativa:
1) esochinasi/glucochinasi
2) fosfo fruttochinasi
3) piruvato chinasi
Il percorso che porta alla sintesi del glucosio deve
superare queste reazioni irreversibili attraverso una serie
di reazioni by-pass. Queste reazioni contribuiscono
anche al controllo del flusso del percorso biochimico
Gluconeogenesi attraverso il
piruvato
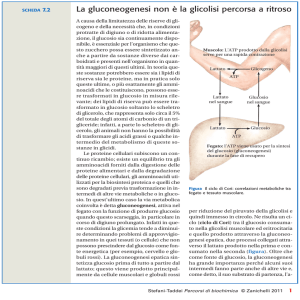
In condizioni normali il piruvato non si trova in alte
concentrazioni nelle cellule, ma altri precursori
come malato, lattato e amminoacidi glucogenici
hanno come intermedio questo metabolita
A causa dell’irreversibilità dell’enzima piruvato
chinasi, la produzione diretta di fosfoenolpiruvato
dal piruvato non è possibile; per questo viene
seguito un percorso a due stadi
Tale breve sequenza è complicata dal fatto che
essa ha luogo in parti differenti della cellula
L’aggiramento della piruvato chinasi richiede quindi due
reazioni, la prima delle quali è catalizzata dalla PIRUVATO
CARBOSSILASI. La reazione, ATP- e Biotina-dipendente,
converte il piruvato in ossalacetato
Questa reazione rappresenta una via anaplerotica attraverso
cui viene mantenuto costante il livello di OAA nel ciclo TCA
ed avviene nella matrice mitocondriale
Questo OAA, formato per poter essere utilizzato nella
gluconeogenesi, deve spostarsi dal mitocondrio al citosol
dove invece avvengono le reazioni della gluconeogenesi
La membrana mitocondriale è tuttavia relativamente
impermeabile all’OAA di conseguenza l’OAA viene ridotto a
MALATO dalla MALATO DEIDROGENASI mitocondriale
Il malato viene quindi trasportato nel citosol e qui, ad opera di
una MALATO DEIDROGENASI citosolica, viene nuovamente
trasformato in OAA
Una volta nel citosol l’OAA viene convertito in PEP ad opera
della FOSFOENOLPIRUVATO CARBOSSICHINASI (PEPCK). La
CO2 fissata dalla piruvato carbossilasi viene ora persa per cui
non si ha fissazione netta di questo gas
Questo passaggio richiede energia per avvenire, in questo caso
sotto forma di GTP
Questo significa che la conversione del piruvato in
fosfoenolpiruvato necessita di due gruppi fosfato ad alta
energia, mentre la reazione inversa ne produce uno solo
Se entrambe le sequenze di queste reazioni fossero attive allo
stesso tempo, l’effetto totale sarebbe che una mole di ATP e una
di GTP sarebbero usate e solo una mole di ATP sarebbe prodotta
MITOCONDRIO
GLUCONEOGENESI
La produzione di fruttosio-1,6-difosfato
La conversione del fosfoenolpiruvato in fruttosio1,6-difosfato è l’esatto inverso dei passaggi
equivalenti nella glicolisi. I cambiamenti di energia
libera per ciascuna di queste reazioni sono molto
piccoli
Conversione del fruttosio-1,6-bisfosfato in fruttosio-6-fosfato
Nella glicolisi, la fosfofruttochinasi usa l’ATP per aggiungere il
gruppo fosfato al fruttosio-6-fosfato in una reazione
praticamente irreversibile
Nella gluconeogenesi l’azione inversa coinvolge la semplice
idrolisi del gruppo fosfato attraverso l’enzima FRUTTOSIO1,6-DIFOSFATASI
Fruttosio-1,6-bisfosfato + H2O fruttosio-6-fosfato + Pi
G0’ = - 16.3 kJ/mol
L’enzima richiede ioni Mg e rappresenta uno dei più
importanti siti di regolazione della via
Conversione del fruttosio-6fosfato in glucosio
La conversione da fruttosio-6-P a glucosio-6-P è
operata dalla FOSFOESOSO ISOMERASI, mentre
la reazione sino a glucosio necessita di un altro
enzima, la GLUCOSIO-6-FOSFATASI, che opera
una semplice idrolisi del gruppo fosfato
Stechiometria e bilancio energetico
della gluconeogenesi
La conversione di 2 moli di piruvato in una mole di glucosio è
una processo esoergonico nel suo complesso
Il G0’ per il processo complessivo è pari a - 47.6 kJ/mol
Occorre infatti considerare che vengono consumate 6 moli di
ATP e 2 moli di NADH (altri 6 ATP in termini di riossidazione)
Se, al contrario, la glicolisi procedesse in senso inverso, si
