METABOLISMO AEROBIO
I NADH prodotti durante la glicolisi nel citosol, in condizioni aerobiche (respirazione
mitocondriale attiva), possono trasferire elettroni all’interno del mitocondrio
attraverso i Sistemi Navetta,
in questo modo vengono recuperati NAD+ citosolici necessari per la glicolisi.
SISTEMA NAVETTA diidrossiacetone-fosfato/glicerolo 3-fosfato
NEL CITOSOL: IL NADH VIENE OSSIDATO A NAD+
E IL DIIDROSSIACETONEFOSFATO RIDOTTO A GLICEROLO 3-FOSFATO
È SFRUTTATA L’ATTIVITA’ DELLA GLICEROLO 3-FOSFATO DEIDROGENASI
AGISCONO NEL SISTEMA NAVETTA 2 ISOFORME DELL’ENZIMA:
• UNA FORMA CITOSOLICA NADH-dipendente
• UNA FORMA MITOCONDRIALE ( MEMBRANA MIT. INTERNA) FAD-dipendente
Glicerolo 3-fosfato deidrogenasi
mitocondriale (FAD-dipendente)
Direttamente al
complesso III
Membrana
mitocondr. interna
METABOLISMO ANAEROBIO
Il NADH viene ri-ossidato attraverso la Fermentazione lattica o alcolica, in questo modo
la glicolisi può continuare a procedere e a produrre ATP anche se la fosforilazione
ossidativa mitocondriale è assente, scarsa o rallentata. La fonte di energia della cellula
è l’ATP glicolitico.
FERMENTAZIONE LATTICA:
Nei mammiferi la produzione di lattato è essenziale negli eritrociti, nei tessuti
glicolitici obbligati – parti del cervello, midollare del rene- e nella cornea. In questi
tipi cellulari non avvengono biosintesi, non ci sono mitocondri (eritrociti) non
avviene o è scarsa la fosforilazione ossidativa perché c’è uno scarso apporto di
ossigeno.
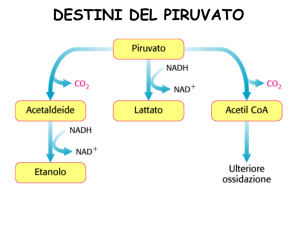
Lattato deidrogenasi
Piruvato
L-lattato
Disponibile per la gliceraldeide 3-P deidrogenasi
LA FERMENTAZIONE LATTICA è UN PROCESSO CHE AVVIENE
INTENSAMENTE NELLE CELLULE MUSCOLARI SOTTO INTENSO SFORZO.
Ciclo di Cori: è un ciclo di metaboliti che
si verifica tra il muscolo e il fegato
MUSCOLO
Glicogeno
FEGATO
SANGUE
ADP+GDP +PI
ATP+ GTP
Glucosio
ATP
glicolisi
gluconeogensi
NADH
Piruvato
Piruvato
NAD+
Glucosio
LATTATO
SANGUE
LATTATO
FERMENTAZIONE ALCOLICA: Molti batteri e alcuni eucarioti (lieviti)
possono sopravvivere in assenza di O2 metabolizzando il piruvato
ottenuto dalla glicolisi
TPP, Mg2+
OSSIDAZIONE
DECARBOSSILAZIONE TPP-DIPENDENTE
Effetto netto: mantenere il
flusso glicolitico e la
produzione di ATP
Reso disponibile per la
Gliceraldeide 3-fosfato
deidrogenasi
Gluconeogenesi
Il glucosio può essere sintetizzato a partire da precursori più semplici,
i precursori possono essere vari a seconda della specie vivente, nei
mammiferi sono: piruvato, lattato, ossalacetato, amminoacidi,
glicerolo.
Avviene nel CITOSOL delle cellule del fegato e dei reni
È operativa in condizioni di digiuno estremo (dopo le 24 h) → forte ipoglicemia e
dopo deplezione delle riserve di glicogeno endogeno.