avrebbe il consumo di 2 moli di NADH e 2 di ATP, ma la
reazione sarebbe tuttavia altamente endoergonica (G0’ =+73.3
kJ/mol)
Substrati per la gluconeogenesi
La gluconeogenesi può avere diversi precursori, vediamo
i più importanti:
Lattato
Precursore più significativo della gluconeogenesi, basti
pensare alla sua produzione tramite la fermentazione
omolattica
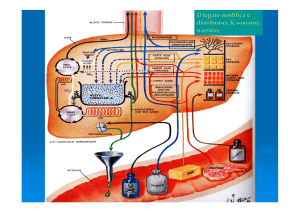
Negli animali il lattato che entra nel fegato viene riossidato
a piruvato il quale, tramite la gluconeogenesi, si trasforma
in glucosio, ceduto al sangue ed assorbito dal muscolo
dove può essere convertito in glicogeno
Amminoacidi
Gli amminoacidi possono venire convertiti in
glucosio soprattutto attraverso le vie degradative
che producono gli intermedi del ciclo TCA, i quali
a loro volta possono venire convertiti in OAA
Questi amminoacidi sono definiti GLUCOGENICI
Tra tutti gli a.a. essenziali solo la LISINA e la
LEUCINA non generano precursori gluconeogenici
Propionato
Un percorso che è di grande importanza
nell’agricoltura è quello che avviene in molti
ruminanti
Nei non ruminanti la maggior forma di energia
utilizzabile è quella sotto forma di glucosio;
nei ruminanti tale ruolo è assunto da un gruppo di
acidi
grassi
volatili
(VFAs).
Nonostante
l’importanza che i VFAs rivestono nella nutrizione
dei ruminanti, gli animali hanno ancora bisogno
del glucosio per una serie di importanti funzioni
quali il rifornimento di energia al cervello e ai
tessuti nervosi
L’importanza del glucosio cresce gradualmente
durante la lattazione. Solo uno dei più importanti
VFAs, il propionato, è capace di servire come
precursore per la sintesi del glucosio
Il primo passo del percorso metabolico è simile a
quello degli acidi grassi: la conversione del
propionato in propionil-CoA. Quindi, gli unici
passaggi inusuali sono quelli che convertono il
propionato in succinil-CoA
Dapprima una propionil-CoA carbossilasi
catalizza la formazione del metilmalonil-CoA
Il meccanismo di reazione è molto simile ad
uno dei primi stadi della sintesi degli acidi
grassi quando la CO2 viene aggiunta all’acetilCoA per produrre malonil-CoA (acetil-CoA
carbossilasi)
La reazione, che è irreversibile, richiede
energia sotto forma di ATP ed utilizza la biotina
come coenzima, trasportatore di CO2
Il metilmalonil-CoA formatosi viene modificato
per
produrre
succinil-CoA
dall’enzima
metilmalonil-CoA mutasi
Questo enzima è abbastanza inusuale perché è
uno dei pochi che usa la vitamina B12 come
cofattore.
La deficienza di cobalto nei ruminanti potrebbe
dunque, interferire con questa reazione che è
fondamentale in tali organismi
Il succinil-CoA è usato per produrre
fosfoenolpiruvato attraverso l’ossalacetato,
che viene poi usato per la sintesi del glucosio6-fosfato
Regolazione della gluconeogenesi
La gluconeogenesi e la glicolisi avvengono entrambe nel
citosol e poiché la gluconeogenesi produce carboidrati
mentre la glicolisi li catabolizza, le due vie devono essere
regolate in modo reciproco
In altre parole le condizioni cellulari che inibiscono una via
attivano l’altra e viceversa
I principali siti di regolazione della gluconeogenesi sono al
livello delle reazioni catalizzate dalla GLUCOSIO-6FOSFATASI,
FRUTTOSIO-1,6-BISFOSFATASI
e
dalla
combinazione della PIRUVATO CARBOSSILASI e della PEP
CARBOSSICHINASI
Glucosio-6-fosfatasi
Non è regolata allostericamente, ma la sua Km per il
glucosio-6-fosfato è molto più alta della sua
concentrazione intracellulare;
in questo modo l’attività è regolata dalla concentrazione
di questo substrato in una cinetica di 1 ordine
Fruttosio-1,6-bisfosfatasi
E’ fortemente inibita dall’AMP così come la fosfofruttochinasi è attivata da AMP
Questo ad indicare che quando la carica energetica
decresce la glicolisi è attivata e la gluconeogenesi è
inibita
Vi sono tuttavia altri meccanismi di controllo