La gluconeogenesi condivide con la glicolisi tutte le reazioni reversibili
le quali procedono in direzione opposta quando utilizzate nella sintesi
Le reazioni irreversibili della glicolisi sono
aggirate attraverso l’azione di enzimi differenti
LA GLUCONEOGENESI A PARTIRE DAL PIRUVATO
2 piruvato + 2 NADH + 4 ATP + 2 GTP + 6 H2O + 2 H+ →
Glucosio + 2 NAD+ + 4 ADP + 2 GDP + 6Pi
La reazione della glicolisi con cui si ottiene PIRUVATO dal FOSOFOENOLPIRUVATO è
IRREVERSIBILE: è catalizzata dalla PIRUVATO CHINASI che NON PUO’ catalizzare la
REAZIONE INVERSA
Per riottenere FOSFOENEOLPIRUVATO è necessario utilizzare una via
alternativa: reazioni catalizzate da enzimi differenti.
Il piruvato prodotto dalla glicolisi è immediatamente trasportato dentro il
mitocondrio dove può subire diversi destini metabolici.
SE È NECESSARIO UTILIZZARLO PER LA SINTESI DI GLUCOSIO, IL PIRUVATO
VIENE CARBOSSILATO A OSSALACETATO.
Bicarbonato
+
piruvato
CARBOSSILAZIONE DEL PIRUVATO
POICHÉ LA GLUCONEOGENESI
AVVIENE NEL CITOSOL,
L’OSSALACETATO DOVRA’ USCIRE
DAL MITOCONDRIO.
Piruvato
carbossilasi
Ossalacetato
Legame
fosfoanidridico
ad alta energia
La biotina
attacca la CO2
Nel sito
attivo
dell’enzima
si produce
CO2
IN CONDIZIONI DI FORTE IPOGLICEMIA: TUTTO L’ossalacetato
presente nel mitocondrio, viene sottratto ad altre vie
metaboliche e indirizzato verso la gluconeogenesi.
Ossalacetato
Malato
deidrogenasi
mitocondriale
NADH + H+
NAD+
H
Malato
l
OH
Malato
deidrogenasi
citosolica
MITOCONDRIO
CITOSOL
NAD+
NADH +
H+
Si genera una scorta di
NADH citosolici che sono
necessari per il
proseguimento della
gluconeogenesi
Ossalacetato
L’ossalacetato nel citosol è convertito in fosfoenolpiruvato dalla
FOSFOENOLPIRUVATO CARBOSSICHINASI
Ossalacetato: viene
decarbossilato con
conseguente formazione
di un enolo che poi viene
fosforilato
2-fosfoglicerato
Enolasi
Fosfoenolpiruvato
FOSFOENOLPIRUVATO
Fosfoenolpiruvato carbossichinasi
mitocondriale
Piruvato carbossilasi
mitocondrio
citosol
Produce il
citosolico necessario
alla gluconeogenesi
Lattato deidrogenasi
GLUCONEOGENESI
Reazioni inverse della
glicolisi
(stessi enzimi e stessi
intermedi metabolici)
BYPASS ALLA
PIRUVATO CHINASI
GLICOLISI
FosfoFruttochinasi 1
Glicolisi
Gluconeogenesi
Glucosio 6-fosfato
H
β-D-Fruttosio 1,6-bisfosfato
(F1,6P)
Fruttosio 1,6bisfosfatasi 1
GLUCONEOGENESI
Reazione alternativa
GLICOLISI
Il glucosio neosintetizzato è liberato dagli epatociti nel torrente
circolatorio per raggiungere i tessuti extraepatici che sono in carenza di
glucosio (in particolare il cervello)
GLUCONEOGENESI
Reazione alternativa
Glucosio 6-fosfato
Reazione alternativa
GLICOLISI
Vari amminoacidi sono precursori del glucosio durante il digiuno, per es.
l’alanina, che partecipa al Ciclo Glucosio/Alanina con cui il piruvato prodotto
nei tessuti extraepatici raggiunge il fegato per essere indirizzato verso la
gluconeogenesi
Piruvato
Glutammato
ALANINA
Transamminazione nel muscolo
α-chetoglutarato
Alanina trasportata al fegato
attraverso il circolo sanguigno
ALANINA
α-chetoglutarato
Piruvato
Glutammato
Transamminazione epatica, il piruvato
può essere utilizzato per sintetizzare
glucosio